
Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens


Abstract
:1. Introduction
2. Classification and Sources of Lactic Acid Bacteria
3. Bacteriocins Produced by Lactic Acid Bacteria
3.1. Classificatio of Lactic Acid Bacteria Bacteriocins
3.2. Production and Biosynthesis of LAB Bacteriocins
3.3. Growth Conditions for Optimum Production of LAB Bbacteriocins
4. Impact of Foodborne Pathogens on Human Health
5. Activity of LAB and Their Bacteriocins against Foodborne Pathogens
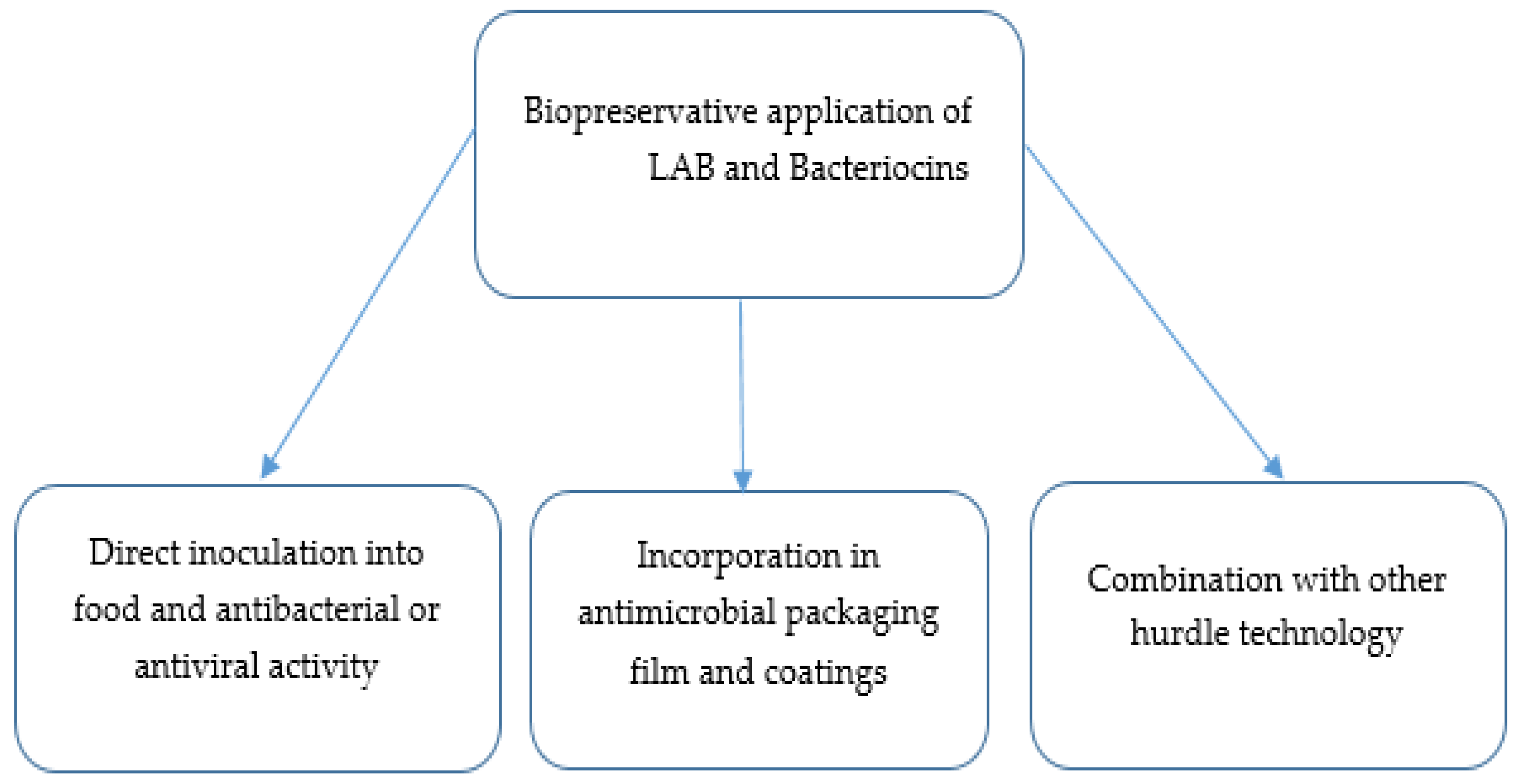
5.1. Direct LAB Incorporation and Activity against Foodborne Bacteria
5.2. Direct Lactic Acid Bacteria Incorporation and Activity against Foodborne Viruses
5.3. Bacteriocins in Antimicrobial Packaging Films and Coatings
5.4. Lactic Acid Bacterial Bacteriocins Combination with Other Hurdle Technologies
6. Future Prospects of LAB and Bacteriocins in the Food Industry
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Geel-Schuttená, G.H.; Flesch, F.; ten Brink, B.; Smith, M.R.; Dijkhuizen, L. Screening and characterization of Lactobacillus strains producing large amounts of exopolysaccharides. Appl. Microbiol. Biotech. 1998, 50, 697–703. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Leroy, F. Bacteriocins from lactic acid bacteria: Production, purification and food applications. J. Mol. Microbiol. Biotech. 2007, 13, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Kaban, G.; Kaya, M. Identification of lactic acid bacteria and Gram-positive catalase-positive cocci isolated from naturally fermented sausage (sucuk). J. Food Sci. 2008, 73, M385–M388. [Google Scholar] [CrossRef] [PubMed]
- Hati, S.; Mandal, S.; Prajapati, J.B. Novel starters for value added fermented dairy products. Curr. Res. Nutr. Food Sci. 2013, 1, 83–91. [Google Scholar] [CrossRef]
- Alakomi, H.-L.; Skyttä, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic acid permeabilizes Gram-negative bacteria by disrupting the outer membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga, M.; Pardo, I.; Ferrer, S. An improved medium for distinguishing between homofermentative and heterofermentative lactic acid bacteria. Int. J. Food Microbiol. 1993, 18, 31–42. [Google Scholar] [CrossRef]
- McDonald, L.C.; McFeeters, R.F.; Daeschel, M.A.; Fleming, H.P. A differential medium for the enumeration of homofermentative and heterofermentative lactic acid bacteria. Appl. Env. Microbiol. 1987, 53, 1382. [Google Scholar] [CrossRef] [Green Version]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, P.K.; Nehra, K. Bacteriocin-nanoconjugates as emerging compounds for enhancing antimicrobial activity of bacteriocins. J. King Saud Univ. Sci. 2019, 31, 758–767. [Google Scholar] [CrossRef]
- Field, D.; Ross, R.P.; Hill, C. Developing bacteriocins of lactic acid bacteria into next generation biopreservatives. Curr. Opin. Food Sci. 2018, 20, 1–6. [Google Scholar] [CrossRef]
- Kaya, H.I.; Ozel, B.; ̧Simsek, O. A natural way of food preservation: Bacteriocins and their applications. In Health and Safety Aspects of Food Processing Technologies; Malik, A., Erginkaya, Z., Erten, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 633–659. [Google Scholar]
- Bourdichon, F.; Laulund, S.; Tenning, P. Inventory of microbial species with a rationale: A comparison of the IDF/EFFCA inventory of microbial food cultures with the EFSA Biohazard Panel qualified presumption of safety. FEMS Microbiol. Lett. 2019, 366, fnz048. [Google Scholar] [CrossRef] [PubMed]
- Panel, E.B.; Herman, L. Statement on the update of the list of QPS-recommended bio-logical agents intentionally added to food or feed as notified to EFSA 14: Suitability of taxonomic units notified to EFSA until March 2021. EFSA J. 2021, 19, e06377. [Google Scholar]
- Parada, J.L.; Caron, C.R.; Medeiros, A.B.P.; Soccol, C.R. Bacteriocins from lactic acid bacteria: Purification, properties and use as biopreservatives. Braz. Arch. Biolol. Tech. 2007, 50, 521–542. [Google Scholar] [CrossRef] [Green Version]
- Naidu, A.S.; Bidlack, W.R.; Clemens, R.A. Probiotic spectra of lactic acid bacteria (LAB). Crit. Rev. Food Sci. Nutr. 1999, 38, 13–126. [Google Scholar] [CrossRef]
- Todorov, S.D. Bacteriocins from Lactobacillus plantarum—production genetic organization. Braz. J. Microbiol. 2009, 40, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Devi, M.; Rebecca, L.J.; Sumathy, S. Bactericidal activity of the lactic acid bacteria Lactobacillus delbreukii. J. Chem. Paharmaceutical Res. 2013, 5, 176–180. [Google Scholar]
- Liu, W.; Pang, H.; Zhang, H.; Cai, Y. Biodiversity of lactic acid bacteria. In Lactic Acid Bacteria; Zhang, Y., Cai, Y., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 103–203. [Google Scholar] [CrossRef]
- Djadouni, F.; Kihal, M. Antimicrobial activity of lactic acid bacteria and the spectrum of their biopeptides against spoiling germs in foods. Braz. Arch. Biol Tech. 2012, 55, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Walter, J. Ecological role of lactobacilli in the gastrointestinal tract: Implications for fundamental and biomedical research. Appl. Environ. Microbiol. 2008, 74, 4985–4996. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, E.J.C.; Tyrrell, K.L.; Citron, D.M. Lactobacillus species: Taxonomic complexity and controversial susceptibilities. Clin. Infect. Dis. 2015, 60, S98–S107. [Google Scholar] [CrossRef] [Green Version]
- Rajilic-Stojanovic, M.; de Vos, W. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef]
- Alkema, W.; Boekhorst, J.; Wels, M.; van Hijum, S.A.F.T. Microbial bioinformatics for food safety and production. Brief. Bioinform. 2016, 17, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belicova, A.; Mikulasova, M.; Dusinsky, R. Probiotic potential and safety properties of Lactobacillus plantarum from Slovak bryndza cheese. Biomed. Res. Intl. 2013, 2013, 760298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbel, S.R.; Vahjen, W.; Wieler, L.H.; Guenther, S. Timely approaches to identify probiotic species of the genus Lactobacillus. Gut Pathog. 2013, 5, 1–13. Available online: http://www.gutpathogens.com/content/5/1/27 (accessed on 14 March 2017).
- Zacharof, M.P.; Lovitt, R.W. Bacteriocins produced by lactic acid bacteria: A review article. APCBEE Procedia 2012, 2, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Aljewicz, M.; Cichosz, G. Influence of probiotic (Lactobacillus acidophilus NCFM, L. paracasei LPC37, and L. rhamnosus HN001) strains on starter cultures and secondary microflora in Swiss-and Dutch-type cheeses. J. Food Process. Preserv. 2017, 41, e13253. [Google Scholar] [CrossRef]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Factories 2014, 13, 1–13. Available online: http://www.microbialcellfactories.com/content/13/S1/S3 (accessed on 2 March 2017). [CrossRef] [Green Version]
- Hu, C.B.; Malaphan, W.; Zendo, T.; Nakayama, J.; Sonomoto, K. Enterocin X, a novel two-peptide bacteriocin from Enterococcus faecium KU-B5, has an antibacterial spectrum entirely different from those of its component peptides. Appl. Environ. Microbiol. 2010, 76, 4542–4545. [Google Scholar] [CrossRef] [Green Version]
- Oppegard, C.; Rogne, P.; Kristiansen, P.E.; Nissen-Meyer, J. Structure analysis of the two-peptide bacteriocin lactococcin G by introducing D-amino acid residues. Microbiology 2010, 156, 1883–1889. [Google Scholar] [CrossRef] [Green Version]
- Diep, D.B.; Mathiesen, G.; Eijsink, V.G.H.; Nes, I.F. Use of lactobacilli and their pheromone-based regulatory mechanism in gene expression and drug delivery. Current Pharm. Biotech. 2009, 10, 62–73. [Google Scholar] [CrossRef]
- Güllüce, M.; Karadayi, M.; Bariş, Ö. Bacteriocins: Promising antimicrobials. Microbial pathogens and strategies for combating them. In Science, Technology and Education; Mendes-Vilas, A., Ed.; FORMATEX: Madrid, Spain, 2013; pp. 1016–1027. [Google Scholar]
- Dimov, S.; Ivanova, P.; Harizanova, N. Genetics of bacteriocins biosynthesis by lactic acid bacteria. Biotechnol. Biotechnol. Eq. 2005, 19, 4–10. [Google Scholar] [CrossRef]
- Goh, H.F.; Phillip, K. Purification and characterization of bacteriocin produced by Weissella confusa A3 of dairy origin. PLoS ONE 2015, 10, e0140434. [Google Scholar] [CrossRef]
- Elayaraja, S.; Annamalai, N.; Mayavu, P.; Balasubramanian, T. Production, purification and characterization of bacteriocin from Lactobacillus murimus AU06 and its broad antibacterial spectrum. Asian Pac. J. Trop. Biomed. 2014, S305–S311. [Google Scholar] [CrossRef] [Green Version]
- Mahrous, H.; Mohamed, A.; El-Mongy, M.A.; El-Batal, A.I.; Hamza, H.A. Study bacteriocin production and optimization using new isolates of Lactobacillus spp. isolated from some dairy products under different culture conditions. Food Nutr. Sci. 2013, 4, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Leal-Sánchez, M.V.; Jimenéz-Díaz, R.; Maldonado-Barragán, A.; Garrido-Fernández, A.; Ruiz-Barba, J.L. Optimization of bacteriocin production by batch fermentation of Lactobacillus plantarum LPCO10. Appl. Env. Microbiol. 2002, 68, 4465–4471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sure, K.P.; Kotnis, P.V.; Bhagwat, P.K.; Ranveer, R.C.; Dandge, P.B.; Sahoo, A.K. Production and characterization of bacteriocin produced by Lactobacillus viridescence (NICM 2167). Braz. Arch. Biol. Technol. 2016, 59, e16150518. [Google Scholar] [CrossRef] [Green Version]
- Mandal, V.; Sen, S.K.; Mandal, N.C. Optimized culture conditions for bacteriocin production by Pediococcus acidilacti LAB5 and its characterization. Indian J. Biochem Phys. 2008, 45, 106–110. [Google Scholar]
- Malheiros, P.S.; Sant’ Anna, V.; Todorov, S.D.; Franco, B.D.G.M. Optimization of growth and bacteriocin production by Lactobacillus sakei subsp. sakei 2a. Braz. J. Microbiol. 2015, 46, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onwuakor, C.E.; Nwaugo, V.O.; Nnadi, C.J.; Emetole, J.M. Effect of varied culture conditions on crude supernatant (bacteriocin) production from four Lactobacillus species isolated from locally fermented maize (ogi). Am. J. Microbiol. Res. 2014, 2, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Huan, X.; Yang, Y.; Liu, W.; Liu, H.; Zhang, Y.; Sun, K.; Zhang, L.; Ma, F. Effects of exogenous factors on bacteriocin production from Lactobacillus paracasei J23 by using a resting cell system. Int. J. Mol. Sci. 2013, 14, 24355–24365. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal foodborne diseases: An ongoing challenge in public health. Biomed. Res. Intl. 2014, 2014, 827965. [Google Scholar] [CrossRef] [Green Version]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Brese, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Sofos, J.N. Bacterial foodborne diseases. In Proceedings of the 15th Congress of FAVA. FAVA-OIE Joint Symposium on Emerging Diseases, Bangkok, Thailand, 27–30 October 2008. [Google Scholar]
- Kendall, P. Bacterial Foodborne Illness. Available online: https://extension.colostate.edu/docs/pubs/foodnut/09300.pdf (accessed on 21 November 2021).
- Kuhn, D.M.; Ghannoum, M.A. Indoor mold, toxigenic fungi, and Stachybotrys chartarum: Infectious disease perspective. Clin. Microbiol. Rev. 2003, 16, 144–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukushima, H.; Katsube, K.; Hata, Y.; Kishi, R.; Fujiwara, S. Rapid separation and concentration of foodborne pathogens in food samples prior to quantification by viable-cell counting and real-time PCR. Appl. Env. Microbiol. 2007, 73, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soomro, A.H.; Masud, T.; Anwaar, K. Role of lactic acid bacteria (LAB) in food preservation and human health. Pak. J. Nutr. 2002, 1, 20–24. [Google Scholar]
- Babatunde, D.A.; Oladejo, P.O. Identification of lactic acid bacteria isolated from Nigerian foods. Medical importance and comparison of their bacteriocins activities. J. Nat. Sci. Res. 2014, 4, 76–87. [Google Scholar]
- Kang, D.-K.; Oh, H.K.; Ham, J.-S.; Kim, J.G.; Yoon, C.H.; Ahn, Y.T.; Kim, H.U. Identification and characterization of hydrogen peroxide-generating Lactobacillus fermentum CS12-1. Asian-Aust. J. Anim. Sci. 2005, 18, 90–95. [Google Scholar] [CrossRef]
- Brashears, M.M.; Divya, J. Lactic Acid Bacteria Cultures that Inhibit Foodborne Pathogens. Papers in Veterinary and Biomedical Science. U.S. Patent 7,323,166 B2, 29 January 2008. [Google Scholar]
- Pishva, E.; Hassannia, N.; Fazeli, M.R.; Havaee, A. Antibacterial effect of authochlorous Lactobacillus strains isolated from traditional yoghurts. Pakistan J. Nutr. 2009, 8, 1132–1137. [Google Scholar] [CrossRef]
- Maarof, H.A.; Abdalah, M.I.M.; Bazalou, M.S.; Abo-Samra, R.G. Effect of probiotics bacteria isolated from yoghurts produced in Damietta city on some pathogenic bacteria. In Proceedings of the 6th Scientific Conference of Animal Wealth Research in the Middle East and North Africa, Hurghada, Egypt, 27–30 September 2013; pp. 38–51. [Google Scholar]
- Khandare, S.S.; Patil, S.D. Inhibitory activity of lactic acid bacteria against isolated pathogens and spoilage organisms associated with fresh meat. Int. J. Curr. Microbiol. App. Sci. 2015, 2, 128–135. [Google Scholar]
- Dutra, V.; Silva, A.C.; Cabrita, P.; Peres, C.; Malcata, X.; Brito, L. Lactobacillus plantarum LB95 impairs the virulence potential of Gram-positive and Gram-negative foodborne pathogens in HT-29 and vero cell cultures. J. Med. Microbiol. 2016, 65, 28–35. [Google Scholar] [CrossRef]
- Uraipan, S.; Hongpattarakere, T. Antagonistic characteristics against foodborne pathogenic bacteria of lactic acid bacteria isolated from feces of healthy Thai infants. Jundishapur J. Microbiol. 2015, 8, e18264. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, R.T. Lactic Acid Bacteria as Bioprotective Agents against Foodborne Pathogens and Spoilage Microorganisms in Fresh Fruits and Vegetables. Doctoral Thesis, Universitat de Girona, Girona, Spain, 2008. [Google Scholar]
- Fossi, B.T.; Anyangwe, I.; Tavea, F.; Lucas, K.E.; Nkuo, T.A. Lactic acid bacteria from traditionally processed corn beer and palm wine against selected foodborne pathogens isolated in south west region of Cameroon. Afr. J. Microbiol. Res. 2016, 10, 1140–1147. [Google Scholar]
- Anteneh, T.; Tetemke, M.; Mogessie, A. Antagonism of lactic acid bacteria against foodborne pathogens during storage of borde and shamita, traditional Ethiopian fermented beverages. Int. Food Res. J. 2011, 18, 1189–1194. [Google Scholar]
- Nigam, A.; Kumar, A.; Madhusudan, H.V.; Bhola, N. In-vitro screening of antibacterial activity of lactic acid bacteria against common enteric pathogens. J. Biomed. Sci. 2012, 1. [Google Scholar] [CrossRef]
- Trias, R.; Baneras, L.; Badosa, E.; Montesinos, E. Bioprotection of Golden Delicious apples and Iceberg lettuce against foodborne bacterial pathogens by lactic acid bacteria. Int. J. Food Microbiol. 2008, 123, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Senda, S.; Nomura, N.; Tokuda, H.; Uchiyama, H. Biofilm formation by Lactic Acid Bacteria and resistance to environmental stress. J. Biosci. Bioeng. 2008, 106, 381–386. [Google Scholar] [CrossRef]
- Jalilsood, T.; Baradaran, A.; Song, A.A.; Foo, H.L.; Mustafa, S.; Saad, W.Z.; Yusoff, K.; Rahim, R.A. Inhibition of pathogenic and spoilage bacteria by novel biofilm-forming Lactobacillus isolate: A potential host for the expression of heterologous proteins. Microb. Cell Fact. 2015, 14, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, N.C.; Ramiro, J.M.P.; Quecan, B.X.V.; de Melo Franco, B.D.G. Use of potential probiotic lactic acid bacteria biofilms for the control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coli O157:H7 biofilms formation. Front. Microbiol. 2016, 7, 863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.T.; Xu, H.; Ye, J.Z.; Wu, W.R.; Shi, D.; Fang, D.Q.; Liu, Y.; Li, L.J. Efficacy of Lactobacillus rhamnosus GG in treatment of acute pediatric diarrhea: A systematic review with meta-analysis. World J. Gastroenterol. 2019, 25, 4999–5016. [Google Scholar] [CrossRef] [PubMed]
- Daliri, F.; Aboagye, A.A.; Daliri, E.B.-M. Inactivation of Foodborne Pathogens by Lactic Acid Bacteria. J. Food Hyg. Saf. 2020, 35, 419–429. [Google Scholar] [CrossRef]
- Kim, K.; Lee, G.; Thanh, H.D.; Kim, J.-H.; Konkit, M.; Yoon, S.; Park, M.; Yang, S.; Park, E.; Kim, W. Exopolysaccharide from Lactobacillus plantarum LRCC5310 offers protection against rotavirus-induced diarrhea and regulates inflammatory response. J. Dairy Sc. 2018, 101, 5702–5712. [Google Scholar] [CrossRef]
- Lange-Starke, A.; Petereit, A.; Truyen, U.; Braun, P.G.; Fehlhaber, K.; Albert, T. Antiviral potential of selected starter cultures, bacteriocins and D, L-lactic acid. Food Environ. Virol. 2014, 6, 42–47. [Google Scholar] [CrossRef]
- Aboubakr, H.A.; El-Banna, A.A.; Youssef, M.M.; Al- Sohaimy, S.A.A.; Goyal, S.M. Antiviral effects of Lactococcus lactis on Feline Calicivirus, A Human Norovirus Surrogate. Food Environ. Virol. 2014, 6, 282–289. [Google Scholar] [CrossRef]
- Martín, V.; Maldonado, A.; Fernández, L.; Rodríguez, J.M.; Connor, R.I. Inhibition of human immunodeficiency virus type 1 by lactic acid bacteria from human breastmilk. Breastfeed Med. 2010, 5, 153–158. [Google Scholar] [CrossRef]
- Sirichokchatchawan, W.; Temeeyasen, G.; Nilubol, D.; Prapasarakul, N. Protective effects of cell-free supernatant and live lactic acid bacteria isolated from Thai pigs against a pandemic strain of Porcine Epidemic Diarrhea Virus. Probiotics Antimicro. 2018, 10, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.-C.; MatRahim, N.-A.; AbuBakar, S.; Lee, H.Y. Antilisterial Potential of Lactic Acid Bacteria in Eliminating Listeria monocytogenes in Host and Ready-to-Eat Food Application. Microbiol. Res. 2021, 12, 234–257. [Google Scholar] [CrossRef]
- Gumienna, M.; Górna, B. Antimicrobial Food Packaging with Biodegradable Polymers and Bacteriocins. Molecules 2021, 26, 3735. [Google Scholar] [CrossRef]
- Balciunas, E.M.; Martinez, F.A.C.; Todorov, S.D.; De Melo Franco, B.D.G.; Converti, A.; De Souza Oliveira, R.P. Novel biotechnological applications of bacteriocins: A review. Food Control 2013, 32, 134–142. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Strack, L.; Carli, R.C.; da Silva, R.V.; Sartor, K.B.; Colla, L.M.; Reinehr, C.O. Food biopreservation using antimicrobials produced by lactic acid bacteria. Res. Soc. Dev. 2020, 9, e998986666. [Google Scholar] [CrossRef]
- Neetoo, H.; Ye, M.; Chen, H. Potential antimicrobials to control Listeria monocytogenes in vacuum-packaged cold-smoked salmon pâté and fillets. Int. J. Food Microbiol. 2008, 123, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Woraprayote, W.; Pumpuang, L.; Tosukhowong, A.; Zendo, T.; Sonomoto, K.; Benjakul, S.; Visessanguan, W. Antimicrobial biodegradable food packaging impregnated with bacteriocin 7293 for control of pathogenic bacteria in pangasius fish fillets. LWT—Food Sc. Tech 2018, 89, 427–433. [Google Scholar] [CrossRef]
- Cao-Hoang, L.; Chaine, A.; Grégoire, L.; Waché, Y. Potential of nisin-incorporated sodium caseinate films to control Listeria in artificially contaminated cheese. Food Microbiol. 2010, 27, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.D.; Funck, G.D.; Dannenberg, G.D.; Cruxen, C.E.D.; ElHalal, S.L.M.; Dias, A.R.G. Bacteriocin-like substances of Lactobacillus curvatus P99: Characterization and application in biodegradable films for control of Listeria monocytogenes in cheese. Food Microbiol. 2017, 63, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Angiolillo, L.; Conte, A.; Del Nobile, M.A. A new method to bio-preserve sea bass fillets. Int. J. Food Microbiol. 2018, 271, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.C.R.; Alvarenga, V.O.; Thomazini, M.; Favaro-Trindade, C.S.; Sant’Ana, A.D. Assessment of the inhibitory effect of free and encapsulated commercial nisin (Nisaplin (R)), tested alone and in combination, on Listeria monocytogenes and Bacillus cereus in refrigerated milk. LWT-Food Sci. Technol. 2016, 68, 67–75. [Google Scholar] [CrossRef]
- Narayanan, A.; Mallesha, N.; Ramana, K.V. Synergized antimicrobial activity of eugenol incorporated polyhydroxybutyrate films against food spoilage microorganisms in conjunction with pediocin. Appl. Biochem. Biotech. 2013, 170, 1379–1388. [Google Scholar] [CrossRef]
- Zapico, P.; Medina, M.; Gaya, P.; Nuñez, M. Synergistic effect of nisin and the lactoperoxidase system on Listeria monocytogenes in skim milk. Int. J. Food Microbiol. 1998, 40, 35–42. [Google Scholar] [CrossRef]
- Branen, J.K.; Davidson, M.P. Enhancement of nisin, lysozyme, and monolaurin antimicrobial activities by ethylenediaminetetraacetic acid and lactoferrin. Int. J. Food Microbiol. 2004, 90, 63–74. [Google Scholar] [CrossRef]
- Prudêncio, C.V.; Dos Santos, M.T.; Vanetti, M.C.D. Strategies for the use of bacteriocins in Gram-negative bacteria: Relevance in food microbiology. J. Food Sci. Technol. 2015, 52, 5408–5417. [Google Scholar] [CrossRef] [Green Version]
- Masschalck, B.; Garcia-Graells, C.; Haver, E.V.; Michiels, C.W. Inactivation of high pressure resistant Escherichia coli by lysozyme and nisin under high pressure. Innov. Food Sci. Emerg. 2000, 1, 39–47. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, S.; Liu, F.; Dong, P.; Huang, W.; Xiong, L. Comparing the effects of high hydrostatic pressure and thermal pasteurization combined with nisin on the quality of cucumber juice drinks. Innov. Food Sci. Emerg. Technol. 2013, 17, 27–36. [Google Scholar] [CrossRef]
- Renye, J.A.; Somkuti, G. Bacteriocins of Food Grade Lactic Acid Bacteria in Hurdle Technology for Milk and Dairy Products. In Emerging Dairy Processing Technologies: Opportunities for the Dairy Industry; Datta, N., Tomasula, P.M., Eds.; JohnWiley & Sons: Hoboken, NJ, USA, 2015; pp. 267–306. [Google Scholar]
- Turgis, M.; Vu, K.D.; Dupont, C.; Lacroix, M. Combined antimicrobial effect of essential oils and bacteriocins against foodborne pathogens and food spoilage bacteria. Food Res. Int. 2012, 48, 696–702. [Google Scholar] [CrossRef]
- Govaris, A.; Solomakos, N.; Pexara, A.; Chatzopoulou, P.S. The antimicrobial effect of oregano essential oil, nisin and their combination against Salmonella Enteritidis in minced sheep meat during refrigerated storage. Int. J. Food Microbiol. 2010, 137, 175–180. [Google Scholar] [CrossRef]
- Moosavy, M.H.; Basti, A.A.; Misaghi, A.; Salehi, T.Z.; Abbasifar, R.; Mousavi, H.A.E.; Alipour, M.; Razavi, N.E.; Gandomi, H.; Noori, N. Effect of Zataria multiflora Boiss. essential oil and nisin on Salmonella Typhimurium and Staphylococcus aureus in a food model system and on the bacterial membranes. Food Res. Int. 2018, 41, 1050–1057. [Google Scholar] [CrossRef]
- Mahantesha, T.; Reddy, K.M.P.; Kumar, N.H.P.; Nara, A.; Ashwin, D.; Buddiga, V. Comparative study of probiotic ice cream and probiotic drink on salivary Streptococcus mutans levels in 6-12 years age group children. J. Int. Oral Health 2015, 7, 47–50. [Google Scholar]
- Yoo, J.Y.; Kim, S.S. Probiotics and prebiotics: Present status and future perspectives on metabolic disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, N.; Okamura, K.; Zendo, T.; Himeno, T.; Nakayama, J.; Sonomoto, K. Identification and characterization of novel bacteriocins produced by Enterococcus mundtii QU 2 isolated from soybean. J. Appl. Microbiol. 2005, 99, 1181–1190. [Google Scholar]

{kind=link}
| Classes | Source | Biochenical Profiles | Examples |
|---|---|---|---|
| I | Lactobacillus lactis subsp. Lactis | Have lanthionine and methyllanthionine; <5 kDa | Nisin [14,16] |
| IIa | Leuconostoc gelidum | Thermostable, non-modified, cationic, hydrophobic peptides; <10 kDa | Leucocin A [16,26,33] |
| IIb | Enterococcus faecium | Cationic peptide pairs | Enterocin X [28,29] |
| III | Lactobacillus helveticus | Large peptides; heat-labile; >30 kDa | Helveticin J [14,16,28] |
| LAB Species | Spectrum of Action | References |
|---|---|---|
| L. casei | E. coli; Salmonella spp. | [53] |
| L. plantarum; L. paraplantarum | Listeria spp.; Salmonella spp.; Escherichia spp.; Aeromonas hydrophila; B. cereus; P. fluorescens | [51,52,53] |
| L. sake | Listeria monocytogenes; Leuconostoc spp.; Pediococcus spp. | [9] |
| Leuconostoc mesenteroides | Enterococcus faecalis; Listeria monocytogenes | [9] |
| Pediococcus pentosaceous | Listeria spp.; Clostridium spp. | [9] |
| Enterococcus faecium | Listeria monocytogenes; Pediococcus spp. | [9] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokoena, M.P.; Omatola, C.A.; Olaniran, A.O. Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens. Molecules 2021, 26, 7055. https://doi.org/10.3390/molecules26227055
Mokoena MP, Omatola CA, Olaniran AO. Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens. Molecules. 2021; 26(22):7055. https://doi.org/10.3390/molecules26227055
Chicago/Turabian StyleMokoena, Mduduzi P., Cornelius A. Omatola, and Ademola O. Olaniran. 2021. "Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens" Molecules 26, no. 22: 7055. https://doi.org/10.3390/molecules26227055
APA StyleMokoena, M. P., Omatola, C. A., & Olaniran, A. O. (2021). Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens. Molecules, 26(22), 7055. https://doi.org/10.3390/molecules26227055