
The Inducible Intein-Mediated Self-Cleaving Tag (IIST) System: A Novel Purification and Amidation System for Peptides and Proteins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
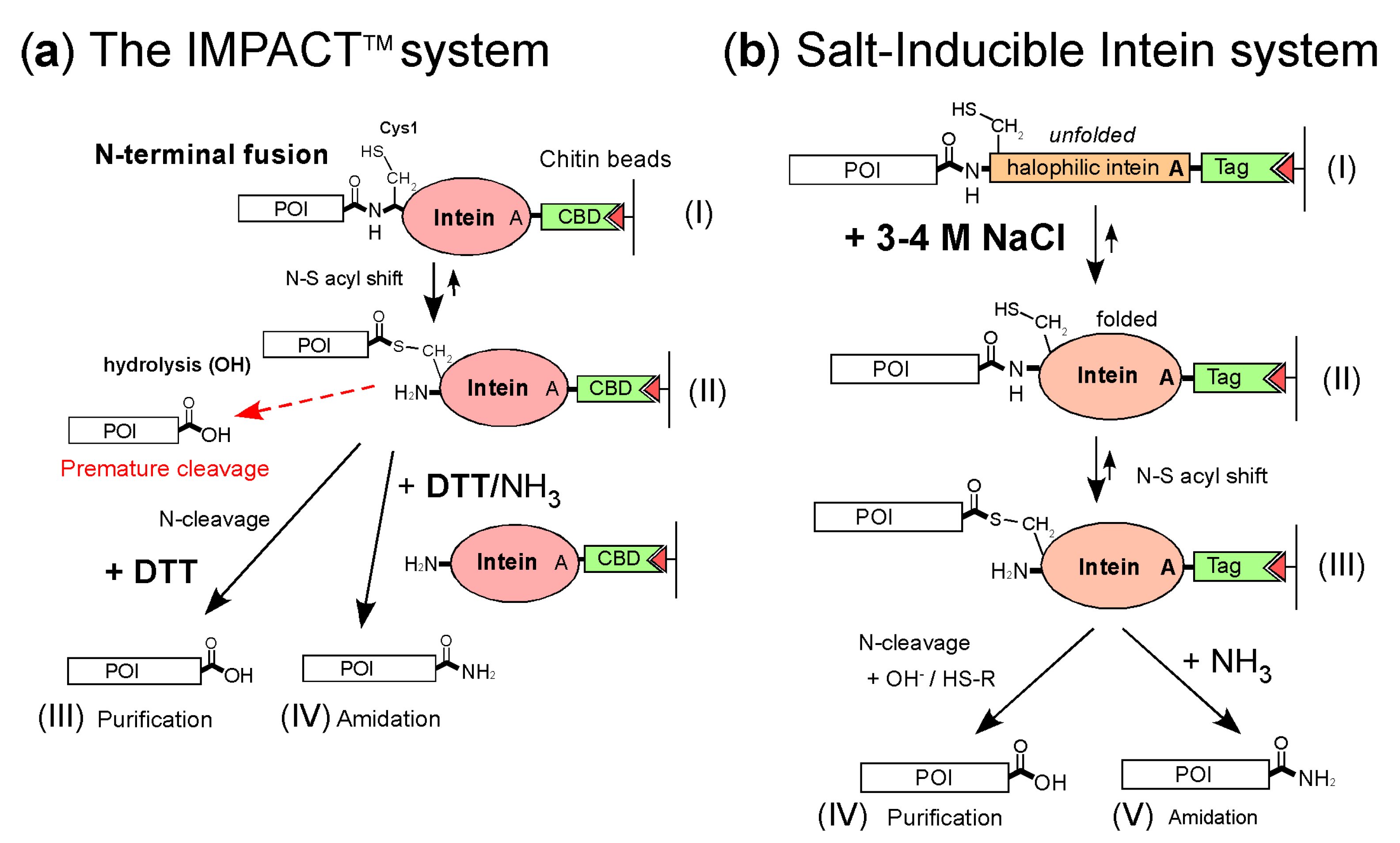
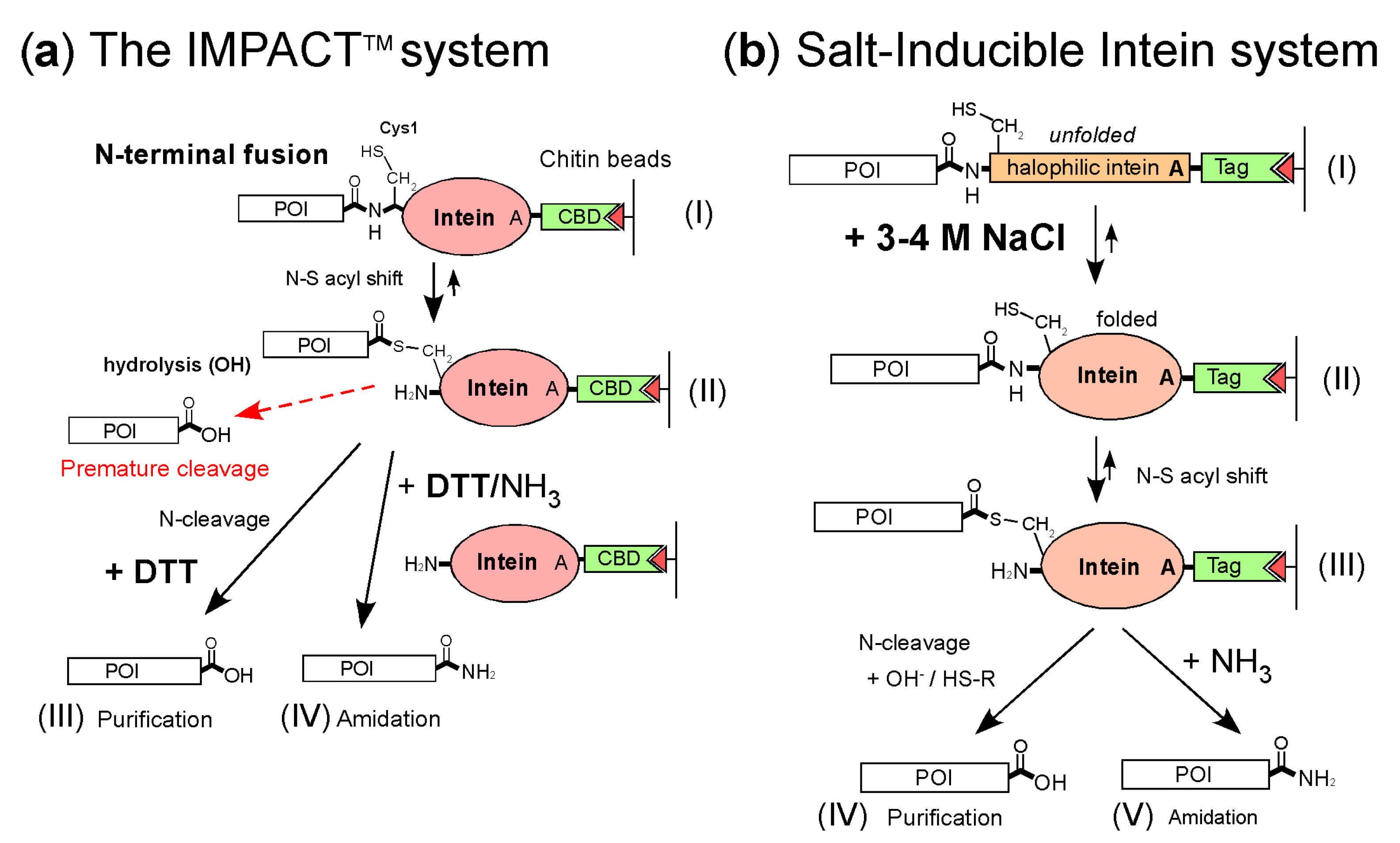
:1. Introduction
2. Results
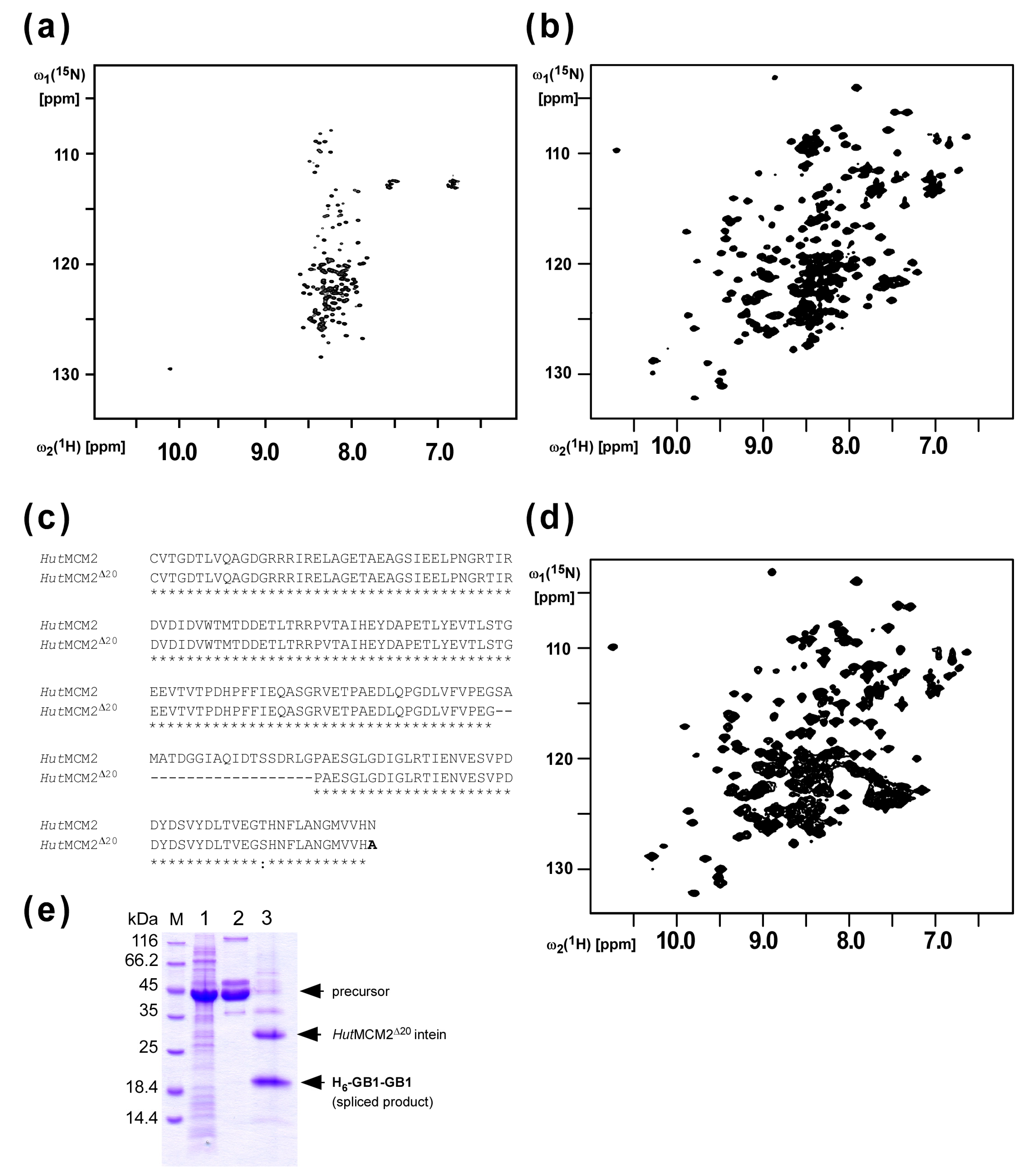
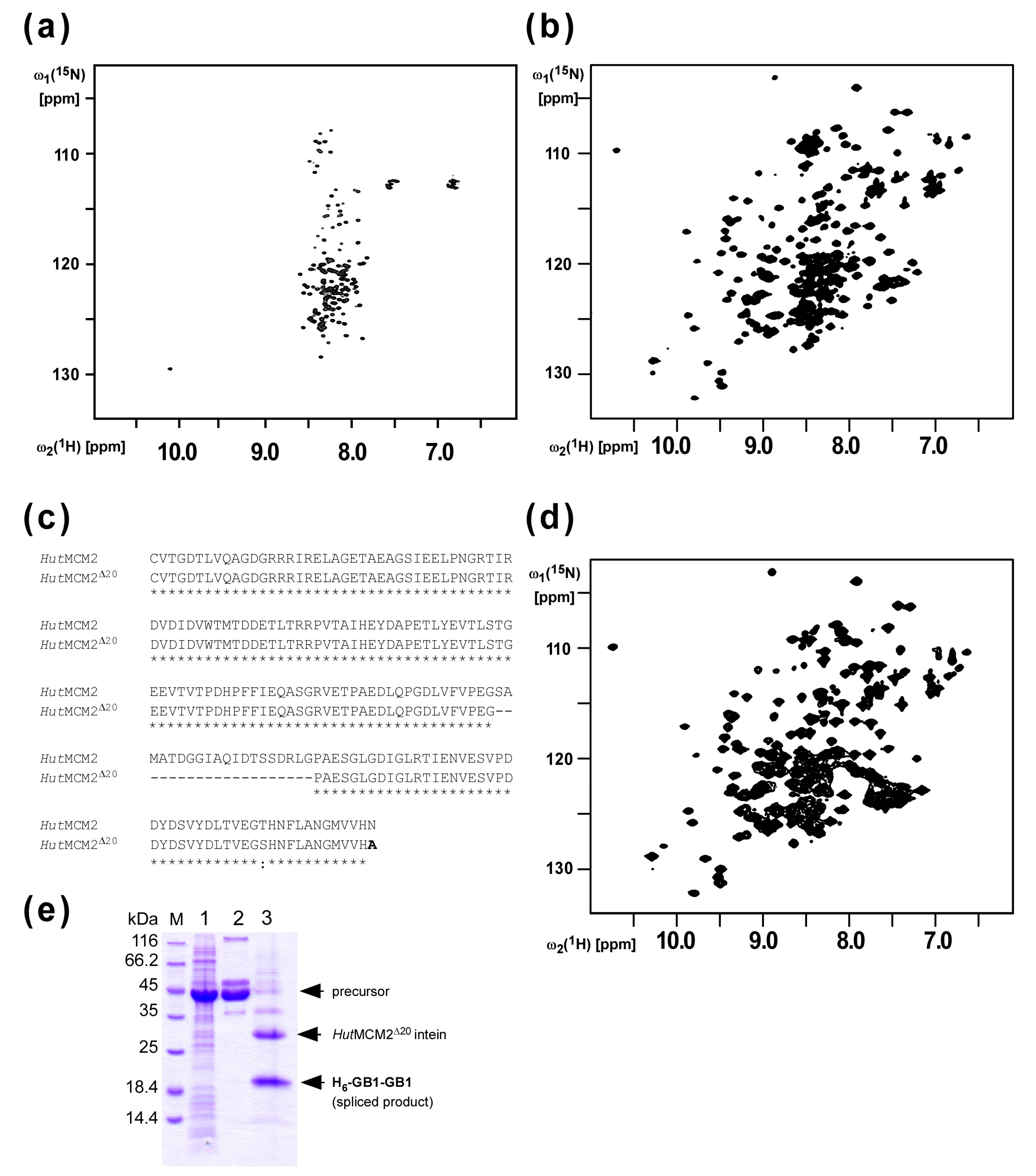
2.1. Design of the Minimized HutMCM2 Intein
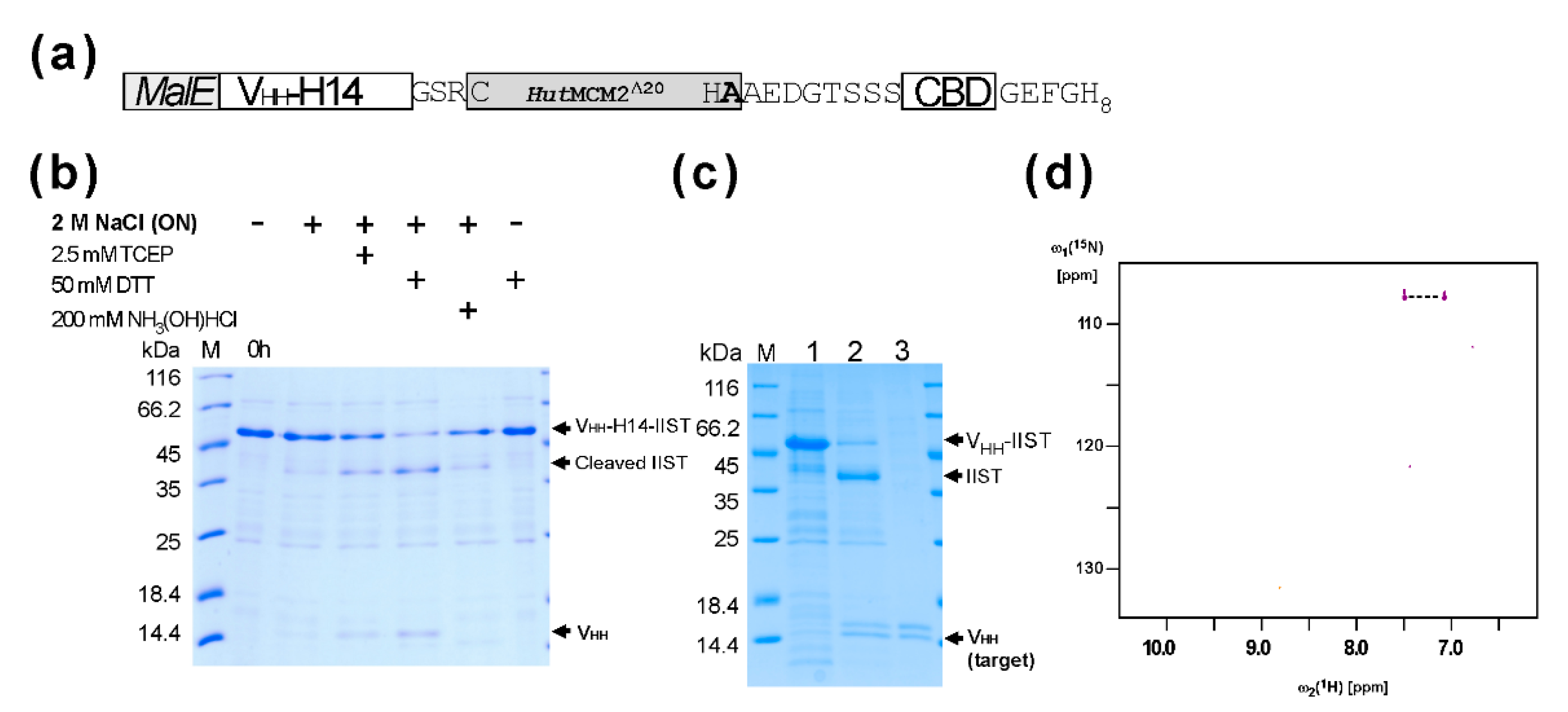
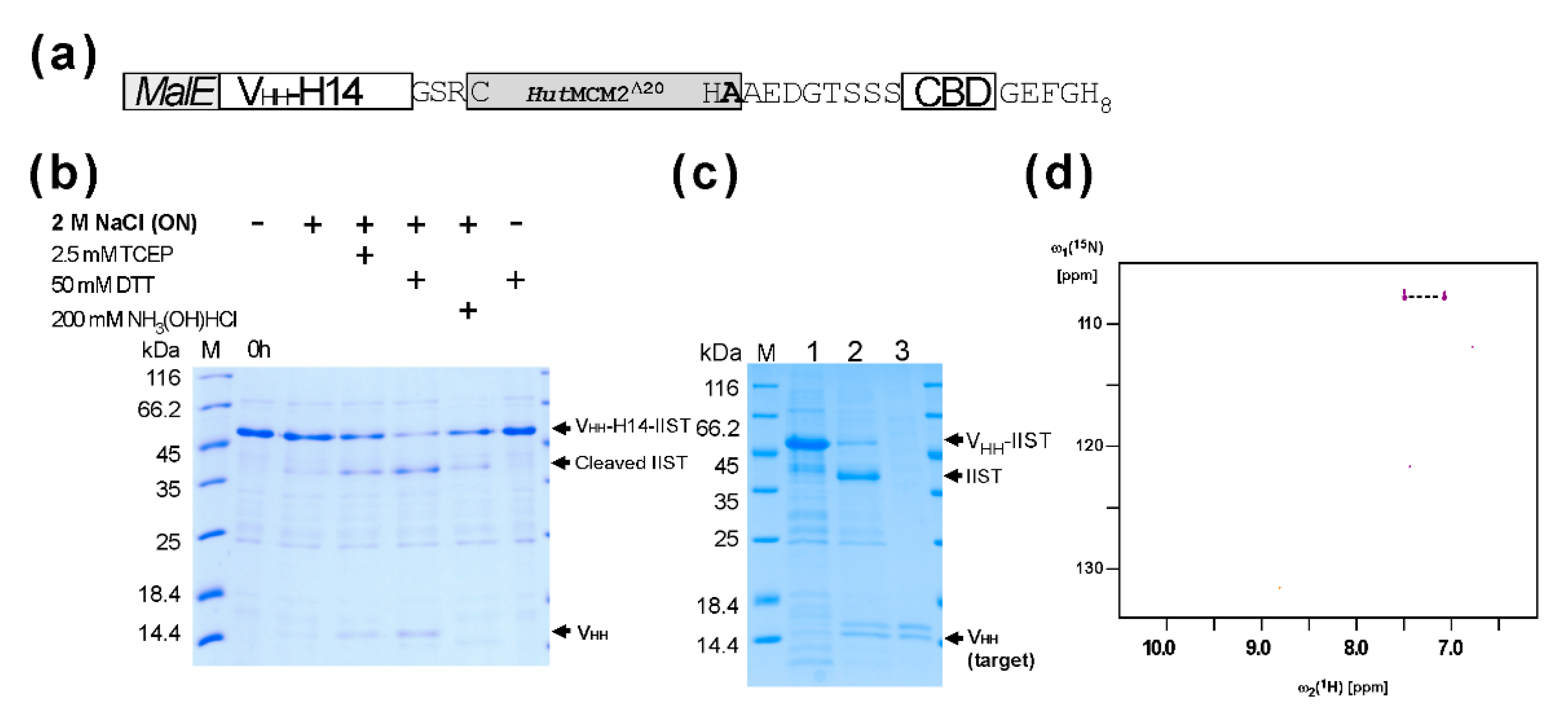
2.2. Purification and the C-Terminal Amidation of a Single Domain Antibody (VHH) with IIST
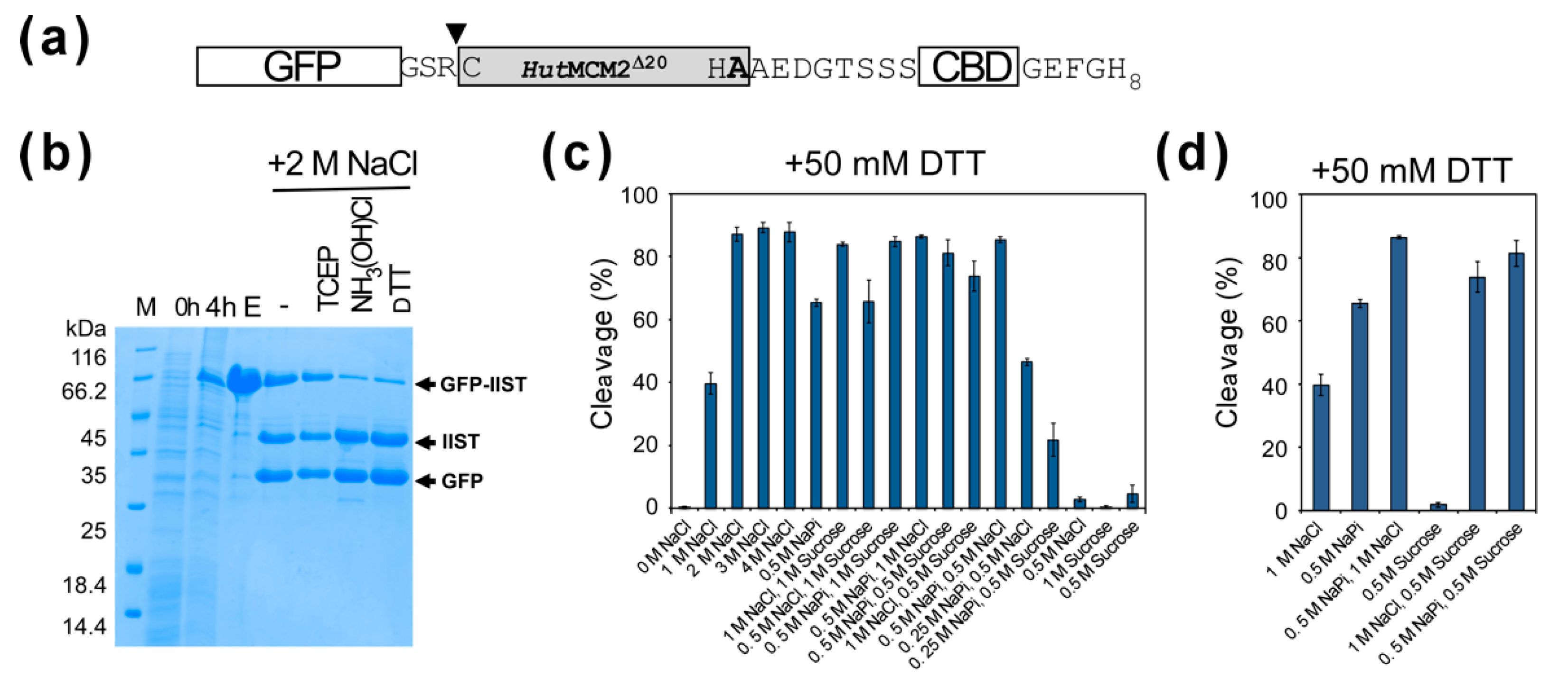
2.3. Comparison of Cleavage Conditions with a Model Protein, GFP
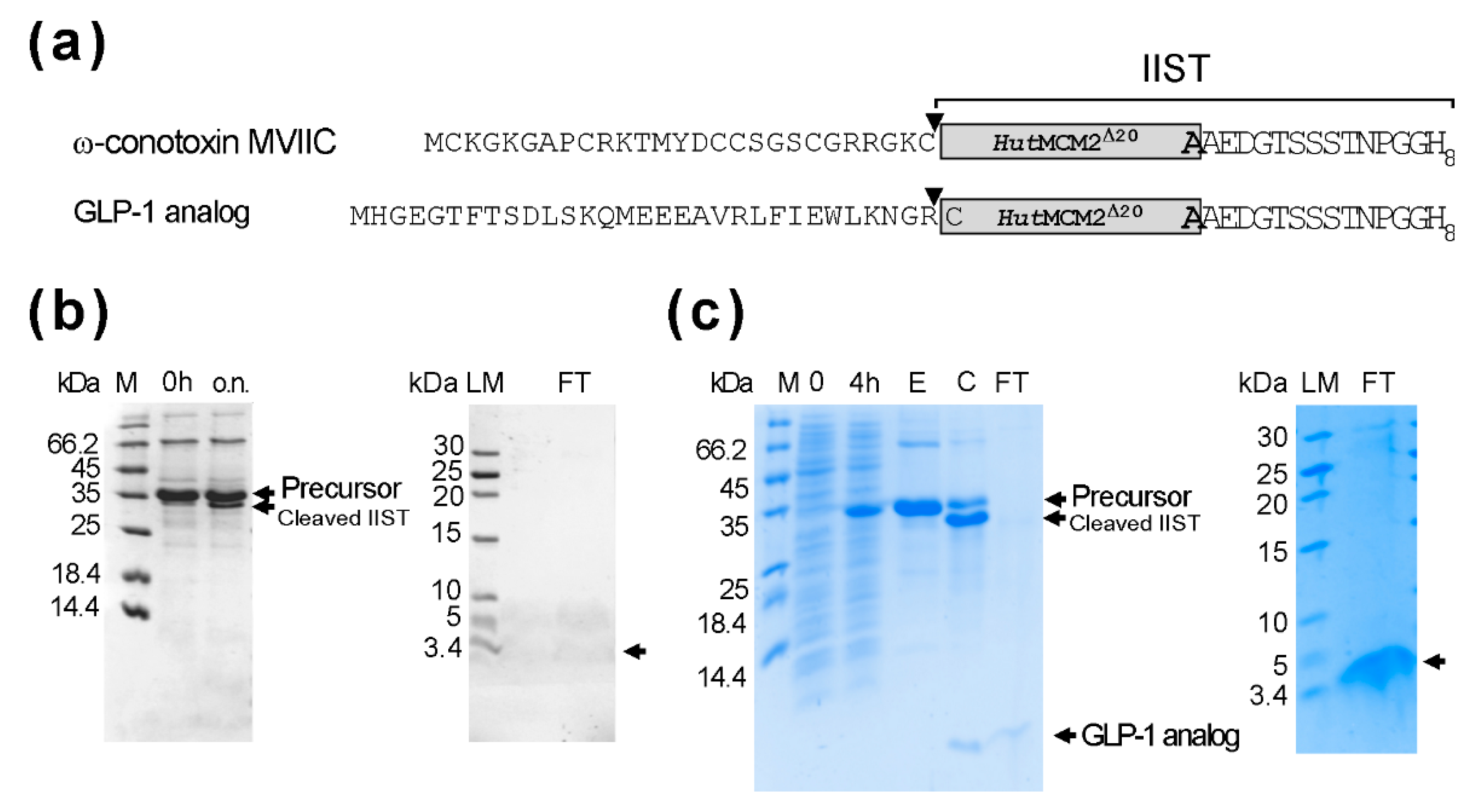
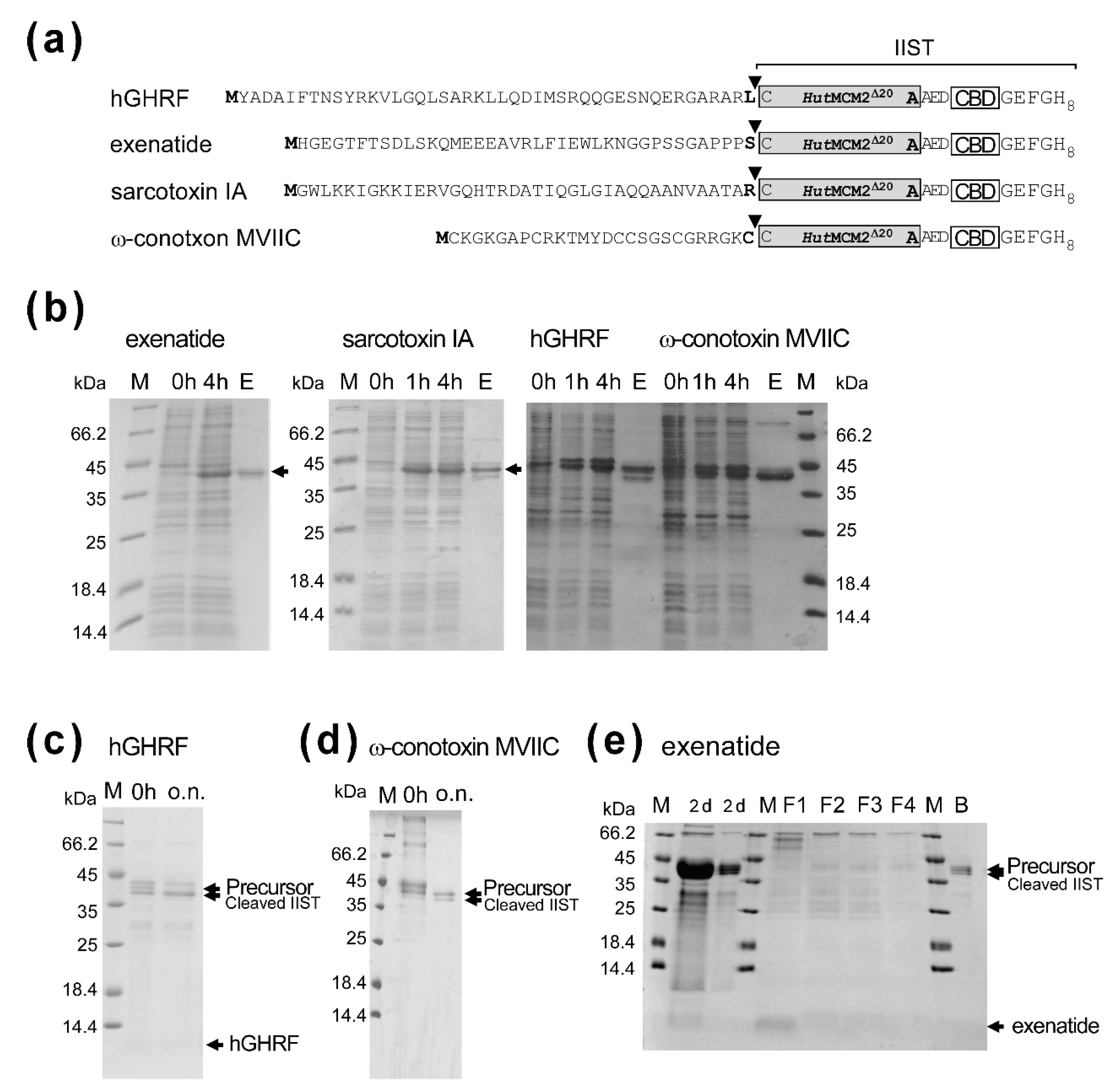
2.4. Applications of IIST with Small Peptides
2.5. Optimization of IIST
3. Discussion
4. Materials and Methods
4.1. Constructions of Plasmids for Protein Expression
4.2. Protein Expression and Purification
4.3. Purification and the C-Terminal Amidation of VHH-H14
4.4. NMR Spectroscopy
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References and Notes
- Eipper, B.A.; Stoffers, D.A.; Mains, R.E. The Biosynthesis of Neuropeptides: Peptide alpha-Amidation. Annu. Rev. Neurosci. 1992, 15, 57–85. [Google Scholar] [CrossRef]
- Merkler, D.J. C-Terminal amidated peptides: Production by the in vitro enzymatic amidation of glycine-extended peptides and the importance of the amide to bioactivity. Enzym. Microb. Technol. 1994, 16, 450–456. [Google Scholar] [CrossRef]
- Chong, S.; Mersha, F.B.; Comb, D.G.; E Scott, M.; Landry, D.; Vence, L.M.; Perler, F.B.; Benner, J.; Kucera, R.B.; A Hirvonen, C.; et al. Single-column purification of free recombinant proteins using a self-cleavable affinity tag derived from a protein splicing element. Gene 1997, 192, 271–281. [Google Scholar] [CrossRef]
- Southworth, M.W.; Amaya, K.; Evans, T.C.; Xu, M.-Q.; Perler, F.B. Purification of proteins fused to either the amino or carboxy terminus of the mycobacterium xenopi gyrase a intein. Biotechniques 1999, 27, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Instruction manual, IMPACT™ Kit (NEB #E6901, New England Biolabs).
- Wood, D.W.; Wu, W.; Belfort, G.; Derbyshire, V.; Belfort, M. A genetic system yields self-cleaving inteins for bioseparations. Nat. Biotechnol. 1999, 17, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Miozzi, J.M.; Stimple, S.D.; Han, T.-C.; Wood, D.W. Column-free purification methods for recombinant proteins using self-cleaving aggregating tags. Polymers 2018, 10, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottingham, I.R.; Millar, A.; Emslie, E.; Colman, A.; Schnieke, A.E.; McKee, C. A method for the amidation of recombinant peptides expressed as intein fusion proteins in Escherichia coli. Nat. Biotechnol. 2001, 19, 974–977. [Google Scholar] [CrossRef] [PubMed]
- Minato, Y.; Ueda, T.; Machiyama, A.; Shimada, I.; Iwaï, H. Segmental isotopic labeling of a 140 kDa dimeric multi-domain protein CheA from Escherichia coli by expressed protein ligation and protein trans-splicing. J. Biomol. NMR 2012, 53, 191–207. [Google Scholar] [CrossRef] [Green Version]
- Guan, D.; Ramirez, M.; Chen, Z. Split intein mediated ultra-rapid purification of tagless protein (SIRP). Biotechnol. Bioeng. 2013, 110, 2471–2481. [Google Scholar] [CrossRef]
- Cooper, M.A.; Taris, J.E.; Shi, C.; Wood, D.W. A convenient split-intein tag method for the purification of tagless target proteins. Curr. Protoc. Protein Sci. 2018, 91, 5.29.1–5.29.23. [Google Scholar] [CrossRef] [PubMed]
- Ciragan, A.; Aranko, A.S.; Tascón, I.; Iwaï, H. Salt-inducible protein splicing in cis and trans by inteins from extremely halophilic archaea as a novel protein-engineering tool. J. Mol. Biol. 2016, 428, 4573–4588. [Google Scholar] [CrossRef]
- Sakakibara, N.; Han, M.; Rollor, C.R.; Gilson, R.C.; Busch, C.; Heo, G.; Kelman, Z. Cloning, purification, and partial characterization of the halobacterium sp. NRC-1 minichromosome maintenance (MCM) helicase. Open Microbiol. J. 2008, 2, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitter, J.N.; Cousin, C.E.; Nicastri, M.C.; Jaramillo, M.V.; Mills, K.V. Salt-dependent conditional protein splicing of an intein from halobacterium salinarum. Biochemistry 2016, 55, 1279–1282. [Google Scholar] [CrossRef]
- Di Ventura, B.; Mootz, H.D. Switchable inteins for conditional protein splicing. Biol. Chem. 2019, 400, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Ciragan, A.; Backlund, S.M.; Mikula, K.M.; Beyer, H.; Ollila, O.H.S.; Iwaï, H. NMR structure and dynamics of tonb investigated by scar-less segmental isotopic labeling using a salt-inducible split intein. Front. Chem. 2020, 8, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeemig, J.S.; Aranko, A.S.; Djupsjöbacka, J.; Heinämäki, K.; Iwaï, H. Solution structure of DnaE intein fromNostoc punctiforme: Structural basis for the design of a new split intein suitable for site-specific chemical modification. FEBS Lett. 2009, 583, 1451–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinelli, S.; Frenken, L.; Bourgeois, D.; De Ron, L.; Bos, W.; Verrips, T.; Anguille, C.; Cambillau, C.; Tegoni, M. The crystal structure of a llama heavy chain variable domain. Nat. Struct. Mol. Biol. 1996, 3, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Renisio, J.-G.; Pérez, J.; Czisch, M.; Guenneugues, M.; Bornet, O.; Frenken, L.; Cambillau, C.; Darbon, H. Solution structure and backbone dynamics of an antigen-free heavy chain variable domain (VHH) fromLlama. Proteins Struct. Funct. Bioinform. 2002, 47, 546–555. [Google Scholar] [CrossRef]
- Fukuchi, S.; Yoshimune, K.; Wakayama, M.; Moriguchi, M.; Nishikawa, K. Unique amino acid composition of proteins in halophilic bacteria. J. Mol. Biol. 2003, 327, 347–357. [Google Scholar] [CrossRef]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.; Dutta, C. Molecular signature of hypersaline adaptation: Insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [Green Version]
- Tadeo, X.; López-Méndez, B.; Trigueros, T.; Laín, A.; Castaño, D.; Millet, O. Structural basis for the aminoacid composition of proteins from halophilic archea. PLoS Biol. 2009, 7, e1000257. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Timasheff, S. The stabilization of proteins by sucrose. J. Biol. Chem. 1981, 256, 7193–7201. [Google Scholar] [CrossRef]
- Guillemin, R.; Brazeau, P.; Bohlen, P.; Esch, F.; Ling, N.; Wehrenberg, W. Growth hormone-releasing factor from a human pancreatic tumor that caused acromegaly. Science 1982, 218, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Nakajima, Y.; Natori, S.; Arata, Y.; Shimada, I. Solution conformation of an antibacterial peptide, sarcotoxin IA, as determined by 1H-NMR. JBIC J. Biol. Inorg. Chem. 1993, 217, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; A Nauck, M. The incretin system: Glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- McDonough, S.I.; Swartz, K.J.; Mintz, I.M.; Boland, L.; Bean, B.P. Inhibition of calcium channels in rat central and peripheral neurons by omega-conotoxin MVIIC. J. Neurosci. 1996, 16, 2612–2623. [Google Scholar] [CrossRef]
- Iwai, H.; Züger, S.; Jin, J.; Tam, P.-H. Highly efficient proteintrans-splicing by a naturally split DnaE intein fromNostoc punctiforme. FEBS Lett. 2006, 580, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Oeemig, J.S.; Beyer, H.M.; Aranko, A.S.; Mutanen, J.; Iwaï, H. Substrate specificities of inteins investigated by QuickDrop-cassette mutagenesis. FEBS Lett. 2020, 594, 3338–3355. [Google Scholar] [CrossRef]
- Schägger, H. Tricine–SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef]
- Arakawa, T.; Timasheff, S. The stabilization of proteins by osmolytes. Biophys. J. 1985, 47, 411–414. [Google Scholar] [CrossRef]
- Novikova, O.; Jayachandran, P.; Kelley, D.S.; Morton, Z.; Merwin, S.; Topilina, N.I.; Belfort, M. Intein clustering suggests functional importance in different domains of life. Mol. Biol. Evol. 2016, 33, 783–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, F.; Ciragan, A.; Iwaï, H. Tandem SUMO fusion vectors for improving soluble protein expression and purification. Protein Expr. Purif. 2015, 116, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Vranken, W.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins Struct. Funct. Bioinform. 2005, 59, 687–696. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aranko, A.S.; Iwaï, H. The Inducible Intein-Mediated Self-Cleaving Tag (IIST) System: A Novel Purification and Amidation System for Peptides and Proteins. Molecules 2021, 26, 5948. https://doi.org/10.3390/molecules26195948
Aranko AS, Iwaï H. The Inducible Intein-Mediated Self-Cleaving Tag (IIST) System: A Novel Purification and Amidation System for Peptides and Proteins. Molecules. 2021; 26(19):5948. https://doi.org/10.3390/molecules26195948
Chicago/Turabian StyleAranko, A. Sesilja, and Hideo Iwaï. 2021. "The Inducible Intein-Mediated Self-Cleaving Tag (IIST) System: A Novel Purification and Amidation System for Peptides and Proteins" Molecules 26, no. 19: 5948. https://doi.org/10.3390/molecules26195948
APA StyleAranko, A. S., & Iwaï, H. (2021). The Inducible Intein-Mediated Self-Cleaving Tag (IIST) System: A Novel Purification and Amidation System for Peptides and Proteins. Molecules, 26(19), 5948. https://doi.org/10.3390/molecules26195948