
Pharmacological and Therapeutic Potential of Myristicin: A Literature Review


Abstract
1. Introduction
2. Results and Discussion
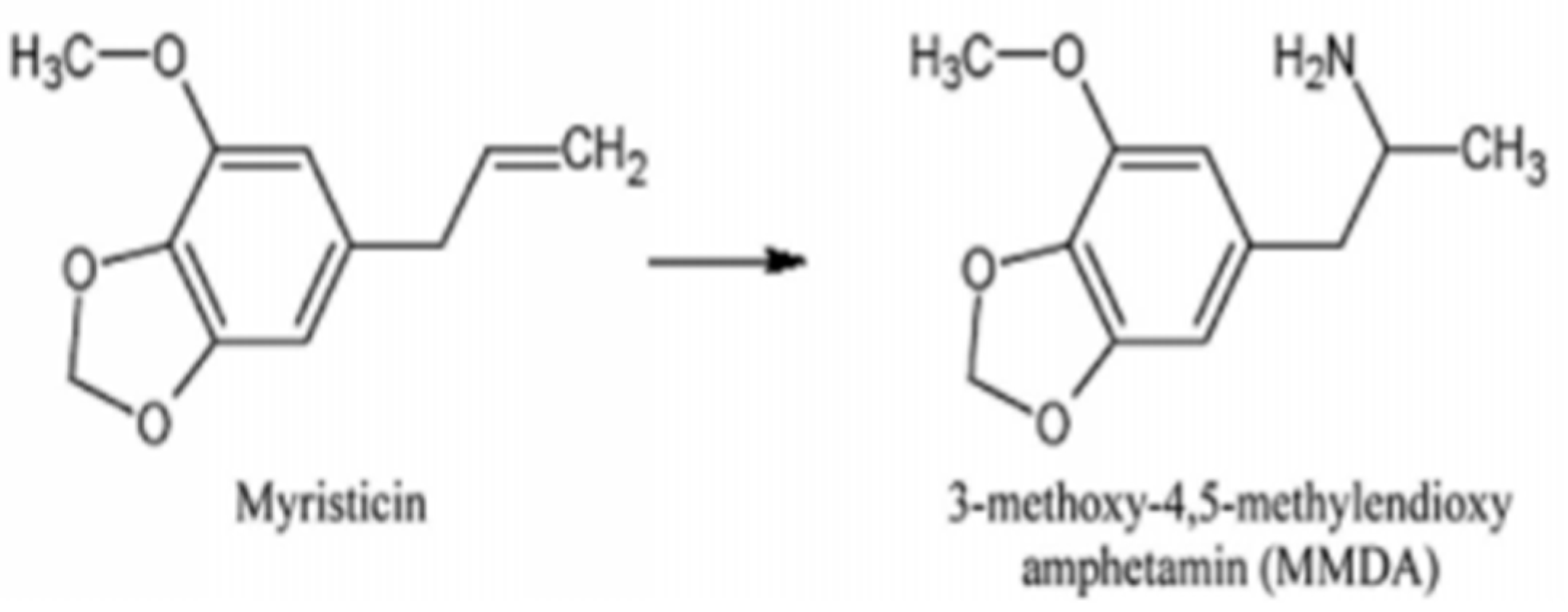
2.1. Metabolization and Toxicity of Myristicin
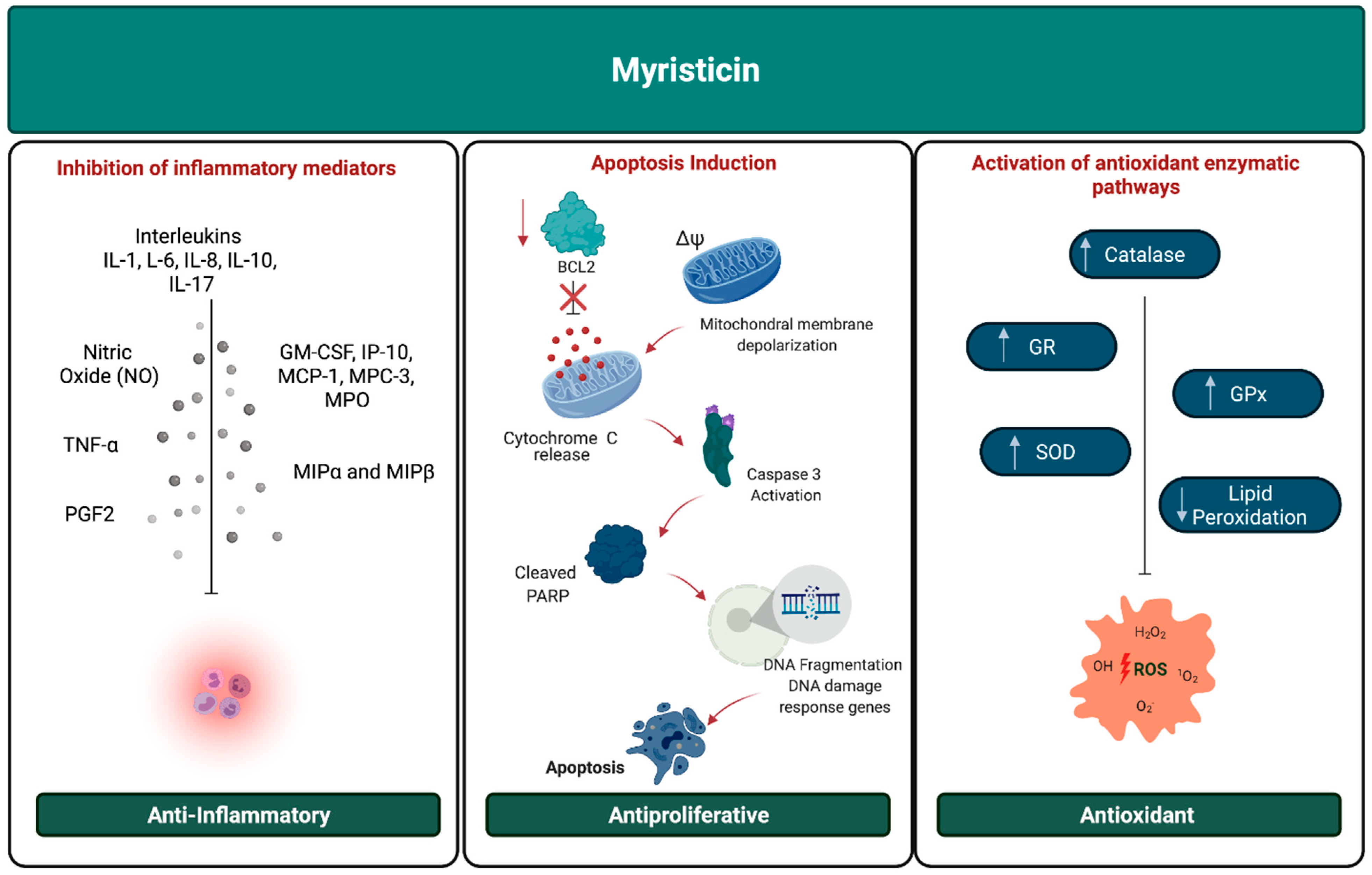
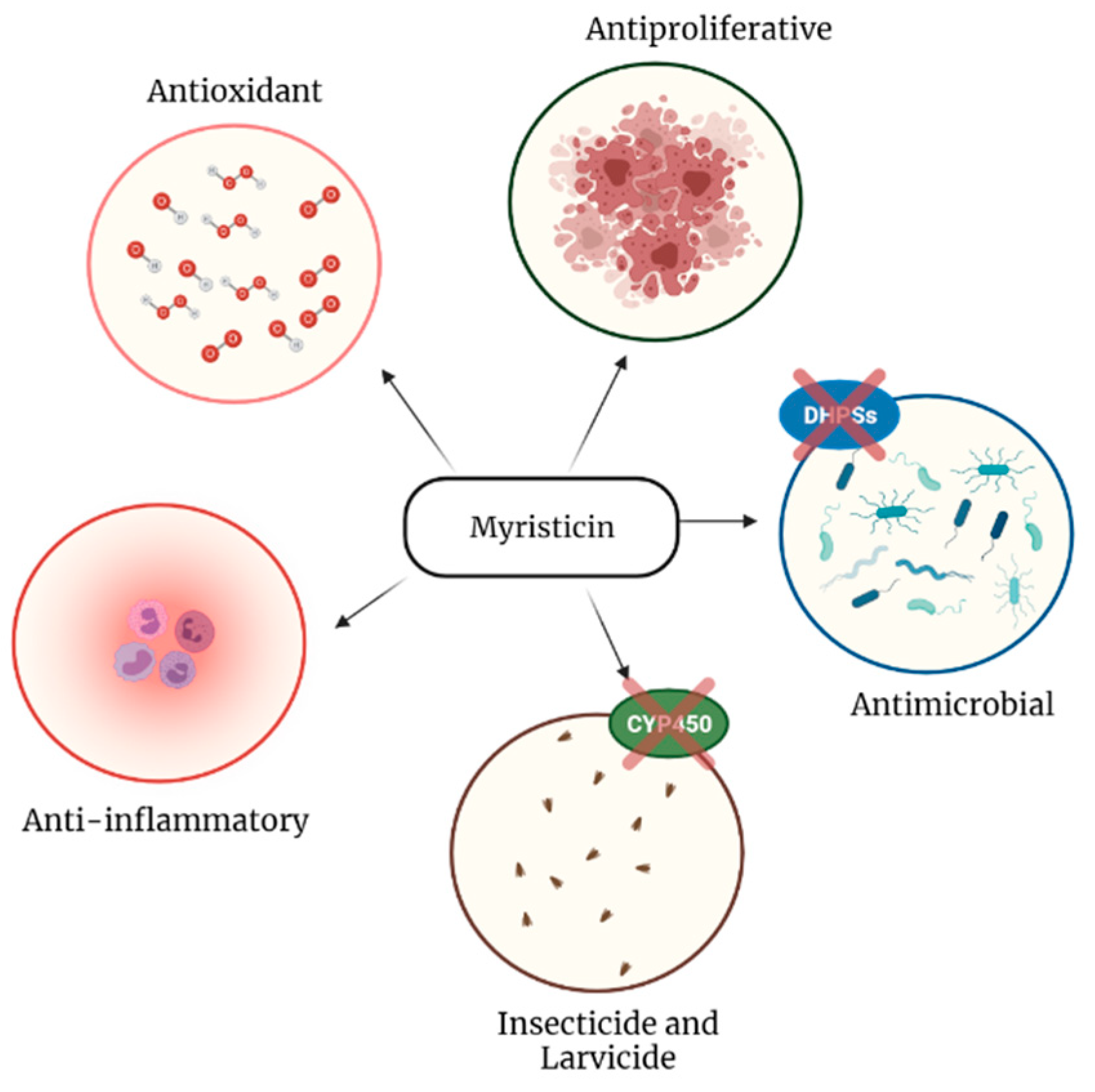
2.2. Antioxidant Activity
2.3. Anti-Inflammatory and Analgesic Activity
2.4. Antiproliferative Activity
2.5. Antimicrobial Activity
2.6. Insecticide and Larvicide Activity
2.7. Other Activities
2.8. Future Perspectives
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Katz, L.; Baltz, R.H. Natural Product Discovery: Past, Present, and Future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural Products: A Continuing Source of Novel Drug Leads. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3670–3695. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from January 1981 to September 2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Ligon, B.L. Penicillin: Its Discovery and Early Development. Semin. Pediatr. Infect. Dis. 2004, 15, 52–57. [Google Scholar] [CrossRef]
- Da Silva Prade, J.; Bálsamo, E.C.; Machado, F.R.; Poetini, M.R.; Bortolotto, V.C.; Araújo, S.M.; Londero, L.; Boeira, S.P.; Sehn, C.P.; de Gomes, M.G.; et al. Anti-Inflammatory Effect of Arnica Montana in a UVB Radiation-Induced Skin-Burn Model in Mice. Cutan. Ocul. Toxicol. 2020, 39, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Brixius, D. A Hard Nut to Crack: Nutmeg Cultivation and the Application of Natural History between the Maluku Islands and Isle de France (1750s–1780s). Br. J. Hist. Sci. 2018, 51, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, P.; Kumar, P.; Singh, V.K.; Singh, D.K. Biological Effects of Myristica fragrans. Annu. Rev. Biomed. Sci. 2009, 11, 21–29. [Google Scholar] [CrossRef]
- Torbati, M.; Nazemiyeh, H.; Lotfipour, F.; Asnaashari, S.; Nemati, M.; Fathiazad, F. Composition and Antibacterial Activity of Heracleum transcaucasicum and Heracleum anisactis Aerial Parts Essential Oil. Adv. Pharm. Bull. 2013, 3, 415–418. [Google Scholar] [CrossRef][Green Version]
- Yang, A.H.; He, X.; Chen, J.X.; He, L.N.; Jin, C.H.; Wang, L.L.; Zhang, F.L.; An, L.J. Identification and Characterization of Reactive Metabolites in Myristicin-Mediated Mechanism-Based Inhibition of CYP1A2. Chem. Biol. Interact. 2015, 237, 133–140. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, Y.K.; Yang, X.N.; Xiao, X.R.; Zhang, T.; Yang, X.W.; Qin, H.B.; Li, F. Metabolic Activation of Myristicin and Its Role in Cellular Toxicity. J. Agric. Food Chem. 2019, 67, 4328–4336. [Google Scholar] [CrossRef]
- Duarte, D.; Mendonça, M.; Ramos, L. O Poder Psicoativo Dos Alimentos: Análise de Um Caso de Intoxicação Com Noz-Moscada. Acta Med. Port. 2021, 34, 298. [Google Scholar] [CrossRef] [PubMed]
- Leiter, E.; Hitchcock, G.; Godwin, S.; Johnson, M.; Sedgwick, W.; Jones, W.; McCall, S.; Ceremuga, T.E. Evaluation of the Anxiolytic Properties of Myristicin, a Component of Nutmeg, in the Male Sprague-Dawley Rat. AANA J. 2011, 79, 109–114. [Google Scholar] [PubMed]
- Rahman, N.A.A.; Fazilah, A.; Effarizah, M.E. Toxicity of Nutmeg (Myristicin): A Review. Int. J. Adv. Sci. Eng. Inf. Technol. 2015, 5, 212–215. [Google Scholar] [CrossRef]
- Sivathanu, S.; Sampath, S.; David, H.S.; Rajavelu, K.K. Myristicin and Phenytoin Toxicity in an Infant. BMJ Case Rep. 2014, 1, 2013–2015. [Google Scholar] [CrossRef] [PubMed]
- Cushy, A.R. Therapeutical and pharmacological section: Nutmeg poisoning. Proc. R. Soc. Med. 1908, 1, 39–44. [Google Scholar]
- Badr, G.; Elsawy, H.; Malki, M.A.; Alfwuaires, M.; El-Gerbed, M.S.A.; Abdel-Moneim, A.M. Protective Effects of Myristicin against Ulcerative Colitis Induced by Acetic Acid in Male Mice. Food Agric. Immunol. 2020, 31, 435–446. [Google Scholar] [CrossRef]
- Amiri, H. Volatile Constituents and Antioxidant Activity of Flowers, Stems and Leaves of Nasturtium Officinale R. Br. Nat. Prod. Res. 2012, 26, 109–115. [Google Scholar] [CrossRef]
- Boulebd, H. DFT Study of the Antiradical Properties of Some Aromatic Compounds Derived from Antioxidant Essential Oils: C–H Bond vs. O–H Bond. Free Radic. Res. 2019, 53, 1125–1134. [Google Scholar] [CrossRef]
- Delnavazi, M.R.; Hadjiakhoondi, A.; Delazar, A.; Ajani, Y.; Tavakoli, S.; Yassa, N. Phytochemical and Antioxidant Investigation of the Aerial Parts of Dorema Glabrum Fisch. & C.A. Mey. Iran. J. Pharm. Res. 2015, 14, 925–931. [Google Scholar] [CrossRef]
- Petrović, G.M.; Stamenković, J.G.; Mitić, V.D.; Stojanović, G.S.; Zlatković, B.K.; Jovanović, O. Chemical Composition and Antioxidant Activity of the Athamanta turbith ssp. Haynaldii Volatiles. Nat. Prod. Commun. 2018, 13, 783–785. [Google Scholar] [CrossRef]
- Rahman, N.; Xin, T.B.; Kamilah, H.; Ariffin, F. Effects of Osmotic Dehydration Treatment on Volatile Compound (Myristicin) Content and Antioxidants Property of Nutmeg (Myristica fragrans) Pericarp. J. Food Sci. Technol. 2018, 55, 183–189. [Google Scholar] [CrossRef]
- Torbati, M.; Nazemiyeh, H.; Lotfipour, F.; Nemati, M.; Asnaashari, S.; Fathiazad, F. Chemical Composition and in Vitro Antioxidant and Antibacterial Activity of Heracleum transcaucasicum and Heracleum anisactis Roots Essential Oil. BioImpacts 2014, 4, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Ansory, H.M.; Sari, E.N.; Nilawati, A.; Handayani, S.; Aznam, N. Sunscreen and Antioxidant Potential of Myristicin in Nutmeg Essential Oils (Myristica fragrans). In Proceedings of the 2nd Bakti Tunas Husada-Health Science International Conference (BTH-HSIC 2019), Jawa Barat, Indonesia, 5–6 October 2019; Atlantis Press: Paris, France, 2020; Volume 26, pp. 138–142. [Google Scholar] [CrossRef]
- Wang, Y.T.; Zhu, L.; Zeng, D.; Long, W.; Zhu, S.M. Chemical Composition and Anti-Inflammatory Activities of Essential Oil from Trachydium roylei. J. Food Drug Anal. 2016, 24, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Sumiwi, S.A.; Sihombing, O.S.; Subarnas, A.; Abdassah, M.; Levita, J. A Study to Predict Anti-Inflammatory Activity of Eugenol, Myristicin, and Limonene of Cinnamomum Sintoc. Int. J. Pharm. Pharm. Sci. 2015, 7, 51–54. [Google Scholar]
- Lee, C.J.; Huang, C.W.; Chen, L.G.; Wang, C.C. (+)-Erythro-∆80-7S,8R-Dihydroxy-3,30,50-Trimethoxy-8-O-40-Neolignan, an Anti-Acne Component in Degreasing Myristica fragrans Houtt. Molecules 2020, 25, 4563. [Google Scholar] [CrossRef] [PubMed]
- Kuropakornpong, P.; Itharat, A.; Panthong, S.; Sireeratawong, S.; Ooraikul, B. In Vitro and in Vivo Anti-Inflammatory Activities of Benjakul: A Potential Medicinal Product from Thai Traditional Medicine. Evid. Based Complement. Altern. Med. 2020, 2020, 9760948. [Google Scholar] [CrossRef]
- Ai, S.; Lin, G.; Bai, Y.; Liu, X.; Piao, L. QSAR Classification-Based Virtual Screening Followed by Molecular Docking Identification of Potential COX-2 Inhibitors in a Natural Product Library. J. Comput. Biol. 2019, 26, 1296–1315. [Google Scholar] [CrossRef] [PubMed]
- Jahandar, F.; Asgarpanah, J.; Najafizadeh, P.; Mousavi, Z. Anti-Inflammatory Activity and Chemical Composition of Pycnocycla Bashagardiana Fruit’s Essential Oil in Animal Models. Iran. J. Basic Med. Sci. 2018, 21, 188–193. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, W. Anti-Inflammatory Effect of Myristicin on RAW 264.7 Macrophages Stimulated with Polyinosinic-Polycytidylic Acid. Molecules 2011, 16, 7132–7142. [Google Scholar] [CrossRef] [PubMed]
- Qiburi, Q.; Ganbold, T.; Bao, Q.; Da, M.; Aoqier, A.; Temuqile, T.; Baigude, H. Bioactive Components of Ethnomedicine Eerdun Wurile Regulate the Transcription of Pro-Inflammatory Cytokines in Microglia. J. Ethnopharmacol. 2020, 246, 112241. [Google Scholar] [CrossRef]
- Lim, H.J.; Woo, K.W.; Lee, K.R.; Lee, S.K.; Kim, H.P. Inhibition of Proinflammatory Cytokine Generation in Lung Inflammation by the Leaves of Perilla frutescens and Its Constituents. Biomol. Ther. 2014, 22, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Huang, B.; Wang, G. Chemical Composition, Antinociceptive and Anti-Inflammatory Properties of Essential Oil from the Roots of Illicium lanceolatum. Nat. Prod. Res. 2012, 26, 1712–1714. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Kushnarenko, S.V.; Özek, G.; Kirpotina, L.N.; Sinharoy, P.; Utegenova, G.A.; Abidkulova, K.T.; Özek, T.; Başer, K.H.C.; Kovrizhina, A.R.; et al. Modulation of Human Neutrophil Responses by the Essential Oils from Ferula Akitschkensis and Their Constituents. J. Agric. Food Chem. 2016, 64, 7156–7170. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Pitonzo, R.; Schillaci, D. Antimicrobial and Antiproliferative Activity of Athamanta sicula L. (Apiaceae). Pharmacogn. Mag. 2011, 7, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Rosa, A.; Marongiu, B.; Atzeri, A.; Dessì, M.A.; Falconieri, D.; Porcedda, S. Extraction and Separation of Volatile and Fixed Oils from Seeds of Myristica fragrans by Supercritical CO2: Chemical Composition and Cytotoxic Activity on Caco-2 Cancer Cells. J. Food Sci. 2012, 77, C448–C453. [Google Scholar] [CrossRef]
- Rengasamy, G.; Venkataraman, A.; Veeraraghavan, V.P.; Jainu, M. Cytotoxic and Apoptotic Potential of Myristica fragrans Houtt. (Mace) Extract on Human Oral Epidermal Carcinoma KB Cell Lines. Braz. J. Pharm. Sci. 2018, 54, 1–7. [Google Scholar] [CrossRef]
- Martins, C.; Doran, C.; Silva, I.C.; Miranda, C.; Rueff, J.; Rodrigues, A.S. Myristicin from Nutmeg Induces Apoptosis via the Mitochondrial Pathway and down Regulates Genes of the DNA Damage Response Pathways in Human Leukaemia K562 Cells. Chem. Biol. Interact. 2014, 218, 1–9. [Google Scholar] [CrossRef]
- Al-Jumaily, E.; Al-Massody, A. Cytogenetic Study of The Pure Myristicin from Nutmeg (Myristica fragrans) On Rhabdomyosarrcoma Cell Line (In Vitro). Dav Int. J. Sci. 2012, 1, 2277–5536. [Google Scholar]
- Martins, C.; Doran, C.; Laires, A.; Rueff, J.; Rodrigues, A.S. Genotoxic and Apoptotic Activities of the Food Flavourings Myristicin and Eugenol in AA8 and XRCC1 Deficient EM9 Cells. Food Chem. Toxicol. 2011, 49, 385–392. [Google Scholar] [CrossRef]
- Marabini, L.; Neglia, L.; Monguzzi, E.; Galli, C.L.; Marinovich, M. Assessment of Toxicity of Myristicin and 1’-Hydroxymyristicin in HepG2 Cell Line. J. Pharmacol. Toxicol. 2017, 12, 170–179. [Google Scholar] [CrossRef][Green Version]
- Rattarom, R.; Sakpakdeejaroen, I.; Hansakul, P.; Itharat, A. Cytotoxic Activity against Small Cell Lung Cancer Cell Line and Chromatographic Fingerprinting of Six Isolated Compounds from the Ethanolic Extract of Benjakul. J. Med. Assoc. Thail. 2014, 97, S70–S75. [Google Scholar]
- Fraternale, D.; Ricci, D.; Calcabrini, C.; Guescini, M.; Martinelli, C.; Sestili, P. Cytotoxic Activity of Essential Oils of Aerial Parts and Ripe Fruits of Echinophora spinosa (Apiaceae). Nat. Prod. Commun. 2013, 8, 1645–1649. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Cantrell, C.L.; Jeliazkova, E.A.; Astatkie, T.; Zheljazkov, V.D. Utilization of Nutmeg (Myristica fragrans Houtt.) Seed Hydrodistillation Time to Produce Essential Oil Fractions with Varied Compositions And Pharmacological Effects. Molecules 2020, 25, 565. [Google Scholar] [CrossRef] [PubMed]
- Maxia, A.; Falconieri, D.; Piras, A.; Porcedda, S.; Marongiu, B.; Frau, M.A.; Gonçalves, M.J.; Cabral, C.; Cavaleiro, C.; Salgueiro, L. Chemical Composition and Antifungal Activity of Essential Oils and Supercritical CO2 Extracts of Apium nodiflorum (L.) Lag. Mycopathologia 2012, 174, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Vokk, R.; Lõugas, T.; Mets, K.; Kravets, M. Dill (Anethum graveolens L.) and Parsley (Petroselinum crispum (Mill.) Fuss) from Estonia: Seasonal Differences in Essential Oil Composition. Agron. Res. 2011, 9, 515–520. [Google Scholar]
- Linde, G.A.; Gazim, Z.C.; Cardoso, B.K.; Jorge, L.F.; Tešević, V.; Glamoćlija, J.; Soković, M.; Colauto, N.B. Antifungal and Antibacterial Activities of Petroselinum crispum Essential Oil. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Valente, V.M.M.; Jham, G.N.; Jardim, C.M.; Dhingra, O.D.; Ghiviriga, I. Major Antifungals in Nutmeg Essential Oil against Aspergillus flavus and A. ochraceus. J. Food Res. 2014, 4, 51. [Google Scholar] [CrossRef]
- Alizadeh, A.; Abdollahzadeh, H. Essential Oil Constituents and Antimicrobial Activity of Pycnocycla bashagardiana Mozaff. from Iran. Nat. Prod. Res. 2017, 31, 2081–2084. [Google Scholar] [CrossRef] [PubMed]
- Rameshkumar, K.B.; Nandu, T.G.; Anu Aravind, A.P.; Mathew, S.P.; Shiburaj, S. Chemical Composition and FtsZ GTPase Inhibiting Activity of the Essential Oil of Piper sarmentosum from Andaman Islands, India. J. Essent. Oil Res. 2017, 29, 430–435. [Google Scholar] [CrossRef]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Development of Antibacterial Conjugates Using Sulfamethoxazole with Monocyclic Terpenes: A Systematic Medicinal Chemistry Based Computational Approach. Comput. Methods Programs Biomed. 2017, 140, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Kumar Singh, V.; Kumar Dwivedy, A.; Kumar Chaudhari, A.; Upadhyay, N.; Singh, A.; Krishna Saha, A.; Ray Chaudhury, S.; Prakash, B.; Dubey, N.K. Assessment of Chemically Characterised Myristica fragrans Essential Oil against Fungi Contaminating Stored Scented Rice and Its Mode of Action as Novel Aflatoxin Inhibitor. Nat. Prod. Res. 2020, 34, 1611–1615. [Google Scholar] [CrossRef]
- You, C.X.; Jiang, H.Y.; Zhang, W.J.; Guo, S.S.; Yang, K.; Lei, N.; Ma, P.; Geng, Z.F.; Du, S.S.; Athanassiou, C. Contact Toxicity and Repellency of the Main Components from the Essential Oil of Clausena anisum-olens against Two Stored Product Insects. J. Insect Sci. 2015, 15, 87. [Google Scholar] [CrossRef]
- Bae, I.K.; Kim, K.; Choi, S.D.; Chang, K.S.; Lee, H.S.; Lee, S.E. Mosquito Larvicidal Activities of Naturally Occurring Compounds Derived from Piper Species. Appl. Biol. Chem. 2017, 60, 113–117. [Google Scholar] [CrossRef]
- Sh Abou-Elnaga, Z. Insecticidal Bioactivity of Eco-Friendly Plant Origin Chemicals against Culex pipiens and Aedes aegypti (Diptera: Culicidae). J. Entomol. Zool. Stud. 2014, 2, 340–347. [Google Scholar]
- Cossolin, J.F.S.; Pereira, M.J.B.; Martínez, L.C.; Turchen, L.M.; Fiaz, M.; Bozdoğan, H.; Serrão, J.E. Cytotoxicity of Piper aduncum (Piperaceae) Essential Oil in Brown Stink Bug Euschistus heros (Heteroptera: Pentatomidae). Ecotoxicology 2019, 28, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Maggi, F.; Cianfaglione, K.; Canale, A.; Benelli, G. Promising Insecticidal Efficacy of the Essential Oils from the Halophyte echinophora spinosa (Apiaceae) Growing in Corsica Island, France. Environ. Sci. Pollut. Res. 2020, 27, 14454–14464. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R.; Iannarelli, R.; Petrelli, R.; Cappellacci, L.; Cianfaglione, K.; Afshar, F.H.; Nicoletti, M.; Canale, A.; Maggi, F. Synergized Mixtures of Apiaceae Essential Oils and Related Plant-Borne Compounds: Larvicidal Effectiveness on the Filariasis Vector Culex quinquefasciatus Say. Ind. Crop. Prod. 2017, 96, 186–195. [Google Scholar] [CrossRef]
- Kumar, R.; Krishan, S.R.I. Anti-Termite Activity of Essential Oil and Its Components from Myristica fragrans against Microcerotermes beesoni. J. Appl. Sci. Environ. Manag. 2011, 15, 559–561. [Google Scholar]
- Luo, C.; Li, D.L.; Wang, Y.; Guo, S.S.; Du, S.S. Bioactivities of 3-Butylidenephthalide and n-Butylbenzene from the Essential Oil of Ligusticum jeholense against Stored-Product Insects. J. Oleo Sci. 2019, 68, 931–937. [Google Scholar] [CrossRef]
- Heshmati Afshar, F.; Maggi, F.; Iannarelli, R.; Cianfaglione, K.; Isman, M.B. Comparative Toxicity of Helosciadium nodiflorum Essential Oils and Combinations of Their Main Constituents against the Cabbage Looper, Trichoplusia ni (Lepidoptera). Ind. Crop. Prod. 2017, 98, 46–52. [Google Scholar] [CrossRef]
- Qi, X.J.; Pang, X.; Cao, J.Q.; Du, S.S. Comparative Analysis on Bioactivity against Three Stored Insects of Ligusticum pteridophyllum Franch. Rhizomes Essential Oil and Supercritical Fluid (SFE-CO2) Extract. Environ. Sci. Pollut. Res. 2020, 27, 15584–15591. [Google Scholar] [CrossRef]
- Dorla, E.; Gauvin-Bialecki, A.; Deuscher, Z.; Allibert, A.; Grondin, I.; Deguine, J.P.; Laurent, P. Insecticidal Activity of the Leaf Essential Oil of Peperomia borbonensis miq. (Piperaceae) and Its Major Components against the Melon Fly Bactrocera cucurbitae (Diptera: Tephritidae). Chem. Biodivers. 2017, 14, e1600493. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.C.; Liu, Z.L. Analysis of the Essential Oil of Illicium Henryi Diels Root Bark and Its Insecticidal Activity against Liposcelis bostrychophila Badonnel. J. Food Prot. 2015, 78, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.M.; Jung, C.S.; Kang, J.; Lee, H.R.; Kim, S.W.; Hyun, J.; Park, I.K. Larvicidal and Acetylcholinesterase Inhibitory Activities of Apiaceae Plant Essential Oils and Their Constituents against Aedes albopictus and Formulation Development. J. Agric. Food Chem. 2015, 63, 9977–9986. [Google Scholar] [CrossRef]
- Pavela, R.; Maggi, F.; Cianfaglione, K.; Bruno, M.; Benelli, G. Larvicidal Activity of Essential Oils of Five Apiaceae taxa and Some of Their Main Constituents Against Culex quinquefasciatus. Chem. Biodivers. 2018, 15, e1700382. [Google Scholar] [CrossRef]
- Ajebli, M.; Eddouks, M. Antihypertensive Activity of Petroselinum crispum through Inhibition of Vascular Calcium Channels in Rats. J. Ethnopharmacol. 2019, 242, 112039. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, C.; Shen, X.; Xiao, L.; Wang, H.; Liu, P.; Wang, L.; Xu, H. Cytoprotective Effects of Myristicin against Hypoxia-Induced Apoptosis and Endoplasmic Reticulum Stress in Rat Dorsal Root Ganglion Neurons. Mol. Med. Rep. 2017, 15, 2280–2288. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, K.R.; Hong, J.H.; Lee, C.J. Effects of Caffeic Acid, Myristicin and Rosemarinic Acid on the Gene Expression and Production of Airway MUC5AC Mucin. Nat. Prod. Sci. 2016, 22, 275–281. [Google Scholar] [CrossRef]
- Estrada-Reyes, R.; Dorantes-Barrón, A.M.; Arrieta-Báez, D.; Gómez-Patiño, M.B.; Bernal-Trujillo, A.; Castro-García, M.; Carro-Juárez, M.; Martínez-Mota, L. Piper auritum Kunth (Piperaceae) Improves the Sexual Performance of Sluggish Male Rats through Enhancing Ejaculation. J. Ethnopharmacol. 2019, 231, 453–463. [Google Scholar] [CrossRef]
- Ghorbanian, D.; Ghasemi-Kasman, M.; Hashemian, M.; Gorji, E.; Gol, M.; Feizi, F.; Kazemi, S.; Ashrafpour, M.; Moghadamnia, A.A. Myristica fragrans Houtt Extract Attenuates Neuronal Loss and Glial Activation in Pentylenetetrazol-Induced Kindling Model. Iran. J. Pharm. Res. 2019, 18, 812–825. [Google Scholar] [CrossRef] [PubMed]
- Moinuddin, G.; Devi, K.; Kumar Khajuria, D. Evaluation of the Anti-Depressant Activity of Myristica fragrans (Nutmeg) in Male Rats. Avicenna J. Phytomed. 2012, 2, 72–78. [Google Scholar] [CrossRef]
- Ogawa, K.; Ito, M. Appetite-Enhancing Effects of Nutmeg Oil and Structure–Activity Relationship of Habituation to Phenylpropanoids. J. Nat. Med. 2019, 73, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Yuliana, N.D.; Iqbal, M.; Jahangir, M.; Wijaya, C.H.; Korthout, H.; Kottenhage, M.; Kim, H.K.; Verpoorte, R. Screening of Selected Asian Spices for Anti Obesity-Related Bioactivities. Food Chem. 2011, 126, 1724–1729. [Google Scholar] [CrossRef]
- Kobets, T.; Duan, J.D.; Brunnemann, K.D.; Etter, S.; Smith, B.; Williams, G.M. Structure-Activity Relationships for DNA Damage by Alkenylbenzenes in Turkey Egg Fetal Liver. Toxicol. Sci. Off. J. Soc. Toxicol. 2016, 150, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Hanif, F.; Singh, D.K. Binary Combination of Carica papaya, Areca catechu and Myristica fragrans with Piperonyl Butoxide/MGK-264 against Freshwater Snail Lymnaea acuminata. Trop. Life Sci. Res. 2013, 24, 1–11. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Biological Activity | Mechanisms | Species/Cell Lines |
|---|---|---|
| Antioxidant | Increases the concentration of catalase, superoxide dismutase, glutathione peroxidase glutathione reductase and decreases levels of lipid peroxidation | - |
| Anti-inflammatory | Inhibits PGE2, COX-2, tumor necrosis factor alpha (TNF-a), interleukins (IL-1, IL-6, IL-8, IL-10 and IL-17), nitric oxide (NO), macrophage inflammatory proteins (MIP-1α r MIP 1β), colony stimulating factor (GM-CSF), IP-10, MCP-1, MCP-3 and myeloperoxidase (MPO) | RAW 264.7, A549, HEK293, HL-60 and human fibroblast cells |
| Antiproliferative | Induces cell apoptosis through changes in mitochondrial membrane potential, cytochrome C release, caspase-3 activation, PARP cleavage, fragmentation of DNA, down-regulation of DNA damage response genes and reduces the expression of bcl-2 gene | K-562, NCI-H460, MCF-7, KB cell line, RD cells, Caco-2, AA8 and EM9, HepG2, NCI-H1688, MRC-5, U937 |
| Antimicrobial | Inhibition of the polymerization of FtsZ, of the enzyme dihydropteroate synthases (DHPSs) and of the GTPase enzyme | Fungi: Aspergillus flavus, Aspergillus fumigatus, Aspergillus niger, Aspergillus ochraceus, Aspergillus versicolor, Penicillium funiculosum, Penicillium ochrochloron, Penicillium verrucosum, Trichoderma viride Bacteria: Bacillus cereus, Enterobacter cloacae, Escherichia coli, Listeria monocytogenes, Pseudomonas aeruginosa, Salmonella enterica, Staphylococcus aureus, Cutibacterium acnes |
| Insecticide and larvicide | Inhibition of the CYP450 enzyme and acetylcholinesterase in insects | Liposcelis bostrychophila and Lasioderma serricorne, Culex pipiens (larva), Aedes aegypti, Euschistus heros, Culex quinquefasciatus (larva), Spodoptera littoralis (larva), Musca domestica (adult), and Spodoptera littoralis, Trichoplusia ni, Tribolium castaneum, Lasioderma serricorne, Liposcelis bostrychophila and Microcerotermes beesoni |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seneme, E.F.; dos Santos, D.C.; Silva, E.M.R.; Franco, Y.E.M.; Longato, G.B. Pharmacological and Therapeutic Potential of Myristicin: A Literature Review. Molecules 2021, 26, 5914. https://doi.org/10.3390/molecules26195914
Seneme EF, dos Santos DC, Silva EMR, Franco YEM, Longato GB. Pharmacological and Therapeutic Potential of Myristicin: A Literature Review. Molecules. 2021; 26(19):5914. https://doi.org/10.3390/molecules26195914
Chicago/Turabian StyleSeneme, Elisa Frederico, Daiane Carla dos Santos, Evelyn Marcela Rodrigues Silva, Yollanda Edwirges Moreira Franco, and Giovanna Barbarini Longato. 2021. "Pharmacological and Therapeutic Potential of Myristicin: A Literature Review" Molecules 26, no. 19: 5914. https://doi.org/10.3390/molecules26195914
APA StyleSeneme, E. F., dos Santos, D. C., Silva, E. M. R., Franco, Y. E. M., & Longato, G. B. (2021). Pharmacological and Therapeutic Potential of Myristicin: A Literature Review. Molecules, 26(19), 5914. https://doi.org/10.3390/molecules26195914