
Serum Proteomic Analysis of Cannabis Use Disorder in Male Patients


 ,
,  , , , , ,
, , , , ,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Demographic Information
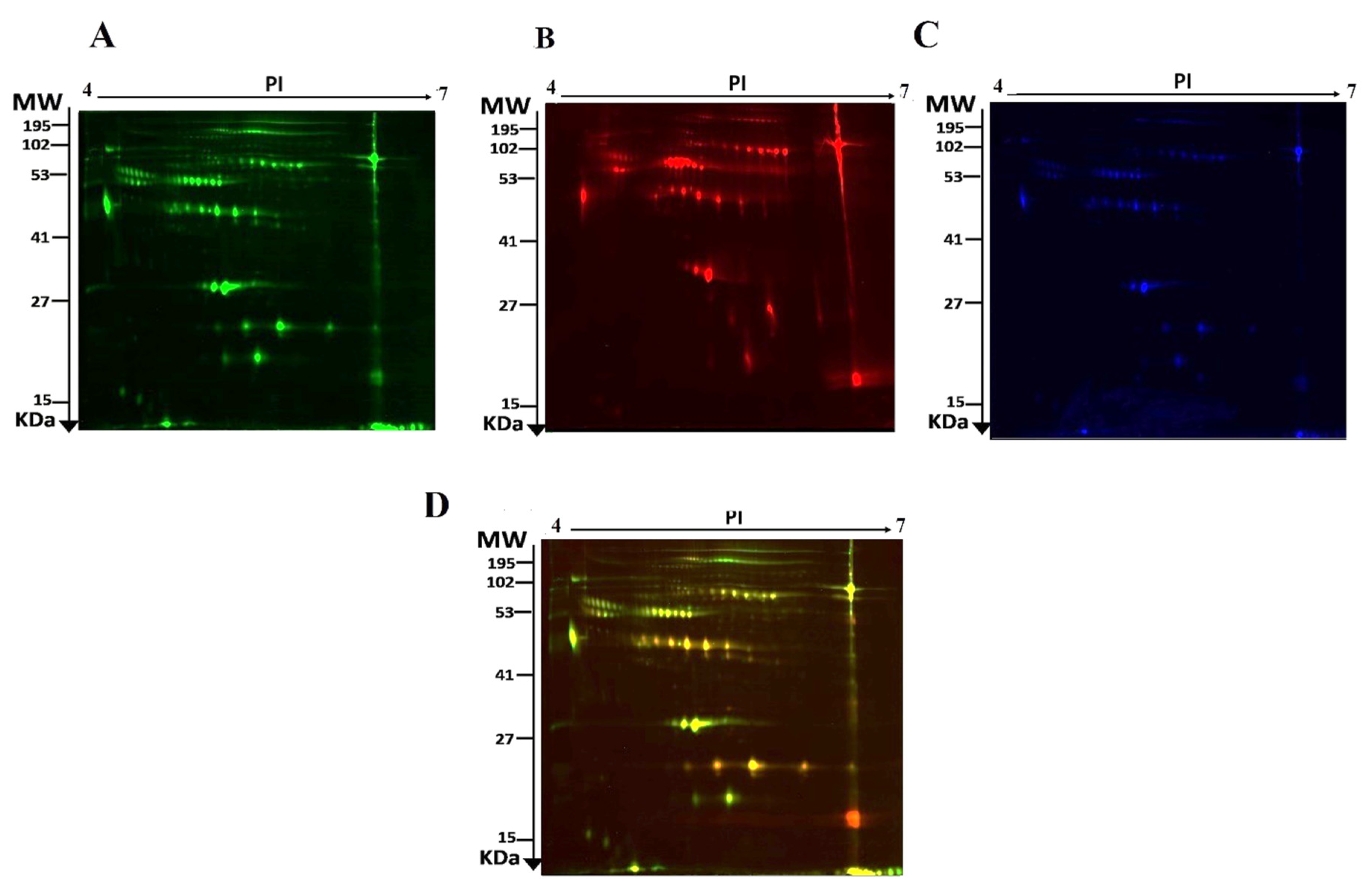
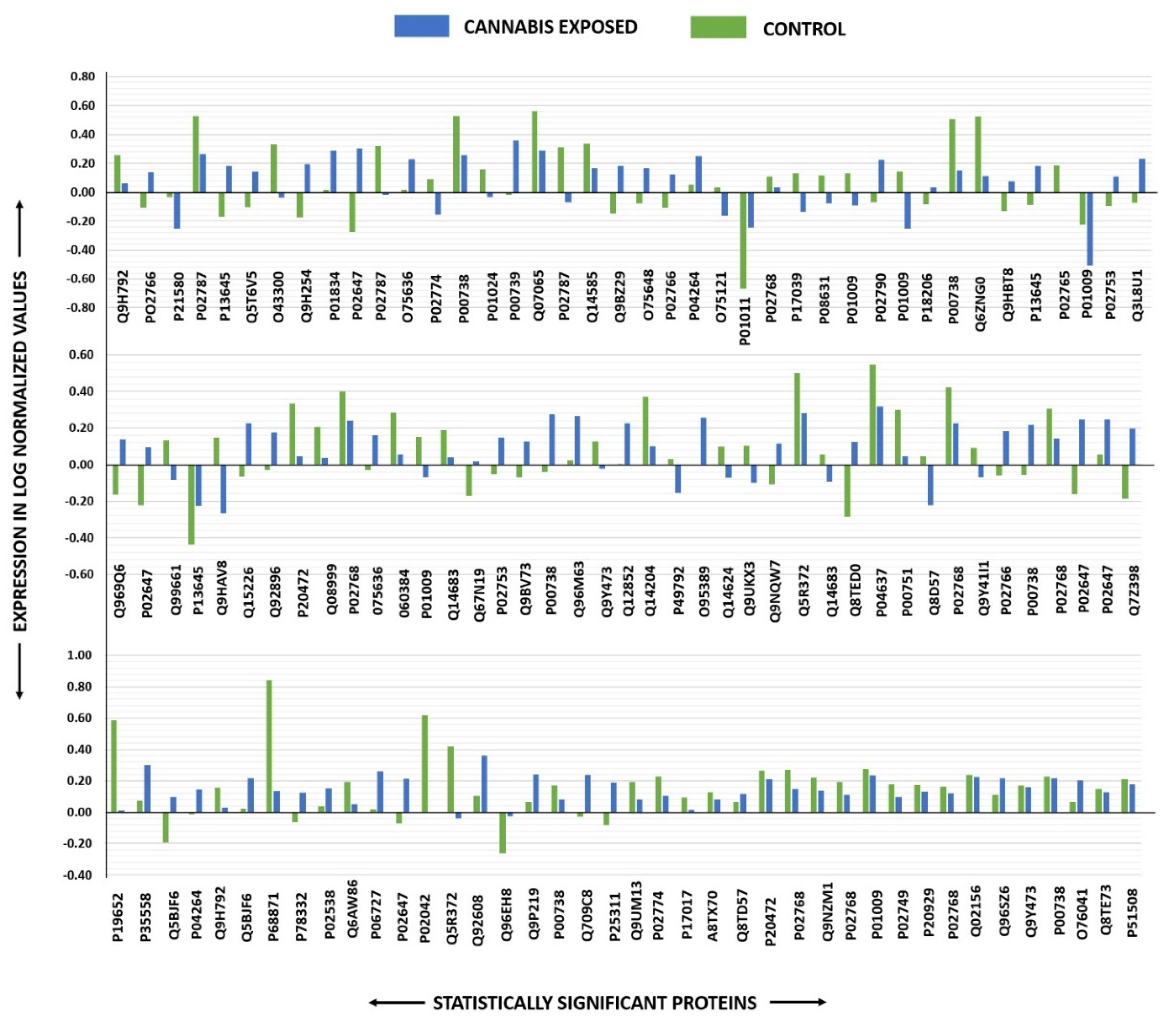
2.2. Identification of Differentially Expressed Proteins and 2D-DIGE Analysis
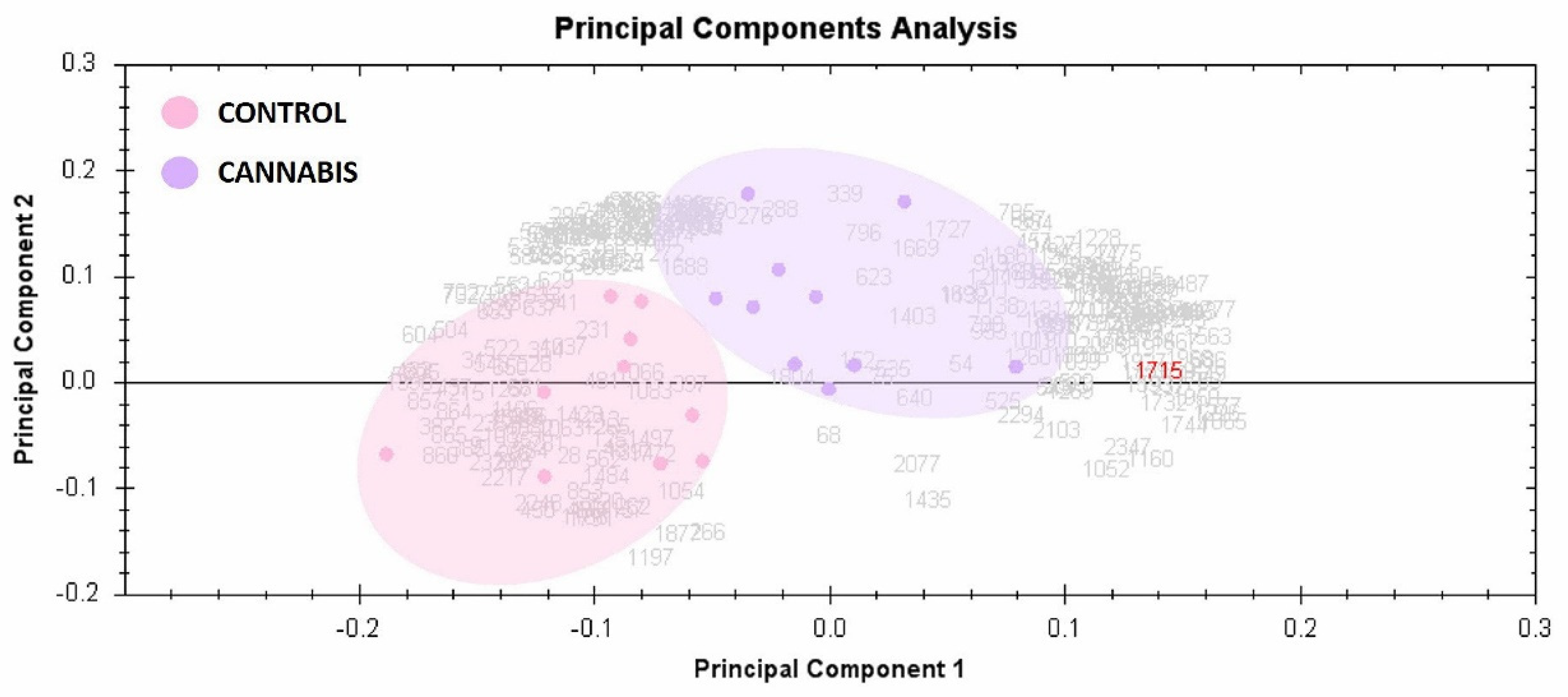
2.3. Principal Component Analysis
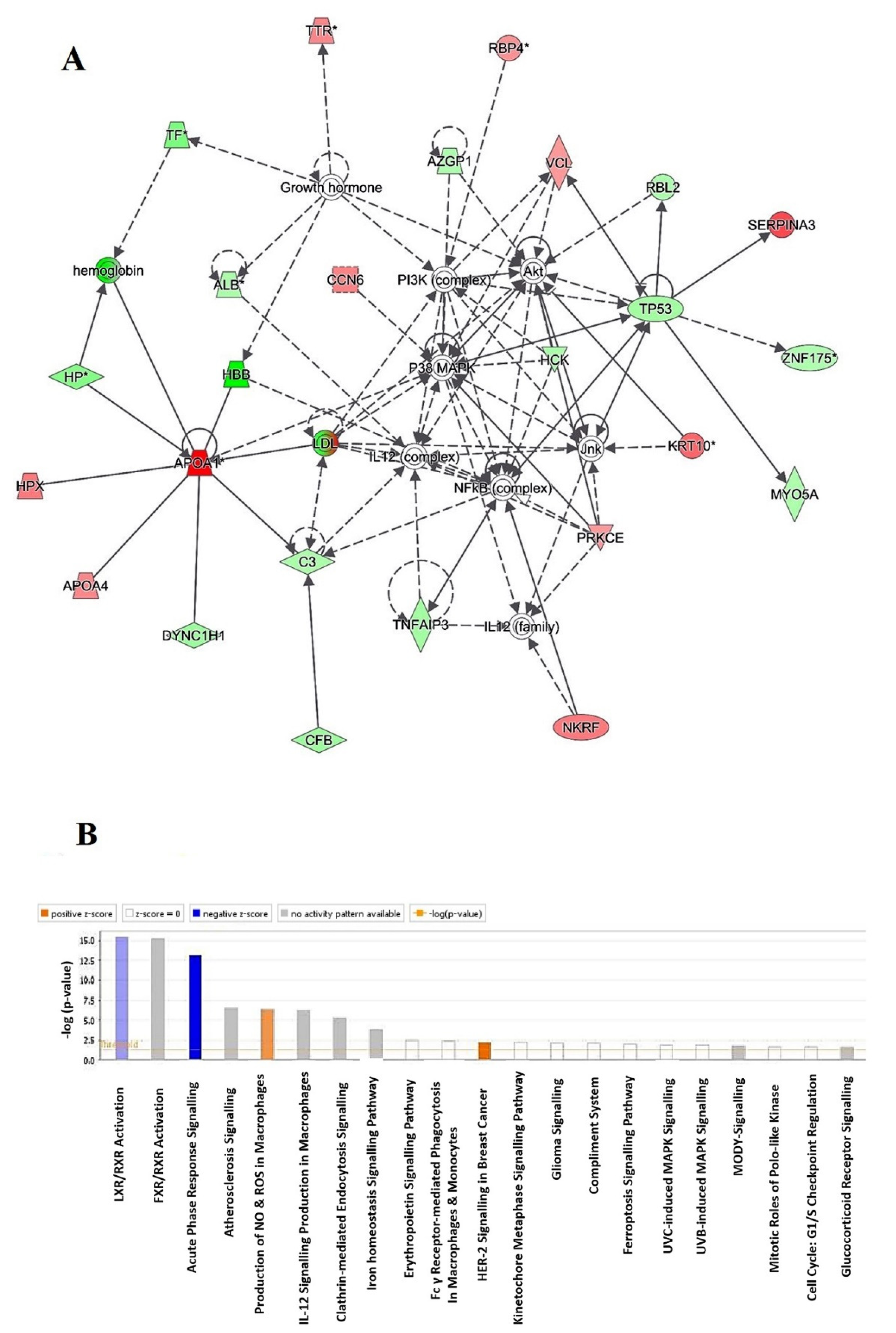
2.4. Protein–Protein Interaction Networks
2.5. Subcellular and Functional Characterization of the Differentially Expressed Proteins
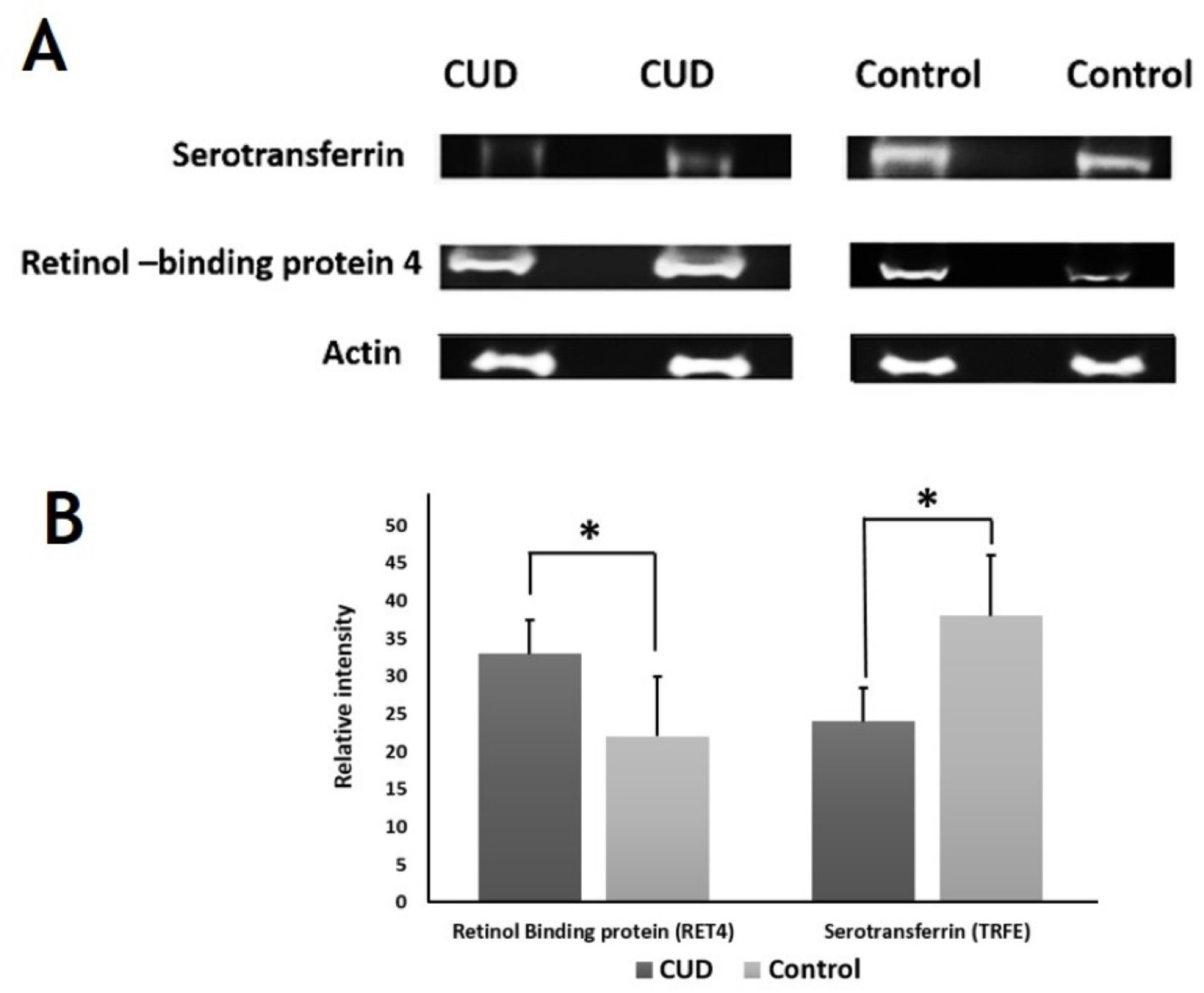
2.6. Immunoblotting Confirmation of Changes in Selected Proteins
3. Discussion
3.1. Ingenuity Pathway Analysis
3.1.1. LXR/RXR Activation
3.1.2. FXR/RXR Activation
3.1.3. Acute Phase Response Signaling
3.1.4. Atherosclerosis Signaling
3.1.5. Production of Nitric Oxide (NO) and Reactive Oxygen Species (ROS) in Macrophages
3.2. Selected Proteins
3.2.1. Albumin
3.2.2. Haptoglobin
3.2.3. Apolipoprotein A-I
3.2.4. Type I and Type II Keratins
3.2.5. Serotransferrin
3.2.6. Transthyretin
3.2.7. Tumor Necrosis Factor Alpha-Induced Protein 3
3.2.8. Inactive Tyrosine Protein Kinase PEAK1
4. Materials and Methods
4.1. Ethical Approval and Participate Consent
4.2. Study Design and Selection Criteria
4.3. Serum Protein Extraction
4.4. Fluorescence Labeling of Samples with CyDyes and 2-Dimensional Difference in Gel Electrophoresis (2D-DIGE)
4.5. Statistical Analysis
4.6. Protein Identification with Mass Spectrometry
4.7. Network Pathway and Functional Analysis
4.8. Immunoblotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Atakan, Z. Cannabis, a complex plant: Different compounds and different effects on individuals. Ther. Adv. Psychopharmacol. 2012, 2, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Casajuana Kögel, C.; López-Pelayo, H.; Balcells-Olivero, M.M.; Colom, J.; Gual, A. Psychoactive constituents of cannabis and their clinical implications: A systematic review. Adicciones 2018, 30, 140–151. [Google Scholar]
- Winton-Brown, T.T.; Allen, P.; Bhattacharrya, S.; Borgwardt, S.J.; Fusar-Poli, P.; Crippa, J.A.; Seal, M.L.; Martin-Santos, R.; Ffytche, D.; Zuardi, A.W. Modulation of auditory and visual processing by delta-9-tetrahydrocannabinol and cannabidiol: An FMRI study. Neuropsychopharmacology 2011, 36, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.A.; Zhou, R.; Jacobson, C.; Weng, J.; Cheng, E.; Lay, J.; Hung, P.; Lerner, J.T.; Sankar, R. Perceived efficacy of cannabidiol-enriched cannabis extracts for treatment of pediatric epilepsy: A potential role for infantile spasms and Lennox–Gastaut syndrome. Epilepsy Behav. 2015, 47, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Laux, L.C.; Bebin, E.M.; Checketts, D.; Chez, M.; Flamini, R.; Marsh, E.D.; Miller, I.; Nichol, K.; Park, Y.; Segal, E. Long-term safety and efficacy of cannabidiol in children and adults with treatment resistant Lennox-Gastaut syndrome or Dravet syndrome: Expanded access program results. Epilepsy Res. 2019, 154, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, N.E. Cannabinoids in the treatment of chemotherapy-induced nausea and vomiting: Beyond prevention of acute emesis. J. Support Oncol. 2007, 5, 1–9. [Google Scholar]
- Badowski, M.E.; Yanful, P.K. Dronabinol oral solution in the management of anorexia and weight loss in AIDS and cancer. Ther. Clin. Risk Manag. 2018, 14, 643. [Google Scholar] [CrossRef]
- Hasin, D.S.; Kerridge, B.T.; Saha, T.D.; Huang, B.; Pickering, R.; Smith, S.M.; Jung, J.; Zhang, H.; Grant, B.F. Prevalence and correlates of DSM-5 cannabis use disorder, 2012–2013: Findings from the National Epidemiologic Survey on Alcohol and Related Conditions–III. Am. J. Psychiatry 2016, 173, 588–599. [Google Scholar] [CrossRef]
- Henquet, C.; Rosa, A.; Delespaul, P.; Papiol, S.; Faňanás, L.; Van Os, J.; Myin-Germeys, I. COMT Val158Met moderation of cannabis-induced psychosis: A momentary assessment study of ‘switching on’hallucinations in the flow of daily life. Acta Psychiatr. Scand. 2009, 119, 156–160. [Google Scholar] [CrossRef]
- Lee, C.-R.; Lee, A.; Goodman, S.; Hammond, D.; Fischer, B. The Lower-Risk Cannabis Use Guidelines’(LRCUG) recommendations: How are Canadian cannabis users complying? Prev. Med. Rep. 2020, 20, 101187. [Google Scholar] [CrossRef]
- Fischer, B.; Russell, C.; Sabioni, P.; Van Den Brink, W.; Le Foll, B.; Hall, W.; Rehm, J.; Room, R. Lower-risk cannabis use guidelines: A comprehensive update of evidence and recommendations. Am. J. Public Health 2017, 107, e1–e12. [Google Scholar] [CrossRef]
- Ashton, C.H. Pharmacology and effects of cannabis: A brief review. Br. J. Psychiatry 2001, 178, 101–106. [Google Scholar] [CrossRef]
- Wu, T.-C.; Tashkin, D.P.; Djahed, B.; Rose, J.E. Pulmonary hazards of smoking marijuana as compared with tobacco. N. Engl. J. Med. 1988, 318, 347–351. [Google Scholar] [CrossRef]
- Malviya, A.; Khan, S.A.; Gupta, A.; Mishra, A. Chronic Marijuana Consumption Leading to High-Grade Atrioventricular Block in a Young Male. Cureus 2021, 13, e16202. [Google Scholar]
- Adapa, S.; Gayam, V.; Konala, V.M.; Annangi, S.; Raju, M.P.; Bezwada, V.; McMillan, C.; Dalal, H.; Mandal, A.; Naramala, S. Cannabis Vaping–Induced Acute Pulmonary Toxicity: Case Series and Review of Literature. J. Investig. Med. High Impact Case Rep. 2020, 8, 2324709620947267. [Google Scholar] [CrossRef]
- Noble, M.J.; Hedberg, K.; Hendrickson, R.G. Acute cannabis toxicity. Clin. Toxicol. 2019, 57, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.M.; Kaplan, B.L. Immune responses regulated by cannabidiol. Cannabis Cannabinoid Res. 2020, 5, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and anti-inflammatory properties of cannabidiol. Antioxidants 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Colleoni, M.; Conti, S.; Parolaro, D.; Franke, C.; Trovato, A.E.; Giagnoni, G. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 369, 294–299. [Google Scholar] [CrossRef]
- Izzo, A.A.; Capasso, R.; Aviello, G.; Borrelli, F.; Romano, B.; Piscitelli, F.; Gallo, L.; Capasso, F.; Orlando, P.; Di Marzo, V. Inhibitory effect of cannabichromene, a major non-psychotropic cannabinoid extracted from Cannabis sativa, on inflammation-induced hypermotility in mice. Br. J. Pharmacol. 2012, 166, 1444–1460. [Google Scholar] [CrossRef]
- Pottiez, G.; Jagadish, T.; Yu, F.; Letendre, S.; Ellis, R.; Duarte, N.A.; Grant, I.; Gendelman, H.E.; Fox, H.S.; Ciborowski, P. Plasma proteomic profiling in HIV-1 infected methamphetamine abusers. PLoS ONE 2012, 7, e31031. [Google Scholar] [CrossRef]
- Shi, W.-L.; Zhao, X.; Liu, Z.-M.; Zhang, M.; Zhou, B.-Y.; Pu, X.-P. Serum proteomics of methamphetamine addicts and up-regulation of complement factor H related to methamphetamine addiction. Neurosci. Lett. 2012, 525, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Alasmari, F.; Alsanea, S.; Masood, A.; Alhazzani, K.; Alanazi, I.O.; Musambil, M.; Alfadda, A.A.; Alshammari, M.A.; Alasmari, A.F.; Benabdelkamel, H. Serum proteomic profiling of patients with amphetamine use disorder. Drug Alcohol. Depend. 2020, 214, 108157. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.F.; Mikulskis, A.; Kuzdzal, S.; Bennett, D.A.; Kelly, J.; Golenko, E.; DiCesare, J.; Denoyer, E.; Patton, W.F.; Ediger, R. High-resolution serum proteomic profiling of Alzheimer disease samples reveals disease-specific, carrier-protein–bound mass signatures. Clin. Chem. 2005, 51, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Rong, C.; Ke, R.; Meng, S.; Yan, X.; Ke, H.; Wu, S. Differential proteomic analysis of serum exosomes reveals alterations in progression of Parkinson disease. Medicine 2019, 98, e17478. [Google Scholar] [CrossRef]
- Corbett, B.; Kantor, A.; Schulman, H.; Walker, W.; Lit, L.; Ashwood, P.; Rocke, D.M.; Sharp, F.R. A proteomic study of serum from children with autism showing differential expression of apolipoproteins and complement proteins. Mol. Psychiatry 2007, 12, 292–306. [Google Scholar] [CrossRef]
- Das, T.; Chen, Z.; Hendriks, R.W.; Kool, M. A20/Tumor necrosis factor α-induced protein 3 in immune cells controls development of autoinflammation and autoimmunity: Lessons from mouse models. Front. Immunol. 2018, 9, 104. [Google Scholar] [CrossRef]
- Taurines, R.; Dudley, E.; Conner, A.C.; Grassl, J.; Jans, T.; Guderian, F.; Mehler-Wex, C.; Warnke, A.; Gerlach, M.; Thome, J. Serum protein profiling and proteomics in autistic spectrum disorder using magnetic bead-assisted mass spectrometry. Eur. Arch. Psychiatry Clin. Neurosci. 2010, 260, 249–255. [Google Scholar] [CrossRef]
- Bot, M.; Chan, M.K.; Jansen, R.; Lamers, F.; Vogelzangs, N.; Steiner, J.; Leweke, F.M.; Rothermundt, M.; Cooper, J.; Bahn, S. Serum proteomic profiling of major depressive disorder. Transl. Psychiatry 2015, 5, e599. [Google Scholar] [CrossRef]
- Smirnova, L.; Seregin, A.; Boksha, I.; Dmitrieva, E.; Simutkin, G.; Kornetova, E.; Savushkina, O.; Letova, A.; Bokhan, N.; Ivanova, S. The difference in serum proteomes in schizophrenia and bipolar disorder. BMC Genom. 2019, 20, 1–14. [Google Scholar] [CrossRef]
- Juknat, A.; Pietr, M.; Kozela, E.; Rimmerman, N.; Levy, R.; Gao, F.; Coppola, G.; Geschwind, D.; Vogel, Z. Microarray and pathway analysis reveal distinct mechanisms underlying cannabinoid-mediated modulation of LPS-induced activation of BV-2 microglial cells. PLoS ONE 2013, 8, e61462. [Google Scholar] [CrossRef]
- Maqdasy, S.; Trousson, A.; Tauveron, I.; Volle, D.H.; Baron, S.; Lobaccaro, J.-M.A. Once and for all, LXRα and LXRβ are gatekeepers of the endocrine system. Mol. Asp. Med. 2016, 49, 31–46. [Google Scholar] [CrossRef]
- Zelcer, N.; Tontonoz, P. Liver X receptors as integrators of metabolic and inflammatory signaling. J. Clin. Investig. 2006, 116, 607–614. [Google Scholar] [CrossRef]
- Mukwaya, A.; Lennikov, A.; Xeroudaki, M.; Mirabelli, P.; Lachota, M.; Jensen, L.; Peebo, B.; Lagali, N. Time-dependent LXR/RXR pathway modulation characterizes capillary remodeling in inflammatory corneal neovascularization. Angiogenesis 2018, 21, 395–413. [Google Scholar] [CrossRef]
- Korf, H.; Vander Beken, S.; Romano, M.; Steffensen, K.R.; Stijlemans, B.; Gustafsson, J.-Å.; Grooten, J.; Huygen, K. Liver X receptors contribute to the protective immune response against Mycobacterium tuberculosis in mice. J. Clin. Investig. 2009, 119, 1626–1637. [Google Scholar] [CrossRef]
- Claudel, T.; Staels, B.; Kuipers, F. The Farnesoid X receptor: A molecular link between bile acid and lipid and glucose metabolism. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2020–2030. [Google Scholar] [CrossRef]
- Zhang, H.M.; Wang, X.; Wu, Z.H.; Liu, H.L.; Chen, W.; Zhang, Z.Z.; Chen, D.; Zeng, T.S. Beneficial effect of farnesoid X receptor activation on metabolism in a diabetic rat model. Mol. Med. Rep. 2016, 13, 2135–2142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lee, F.Y.; Barrera, G.; Lee, H.; Vales, C.; Gonzalez, F.J.; Willson, T.M.; Edwards, P.A. Activation of the nuclear receptor FXR improves hyperglycemia and hyperlipidemia in diabetic mice. Proc. Natl. Acad. Sci. USA 2006, 103, 1006–1011. [Google Scholar] [CrossRef]
- Nishimoto, N.; Yoshizaki, K.; Tagoh, H.; Monden, M.; Kishimoto, S.; Hirano, T.; Kishimoto, T. Elevation of serum interleukin 6 prior to acute phase proteins on the inflammation by surgical operation. Clin. Immunol. Immunopathol. 1989, 50, 399–401. [Google Scholar] [CrossRef]
- Pini, M.; Rhodes, D.H.; Fantuzzi, G. Hematological and acute-phase responses to diet-induced obesity in IL-6 KO mice. Cytokine 2011, 56, 708–716. [Google Scholar] [CrossRef]
- Sander, L.E.; Sackett, S.D.; Dierssen, U.; Beraza, N.; Linke, R.P.; Müller, M.; Blander, J.M.; Tacke, F.; Trautwein, C. Hepatic acute-phase proteins control innate immune responses during infection by promoting myeloid-derived suppressor cell function. J. Exp. Med. 2010, 207, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.S.; Yamamoto, Y.; Newton, C.; Friedman, H.; Klein, T. Psychoactive cannabinoids increase mortality and alter acute phase cytokine responses in mice sublethally infected with Legionella pneumophila. Proc. Soc. Exp. Biol. Med. 1997, 214, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Laprairie, R.; Bagher, A.; Kelly, M.; Denovan-Wright, E. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef]
- Van Stijn, C.M.; Kim, J.; Barish, G.D.; Tietge, U.J.; Tangirala, R.K. Adiponectin expression protects against angiotensin II-mediated inflammation and accelerated atherosclerosis. PLoS ONE 2014, 9, e86404. [Google Scholar] [CrossRef]
- Mach, F.; Montecucco, F.; Steffens, S. Cannabinoid receptors in acute and chronic complications of atherosclerosis. Br. J. Pharmacol. 2008, 153, 290–298. [Google Scholar] [CrossRef]
- Lépicier, P.; Bouchard, J.F.; Lagneux, C.; Lamontagne, D. Endocannabinoids protect the rat isolated heart against ischaemia. Br. J. Pharmacol. 2003, 139, 805–815. [Google Scholar] [CrossRef]
- Steffens, S.; Veillard, N.R.; Arnaud, C.; Pelli, G.; Burger, F.; Staub, C.; Zimmer, A.; Frossard, J.-L.; Mach, F. Low dose oral cannabinoid therapy reduces progression of atherosclerosis in mice. Nature 2005, 434, 782–786. [Google Scholar] [CrossRef]
- Chetty, K.; Lavoie, A.; Dehghani, P. A literature review of cannabis and myocardial infarction–what clinicians may not be aware of. CJC Open 2020, 3, 12–21. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, H.; Wei, J.; Sartor, G.C.; Bao, M.M.; Pierce, C.T.; Wahlestedt, C.R.; Dykxhoorn, D.M.; Dong, C. Cocaine exposure increases blood pressure and aortic stiffness via the miR-30c-5p–malic enzyme 1–reactive oxygen species pathway. Hypertension 2018, 71, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; Kuzontkoski, P.M.; Groopman, J.E.; Prasad, A. Cannabidiol induces programmed cell death in breast cancer cells by coordinating the cross-talk between apoptosis and autophagy. Mol. Cancer Ther. 2011, 10, 1161–1172. [Google Scholar] [CrossRef]
- Han, K.H.; Lim, S.; Ryu, J.; Lee, C.-W.; Kim, Y.; Kang, J.-H.; Kang, S.-S.; Ahn, Y.K.; Park, C.-S.; Kim, J.J. CB1 and CB2 cannabinoid receptors differentially regulate the production of reactive oxygen species by macrophages. Cardiovasc. Res. 2009, 84, 378–386. [Google Scholar] [CrossRef]
- Hampson, A.; Grimaldi, M.; Axelrod, J.; Wink, D. Cannabidiol and (−) Δ9-tetrahydrocannabinol are neuroprotective antioxidants. Proc. Natl. Acad. Sci. USA 1998, 95, 8268–8273. [Google Scholar] [CrossRef] [PubMed]
- Mbvundula, E.C.; Bunning, R.A.; Rainsford, K. Effects of cannabinoids on nitric oxide production by chondrocytes and proteoglycan degradation in cartilage. Biochem. Pharmacol. 2005, 69, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.A.; Brockie, H.C.; Pertwee, R.G. Inhibition of nitric oxide production in RAW264. 7 macrophages by cannabinoids and palmitoylethanolamide. Eur. J. Pharmacol. 2000, 401, 121–130. [Google Scholar] [CrossRef]
- Romano, B.; Borrelli, F.; Fasolino, I.; Capasso, R.; Piscitelli, F.; Cascio, M.; Pertwee, R.; Coppola, D.; Vassallo, L.; Orlando, P. The cannabinoid TRPA1 agonist cannabichromene inhibits nitric oxide production in macrophages and ameliorates murine colitis. Br. J. Pharmacol. 2013, 169, 213–229. [Google Scholar] [CrossRef]
- Coffey, R.G.; Yamamoto, Y.; Snella, E.; Pross, S. Tetrahydrocannabinol inhibition of macrophage nitric oxide production. Biochem. Pharmacol. 1996, 52, 743–751. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Yang, K.-H.; Pulaski, J.T.; Kaminski, N.E. Attenuation of inducible nitric oxide synthase gene expression by delta 9-tetrahydrocannabinol is mediated through the inhibition of nuclear factor-kappa B/Rel activation. Mol. Pharmacol. 1996, 50, 334–341. [Google Scholar]
- Bohnert, T.; Gan, L.-S. Plasma protein binding: From discovery to development. J. Pharm. Sci. 2013, 102, 2953–2994. [Google Scholar] [CrossRef]
- Eichel, E.A.; Ellingrod, V.L. Protein binding changes and drug interactions: What do we know. Curr. Psychiatry 2018, 17, 38–41. [Google Scholar]
- Vázquez, M.; Guevara, N.; Maldonado, C.; Guido, P.C.; Schaiquevich, P. Potential Pharmacokinetic Drug-Drug Interactions between Cannabinoids and Drugs Used for Chronic Pain. BioMed Res. Int. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Brown, J.D. Potential adverse drug events with tetrahydrocannabinol (THC) due to drug–drug interactions. J. Clin. Med. 2020, 9, 919. [Google Scholar] [CrossRef]
- Wang, Y.; Kinzie, E.; Berger, F.G.; Lim, S.-K.; Baumann, H. Haptoglobin, an inflammation-inducible plasma protein. Redox Rep. 2001, 6, 379–385. [Google Scholar] [CrossRef]
- Sharpe-Timms, K.; Nabli, H.; Zimmer, R.; Birt, J.; Davis, J. Inflammatory cytokines differentially up-regulate human endometrial haptoglobin production in women with endometriosis. Hum. Reprod. 2010, 25, 1241–1250. [Google Scholar] [CrossRef]
- Formukong, E.; Evans, A.; Evans, F. Analgesic and antiinflammatory activity of constituents of Cannabis sativa L. Inflammation 1988, 12, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Sumariwalla, P.F.; Feldmann, M.; Gallily, R. Cannabinoids in models of chronic inflammatory conditions. Phytochem. Rev. 2005, 4, 11–18. [Google Scholar] [CrossRef]
- Schaer, C.; Deuel, J.; Bittermann, A.; Rubio, I.; Schoedon, G.; Spahn, D.; Wepf, R.; Vallelian, F.; Schaer, D. Mechanisms of haptoglobin protection against hemoglobin peroxidation triggered endothelial damage. Cell Death Differ. 2013, 20, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Jelena, A.; Mirjana, M.; Desanka, B.; Svetlana, I.-M.; Aleksandra, U.; Goran, P.; Ilijana, G. Haptoglobin and the inflammatory and oxidative status in experimental diabetic rats: Antioxidant role of haptoglobin. J. Physiol. Biochem. 2013, 69, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Wolff, V.; Schlagowski, A.-I.; Rouyer, O.; Charles, A.-L.; Singh, F.; Auger, C.; Schini-Kerth, V.; Marescaux, C.; Raul, J.-S.; Zoll, J. Tetrahydrocannabinol induces brain mitochondrial respiratory chain dysfunction and increases oxidative stress: A potential mechanism involved in cannabis-related stroke. BioMed Res. Int. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Raja, A.; Ahmadi, S.; de Costa, F.; Li, N.; Kerman, K. Attenuation of Oxidative Stress by Cannabinoids and Cannabis Extracts in Differentiated Neuronal Cells. Pharmaceuticals 2020, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Valvassori, S.S.; Elias, G.; de Souza, B.; Petronilho, F.; Dal-Pizzol, F.; Kapczinski, F.; Trzesniak, C.; Tumas, V.; Dursun, S.; Nisihara Chagas, M.H. Effects of cannabidiol on amphetamine-induced oxidative stress generation in an animal model of mania. J. Psychopharmacol. 2011, 25, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Alagbonsi, I.A.; Olayaki, L.A. Role of oxidative stress in Cannabis sativa-associated spermatotoxicity: Evidence for ameliorative effect of combined but not separate melatonin and vitamin C. Middle East Fertil. Soc. J. 2017, 22, 136–144. [Google Scholar] [CrossRef]
- Nguyen, D.; Nickel, M.; Mizuguchi, C.; Saito, H.; Lund-Katz, S.; Phillips, M.C. Interactions of apolipoprotein AI with high-density lipoprotein particles. Biochemistry 2013, 52, 1963–1972. [Google Scholar] [CrossRef]
- Wacker, B.K.; Dronadula, N.; Bi, L.; Stamatikos, A.; Dichek, D.A. Apo AI (Apolipoprotein AI) vascular gene therapy provides durable protection against atherosclerosis in hyperlipidemic rabbits. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.M.; Jones, J.M.; Hall, A.G.; Tabner, S.A.; Kmiec, R.L. Theoretical explanation for reduced body mass index and obesity rates in cannabis users. Cannabis Cannabinoid Res. 2018, 3, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Verty, A.N.; Evetts, M.J.; Crouch, G.J.; McGregor, I.S.; Stefanidis, A.; Oldfield, B.J. The cannabinoid receptor agonist THC attenuates weight loss in a rodent model of activity-based anorexia. Neuropsychopharmacology 2011, 36, 1349–1358. [Google Scholar] [CrossRef]
- Mendizábal, V.E.; Adler-Graschinsky, E. Cannabinoids as therapeutic agents in cardiovascular disease: A tale of passions and illusions. Br. J. Pharmacol. 2007, 151, 427–440. [Google Scholar] [CrossRef]
- Robert, J.; Button, E.B.; Yuen, B.; Gilmour, M.; Kang, K.; Bahrabadi, A.; Stukas, S.; Zhao, W.; Kulic, I.; Wellington, C.L. Clearance of beta-amyloid is facilitated by apolipoprotein E and circulating high-density lipoproteins in bioengineered human vessels. Elife 2017, 6, e29595. [Google Scholar] [CrossRef]
- Button, E.B.; Boyce, G.K.; Wilkinson, A.; Stukas, S.; Hayat, A.; Fan, J.; Wadsworth, B.J.; Robert, J.; Martens, K.M.; Wellington, C.L. ApoA-I deficiency increases cortical amyloid deposition, cerebral amyloid angiopathy, cortical and hippocampal astrogliosis, and amyloid-associated astrocyte reactivity in APP/PS1 mice. Alzheimer’s Res. Ther. 2019, 11, 1–18. [Google Scholar] [CrossRef]
- Kim, S.H.; Yang, J.W.; Kim, K.H.; Kim, J.U.; Yook, T.H. A Review on Studies of Marijuana for Alzheimer’s Disease–Focusing on CBD, THC. J. Pharmacopunct. 2019, 22, 225. [Google Scholar]
- Boiko, A.S.; Mednova, I.A.; Kornetova, E.G.; Semke, A.V.; Bokhan, N.A.; Loonen, A.J.; Ivanova, S.A. Apolipoprotein serum levels related to metabolic syndrome in patients with schizophrenia. Heliyon 2019, 5, e02033. [Google Scholar] [CrossRef] [PubMed]
- Schoevers, J.; Leweke, J.E.; Leweke, F.M. Cannabidiol as a treatment option for schizophrenia: Recent evidence and current studies. Curr. Opin. Psychiatry 2020, 33, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Crippa, J.A.; Allen, P.; Martin-Santos, R.; Borgwardt, S.; Fusar-Poli, P.; Rubia, K.; Kambeitz, J.; O’Carroll, C.; Seal, M.L. Induction of psychosis byδ9-tetrahydrocannabinol reflects modulation of prefrontal and striatal function during attentional salience processing. Arch. Gen. Psychiatry 2012, 69, 27–36. [Google Scholar] [CrossRef]
- Cortes-Briones, J.A.; Cahill, J.D.; Skosnik, P.D.; Mathalon, D.H.; Williams, A.; Sewell, R.A.; Roach, B.J.; Ford, J.M.; Ranganathan, M.; D’Souza, D.C. The psychosis-like effects of Δ9-tetrahydrocannabinol are associated with increased cortical noise in healthy humans. Biol. Psychiatry 2015, 78, 805–813. [Google Scholar] [CrossRef]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef] [PubMed]
- Dmello, C.; Srivastava, S.S.; Tiwari, R.; Chaudhari, P.R.; Sawant, S.; Vaidya, M.M. Multifaceted role of keratins in epithelial cell differentiation and transformation. J. Biosci. 2019, 44, 1–16. [Google Scholar] [CrossRef]
- Pan, X.; Hobbs, R.P.; Coulombe, P.A. The expanding significance of keratin intermediate filaments in normal and diseased epithelia. Curr. Opin. Cell Biol. 2013, 25, 47–56. [Google Scholar] [CrossRef]
- Alam, H.; Sehgal, L.; Kundu, S.T.; Dalal, S.N.; Vaidya, M.M. Novel function of keratins 5 and 14 in proliferation and differentiation of stratified epithelial cells. Mol. Biol. Cell 2011, 22, 4068–4078. [Google Scholar] [CrossRef] [PubMed]
- Ramms, L.; Fabris, G.; Windoffer, R.; Schwarz, N.; Springer, R.; Zhou, C.; Lazar, J.; Stiefel, S.; Hersch, N.; Schnakenberg, U. Keratins as the main component for the mechanical integrity of keratinocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 18513–18518. [Google Scholar] [CrossRef]
- Gu, L.-H.; Coulombe, P.A. Keratin function in skin epithelia: A broadening palette with surprising shades. Curr. Opin. Cell Biol. 2007, 19, 13–23. [Google Scholar] [CrossRef]
- Yi, H.; Yoon, H.-N.; Kim, S.; Ku, N.-O. The role of keratins in the digestive system: Lessons from transgenic mouse models. Histochem. Cell Biol. 2018, 150, 351–359. [Google Scholar] [CrossRef]
- Tolosano, E. Increasing serum transferrin to reduce tissue iron overload due to ineffective erythropoiesis. Haematologica 2015, 100, 565. [Google Scholar] [CrossRef] [PubMed]
- Nedumaran, B.; Rudra, P.; Gaydos, J.; Kumar, S.; Meacham, R.B.; Burnham, E.L.; Malykhina, A.P. Impact of regular Cannabis use on biomarkers of lower urinary tract function. Urology 2017, 109, 223.e9–223.e16. [Google Scholar] [CrossRef]
- Kelsen, S.G.; Duan, X.; Ji, R.; Perez, O.; Liu, C.; Merali, S. Cigarette smoke induces an unfolded protein response in the human lung: A proteomic approach. Am. J. Respir. Cell Mol. Biol. 2008, 38, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.J.; Wijayagunaratne, R.C.; D’Souza, D.G.; Darras, V.M.; Van Herck, S.L. Transport of thyroid hormones via the choroid plexus into the brain: The roles of transthyretin and thyroid hormone transmembrane transporters. Front. Neurosci. 2015, 9, 66. [Google Scholar] [CrossRef]
- Ghadami, S.A.; Chia, S.; Ruggeri, F.S.; Meisl, G.; Bemporad, F.; Habchi, J.; Cascella, R.; Dobson, C.M.; Vendruscolo, M.; Knowles, T.P. Transthyretin inhibits primary and secondary nucleations of amyloid-β peptide aggregation and reduces the toxicity of its oligomers. Biomacromolecules 2020, 21, 1112–1125. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Ladiwala, A.R.A.; Du, D.; Yadav, J.K.; Tessier, P.M.; Wright, P.E.; Kelly, J.W.; Buxbaum, J.N. Mechanisms of transthyretin inhibition of β-amyloid aggregation in vitro. J. Neurosci. 2013, 33, 19423–19433. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D. NF-κB signaling in macrophages: Dynamics, crosstalk, and signal integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef] [PubMed]
- Vereecke, L.; Beyaert, R.; van Loo, G. Genetic relationships between A20/TNFAIP3, chronic inflammation and autoimmune disease. Biochem. Soc. Trans. 2011, 39, 1086–1091. [Google Scholar] [CrossRef]
- Majumdar, I.; Ahuja, V.; Paul, J. Altered expression of tumor necrosis factor alpha-induced protein 3 correlates with disease severity in ulcerative colitis. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, Z.; Yuan, J.; Li, L. Altered TNFAIP3 mRNA expression in peripheral blood mononuclear cells from patients with rheumatoid arthritis. Biomed. Rep. 2015, 3, 675–680. [Google Scholar] [CrossRef][Green Version]
- Jiang, X.; Tian, H.; Fan, Y.; Chen, J.; Song, Y.; Wang, S.; Zhu, F.; Guo, C.; Zhang, L.; Shi, Y. Expression of tumor necrosis factor alpha-induced protein 3 mRNA in peripheral blood mononuclear cells negatively correlates with disease severity in psoriasis vulgaris. Clin. Vaccine Immunol. 2012, 19, 1938–1942. [Google Scholar] [CrossRef]
- Hadisaputri, Y.E.; Miyazaki, T.; Yokobori, T.; Sohda, M.; Sakai, M.; Ozawa, D.; Hara, K.; Honjo, H.; Kumakura, Y.; Kuwano, H. TNFAIP3 overexpression is an independent factor for poor survival in esophageal squamous cell carcinoma. Int. J. Oncol. 2017, 50, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Agajanian, M.; Campeau, A.; Hoover, M.; Hou, A.; Brambilla, D.; Kim, S.L.; Klemke, R.L.; Kelber, J.A. PEAK1 acts as a molecular switch to regulate context-dependent TGFβ responses in breast cancer. PLoS ONE 2015, 10, e0135748. [Google Scholar] [CrossRef]
- Croucher, D.R.; Hochgräfe, F.; Zhang, L.; Liu, L.; Lyons, R.J.; Rickwood, D.; Tactacan, C.M.; Browne, B.C.; Ali, N.; Chan, H. Involvement of Lyn and the atypical kinase SgK269/PEAK1 in a basal breast cancer signaling pathway. Cancer Res. 2013, 73, 1969–1980. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Tang, W.; Fan, X.; Wang, X.; Wu, H.; Xu, H.; Xu, W.; Gao, W.; Wu, G. Overexpression of PEAK1 contributes to epithelial–mesenchymal transition and tumor metastasis in lung cancer through modulating ERK1/2 and JAK2 signaling. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Pub.: Washington, DC, USA, 2013. [Google Scholar]
- Chen, Y.Y.; Lin, S.Y.; Yeh, Y.Y.; Hsiao, H.H.; Wu, C.Y.; Chen, S.T.; Wang, A.H.J. A modified protein precipitation procedure for efficient removal of albumin from serum. Electrophoresis 2005, 26, 2117–2127. [Google Scholar] [CrossRef] [PubMed]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Jammah, A.A.; Ekhzaimy, A.A. Differences in the plasma proteome of patients with hypothyroidism before and after thyroid hormone replacement: A proteomic analysis. Int. J. Mol. Sci. 2018, 19, 88. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Masood, A.; Benabdelkamel, H.; Ekhzaimy, A.A.; Alfadda, A.A. Plasma-Based Proteomics Profiling of Patients with Hyperthyroidism after Antithyroid Treatment. Molecules 2020, 12, 2831. [Google Scholar] [CrossRef]
- Benabdelkamel, H.; Masood, A.; Almidani, G.M.; Alsadhan, A.A.; Bassas, A.F.; Duncan, M.W.; Alfadda, A.A. Mature adipocyte proteome reveals differentially altered protein abundances between lean, Overweight and morbidly obese human subjects. Mol. Cell Endocrinol. 2015, 401, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Moustafa, A.; Sallam, R.; Bassas, A.; Duncan, M. Proteomic analysis of mature adipocytes from obese patients in relation to aging. Exp. Gerontol. 2013, 48, 1196–1203. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CUD Group | Control Group | |
|---|---|---|
| Number of patients | 10 | 10 |
| Gender | 10 Male, 0 Female | 10 Male, 0 Female |
| Age in years (Mean ± SD) | 24.7 ± 3.63 | 30.4 ± 4.36 |
| Infectious diseases (HIV, HCV, TB) | Negative | Negative |
| Cannabis dosage form | Smoking | None |
| Cannabis use history | ||
| 2–5 years | 4 Patients | None |
| 6–10 years | 3 Patients | None |
| ≥11 years | 3 Patients | None |
| Sl No: | Spot No a | Accession No | Protein Name | Mascot ID | p-Value b (ANOVA) | Ratio CB/C c | Exp d |
|---|---|---|---|---|---|---|---|
| Upregulated Proteins | |||||||
| 84 | 990 | Q5BJF6 | Outer dense fiber protein 2 | ODFP2_HUMAN | 0.05 | 2 | UP |
| 10 | 1470 | P02647 | Apolipoprotein A-I | APOA1_HUMAN | 0.01 | 3.8 | UP |
| 25 | 185 | P01011 | Alpha-1-antichymotrypsin | AACT_HUMAN | 0.02 | 2.7 | UP |
| 79 | 1627 | P02647 | Apolipoprotein A-I | APOA1_HUMAN | 0.04 | 2.6 | UP |
| 70 | 1180 | Q8TED0 | U3 small nucleolar RNA-associated protein 15 homolog | UTP15_HUMAN | 0.03 | 2.6 | UP |
| 16 | 1322 | P00739 | Haptoglobin-related protein | HPTR_HUMAN | 0.01 | 2.4 | UP |
| 81 | 993 | Q7Z398 | Zinc finger protein 550 | ZN550_HUMAN | 0.04 | 2.4 | UP |
| 8 | 1226 | Q9H254 | Spectrin beta chain, non-erythrocytic 4 | SPTN4_HUMAN | 0.01 | 2.3 | UP |
| 5 | 1797 | P13645 | Keratin, type I cytoskeletal 10 | K1C10_HUMAN | 0.007 | 2.2 | UP |
| 42 | 1260 | P02647 | Apolipoprotein A-I | APOA1_HUMAN | 0.04 | 2.1 | UP |
| 20 | 1427 | Q9BZ29 | Dedicator of cytokinesis protein 9 | DOCK9_HUMAN | 0.02 | 2.1 | UP |
| 58 | 886 | P00738 | Haptoglobin | HPT_HUMAN | 0.02 | 2.1 | UP |
| 112 | 676 | P02749 | Beta-2-glycoprotein 1 | APOH_HUMAN | 0.05 | 2 | UP |
| 40 | 1475 | Q3L8U1 | Chromodomain-helicase-DNA-binding protein 9 | CHD9_HUMAN | 0.04 | 2.0 | UP |
| 30 | 334 | P02790 | Hemopexin | HEMO_HUMAN | 0.04 | 2 | UP |
| 46 | 1705 | O15226 | NF-kappa-B-repressing factor | NKRF_HUMAN | 0.0 | 2 | UP |
| 41 | 1295 | Q969Q6 | Serine/threonine-protein phosphatase 2A regulatory subunit B’’ subunit gamma | P2R3C_HUMAN | 0.05 | 2.0 | UP |
| 93 | 1908 | P02647 | Apolipoprotein A-I | APOA1_HUMAN | 0.03 | 1.9 | UP |
| 64 | 903 | O95389 | Cellular communication network factor 6 | WISP3_HUMAN | 0.03 | 1.9 | UP |
| 77 | 953 | P00738 | Haptoglobin | HPT_HUMAN | 0.04 | 1.9 | UP |
| 99 | 796 | P00738 | Haptoglobin | HPT_HUMAN | 0.02 | 1.9 | UP |
| 9 | 1487 | P01834 | Immunoglobulin kappa constant | IGKC_HUMAN | 0.01 | 1.9 | UP |
| 36 | 1231 | P13645 | Keratin, type I cytoskeletal 10 | K1C10_HUMAN | 0.04 | 1.9 | UP |
| 100 | 987 | Q709C8 | Vacuolar protein sorting-associated protein 13C | VP13C_HUMAN | 0.03 | 1.9 | UP |
| 92 | 981 | P06727 | Apolipoprotein A-IV | APOA4_HUMAN | 0.04 | 1.8 | UP |
| 96 | 623 | Q92608 | Dedicator of cytokinesis protein 2 | DOCK2_HUMAN | 0.02 | 1.8 | UP |
| 21 | 1120 | O75648 | Mitochondrial tRNA-specific 2-thiouridylase 1 | MTU1_HUMAN | 0.02 | 1.8 | UP |
| 6 | 1937 | Q5T6V5 | Queuosine salvage protein | CI064_HUMAN | 0.008 | 1.8 | UP |
| 2 | 1792 | P02766 | Transthyretin | TTHY_HUMAN | 0.005 | 1.8 | UP |
| 76 | 1865 | P02766 | Transthyretin | TTHY_HUMAN | 0.04 | 1.8 | UP |
| 59 | 457 | Q96M63 | Coiled-coil domain-containing protein 114 | CC114_HUMAN | 0.05 | 1.7 | UP |
| 61 | 1305 | Q12852 | Mitogen-activated protein kinase kinase kinase 12 | M3K12_HUMAN | 0.03 | 1.7 | UP |
| 83 | 974 | P35558 | Phosphoenolpyruvate carboxykinase, cytosolic [GTP] | PCKGC_HUMAN | 0.05 | 1.7 | UP |
| 115 | 714 | Q02156 | Protein kinase C epsilon type | KPCE_HUMAN | 0.04 | 1.7 | UP |
| 22 | 1759 | P02766 | Transthyretin | TTHY_HUMAN | 0.02 | 1.7 | UP |
| 67 | 1067 | Q9NQW7 | Xaa-Pro aminopeptidase 1 | XPP1_HUMAN | 0.03 | 1.7 | UP |
| 80 | 1659 | P02647 | Apolipoprotein A-I | APOA1_HUMAN | 0.04 | 1.6 | UP |
| 57 | 1446 | Q9BV73 | Centrosome-associated protein CEP250 | CP250_HUMAN | 0.03 | 1.6 | UP |
| 12 | 1136 | O75636 | Ficolin-3 | FCN3_HUMAN | 0.02 | 1.6 | UP |
| 51 | 1277 | O75636 | Ficolin-3 | FCN3_HUMAN | 0.05 | 1.6 | UP |
| 47 | 1132 | Q92896 | Golgi apparatus protein 1 | GSLG1_HUMAN | 0.05 | 1.6 | UP |
| 118 | 797 | P00738 | Haptoglobin | HPT_HUMAN | 0.04 | 1.6 | UP |
| 44 | 1962 | P13645 | Keratin, type I cytoskeletal 10 | K1C10_HUMAN | 0.04 | 1.6 | UP |
| 23 | 1193 | P04264 | Keratin, type II cytoskeletal 1 | K2C1_HUMAN | 0.02 | 1.6 | UP |
| 87 | 955 | Q5BJF6 | Outer dense fiber protein 2 | ODFP2_HUMAN | 0.05 | 1.6 | UP |
| 39 | 1784 | P02753 | Retinol-binding protein 4 | RET4_HUMAN | 0.04 | 1.6 | UP |
| 56 | 1669 | P02753 | Retinol-binding protein 4 | RET4_HUMAN | 0.02 | 1.6 | UP |
| 35 | 1473 | Q9HBT8 | Zinc finger protein 286A | Z286A_HUMAN | 0.04 | 1.6 | UP |
| 55 | 1138 | Q6ZN19 | Zinc finger protein 841 | ZN841_HUMAN | 0.02 | 1.6 | UP |
| 105 | 788 | A8TX70 | Collagen alpha-5(VI) chain | CO6A5_HUMAN | 0.04 | 1.5 | UP |
| 97 | 771 | Q96EH8 | E3 ubiquitin-protein ligase NEURL3 | LINCR_HUMAN | 0.05 | 1.5 | UP |
| 85 | 1211 | P04264 | Keratin, type II cytoskeletal 1 | K2C1_HUMAN | 0.05 | 1.5 | UP |
| 90 | 913 | P02538 | Keratin, type II cytoskeletal 6A | K2C6A_HUMAN | 0.04 | 1.5 | UP |
| 89 | 1039 | P78332 | RNA-binding protein 6 | RBM6_HUMAN | 0.05 | 1.5 | UP |
| 32 | 1563 | P18206 | Vinculin | VINC_HUMAN | 0.04 | 1.5 | UP |
| Downregulated Proteins | |||||||
| 88 | 2217 | P68871 | Hemoglobin subunit beta | HBB_HUMAN | 0.05 | −5.0 | DOWN |
| 94 | 2323 | P02042 | Hemoglobin subunit delta | HBD_HUMAN | 0.03 | −4.2 | DOWN |
| 82 | 562 | P19652 | Alpha-1-acid glycoprotein 2 | A1AG2_HUMAN | 0.04 | −3.7 | DOWN |
| 95 | 481 | Q5R372 | Rab GTPase-activating protein 1-like | RBG1L_HUMAN | 0.02 | −2.9 | DOWN |
| 34 | 1448 | Q6ZNG0 | Zinc finger protein 620 | ZN620_HUMAN | 0.04 | −2.6 | DOWN |
| 45 | 456 | Q9HAC8 | Ubiquitin domain-containing protein 1 | UBTD1_HUMAN | 0.04 | −2.6 | DOWN |
| 31 | 450 | P01009 | Alpha-1-antitrypsin | A1AT_HUMAN | 0.04 | −2.5 | DOWN |
| 18 | 385 | P02787 | Serotransferrin | TRFE_HUMAN | 0.02 | −2.4 | DOWN |
| 7 | 1082 | O43300 | Leucine-rich repeat transmembrane neuronal protein 2 | LRRT2_HUMAN | 0.009 | −2.3 | DOWN |
| 11 | 382 | P02787 | Serotransferrin | TRFE_HUMAN | 0.01 | −2.2 | DOWN |
| 33 | 853 | P00738 | Haptoglobin | HPT_HUMAN | 0.04 | −2.2 | DOWN |
| 4 | 860 | P02787 | Serotransferrin | TRFE_HUMAN | 0.006 | −1.9 | DOWN |
| 14 | 865 | P00738 | Haptoglobin | HPT_HUMAN | 0.01 | −1.9 | DOWN |
| 17 | 857 | Q07065 | Cytoskeleton-associated protein 4 | CKAP4_HUMAN | 0.01 | −1.9 | DOWN |
| 27 | 235 | P17039 | Zinc finger protein 30 | ZNF30_HUMAN | 0.026 | −1.9 | DOWN |
| 38 | 497 | P01009 | Alpha-1-antitrypsin | A1AT_HUMAN | 0.04 | −1.9 | DOWN |
| 48 | 1877 | P20472 | Parvalbumin alpha | PRVA_HUMAN | 0.05 | −1.9 | DOWN |
| 62 | 1394 | Q14204 | Cytoplasmic dynein 1 heavy chain 1 | DYHC1_HUMAN | 0.03 | −1.9 | DOWN |
| 73 | 1106 | Q8TD57 | Dynein heavy chain 3, axonemal | DYH3_HUMAN | 0.04 | −1.9 | DOWN |
| 13 | 7 | P02774 | Vitamin D-binding protein | VTDB_HUMAN | 0.02 | −1.8 | DOWN |
| 72 | 388 | P00751 | Complement factor B | CFAB_HUMAN | 0.03 | −1.8 | DOWN |
| 3 | 202 | P21580 | Tumor necrosis factor alpha-induced protein 3 | TNAP3_HUMAN | 0.005 | −1.7 | DOWN |
| 29 | 661 | P01009 | Alpha-1-antitrypsin | A1AT_HUMAN | 0.04 | −1.7 | DOWN |
| 43 | 636 | Q99661 | Kinesin-like protein KIF2C | KIF2C_HUMAN | 0.04 | −1.7 | DOWN |
| 52 | 354 | O60384 | Putative zinc finger protein 861 | YS022_HUMAN | 0.05 | −1.7 | DOWN |
| 53 | 650 | P01009 | Alpha-1-antitrypsin | A1AT_HUMAN | 0.05 | −1.7 | DOWN |
| 68 | 423 | Q5R372 | Rab GTPase-activating protein 1-like | RBG1L_HUMAN | 0.03 | −1.7 | DOWN |
| 71 | 805 | P04637 | Cellular tumor antigen p53 | P53_HUMAN | 0.03 | −1.7 | DOWN |
| 98 | 341 | Q9P219 | Protein Daple | DAPLE_HUMAN | 0.05 | −1.7 | DOWN |
| 104 | 1465 | P17017 | Zinc finger protein 14 | ZNF14_HUMAN | 0.03 | −1.7 | DOWN |
| 15 | 864 | P01024 | Complement C3 | CO3_HUMAN | 0.01 | −1.6 | DOWN |
| 24 | 108 | O75121 | Microfibrillar-associated protein 3-like | MFA3L_HUMAN | 0.02 | −1.6 | DOWN |
| 26 | 266 | P02768 | Albumin | ALBU_HUMAN | 0.03 | −1.6 | DOWN |
| 28 | 357 | P08631 | Tyrosine protein kinase HCK | HCK_HUMAN | 0.03 | −1.6 | DOWN |
| 37 | 548 | P02765 | Alpha-2-HS-glycoprotein | FETUA_HUMAN | 0.04 | −1.6 | DOWN |
| 54 | 1037 | Q14683 | Structural maintenance of chromosomes protein 1A | SMC1A_HUMAN | 0.05 | −1.6 | DOWN |
| 66 | 1083 | Q9UKX3 | Myosin-13 | MYH13_HUMAN | 0.03 | −1.6 | DOWN |
| 74 | 579 | P02768 | Albumin | ALBU_HUMAN | 0.04 | −1.6 | DOWN |
| 86 | 1054 | Q9H792 | Inactive tyrosine protein kinase PEAK1 | SG269_HUMAN | 0.05 | −1.6 | DOWN |
| 102 | 702 | Q9UM13 | Anaphase-promoting complex subunit 10 | APC10_HUMAN | 0.03 | −1.6 | DOWN |
| 103 | 533 | P02774 | Vitamin D-binding protein | VTDB_HUMAN | 0.03 | −1.6 | DOWN |
| 108 | 633 | P02768 | Albumin | ALBU_HUMAN | 0.05 | −1.6 | DOWN |
| 114 | 537 | P02768 | Albumin | ALBU_HUMAN | 0.05 | −1.6 | DOWN |
| 120 | 550 | Q8TE73 | Dynein heavy chain 5, axonemal | DYH5_HUMAN | 0.05 | −1.6 | DOWN |
| 1 | 253 | Q9H792 | Inactive tyrosine protein kinase PEAK1 | SG269_HUMAN | 0.004 | −1.5 | DOWN |
| 19 | 820 | Q14585 | Zinc finger protein 345 | ZN345_HUMAN | 0.02 | −1.5 | DOWN |
| 49 | 504 | Q08999 | Retinoblastoma-like protein 2 | RBL2_HUMAN | 0.05 | −1.5 | DOWN |
| 50 | 1484 | P02768 | Albumin | ALBU_HUMAN | 0.05 | −1.5 | DOWN |
| 60 | 522 | Q9Y473 | Zinc finger protein 175 | ZN175_HUMAN | 0.03 | −1.5 | DOWN |
| 63 | 48 | P49792 | E3 SUMO-protein ligase RanBP2 | RBP2_HUMAN | 0.03 | −1.5 | DOWN |
| 65 | 28 | Q14624 | Inter-alpha-trypsin inhibitor heavy chain H4 | ITIH4_HUMAN | 0.03 | −1.5 | DOWN |
| 69 | 1213 | Q14683 | Structural maintenance of chromosomes protein 1A | SMC1A_HUMAN | 0.03 | −1.5 | DOWN |
| 75 | 215 | Q9Y4I1 | Unconventional myosin-Va | MYO5A_HUMAN | 0.04 | −1.5 | DOWN |
| 78 | 528 | P02768 | Albumin | ALBU_HUMAN | 0.04 | −1.5 | DOWN |
| 91 | 604 | Q6AW86 | Zinc finger protein 324B | Z324B_HUMAN | 0.04 | −1.5 | DOWN |
| 101 | 732 | P25311 | Zinc-alpha-2-glycoprotein | ZA2G_HUMAN | 0.03 | −1.5 | DOWN |
| 106 | 584 | Q8TD57 | Dynein heavy chain 3, axonemal | DYH3_HUMAN | 0.04 | −1.5 | DOWN |
| 107 | 1688 | P20472 | Parvalbumin alpha | PRVA_HUMAN | 0.04 | −1.5 | DOWN |
| 109 | 709 | Q9NZM1 | Myoferlin | MYOF_HUMAN | 0.05 | −1.5 | DOWN |
| 110 | 198 | P02768 | Albumin | ALBU_HUMAN | 0.05 | −1.5 | DOWN |
| 111 | 743 | P01009 | Alpha-1-antitrypsin | A1AT_HUMAN | 0.05 | −1.5 | DOWN |
| 113 | 556 | P20929 | Nebulin | NEBU_HUMAN | 0.05 | −1.5 | DOWN |
| 116 | 588 | Q96SZ6 | Mitochondrial tRNA methylthiotransferase CDK5RAP1 | CK5P1_HUMAN | 0.04 | −1.5 | DOWN |
| 117 | 275 | Q9Y473 | Zinc finger protein 175 | ZN175_HUMAN | 0.05 | −1.5 | DOWN |
| 119 | 768 | O76041 | Nebulette | NEBL_HUMAN | 0.04 | −1.5 | DOWN |
| 121 | 549 | P51508 | Zinc finger protein 81 | ZNF81_HUMAN | 0.03 | −1.5 | DOWN |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alasmari, F.; Alsanea, S.; Alfadda, A.A.; Alanazi, I.O.; Musambil, M.; Masood, A.; Alqahtani, F.; Fantoukh, O.I.; Alasmari, A.F.; Benabdelkamel, H. Serum Proteomic Analysis of Cannabis Use Disorder in Male Patients. Molecules 2021, 26, 5311. https://doi.org/10.3390/molecules26175311
Alasmari F, Alsanea S, Alfadda AA, Alanazi IO, Musambil M, Masood A, Alqahtani F, Fantoukh OI, Alasmari AF, Benabdelkamel H. Serum Proteomic Analysis of Cannabis Use Disorder in Male Patients. Molecules. 2021; 26(17):5311. https://doi.org/10.3390/molecules26175311
Chicago/Turabian StyleAlasmari, Fawaz, Sary Alsanea, Assim A. Alfadda, Ibrahim O. Alanazi, Mohthash Musambil, Afshan Masood, Faleh Alqahtani, Omer I. Fantoukh, Abdullah F. Alasmari, and Hicham Benabdelkamel. 2021. "Serum Proteomic Analysis of Cannabis Use Disorder in Male Patients" Molecules 26, no. 17: 5311. https://doi.org/10.3390/molecules26175311
APA StyleAlasmari, F., Alsanea, S., Alfadda, A. A., Alanazi, I. O., Musambil, M., Masood, A., Alqahtani, F., Fantoukh, O. I., Alasmari, A. F., & Benabdelkamel, H. (2021). Serum Proteomic Analysis of Cannabis Use Disorder in Male Patients. Molecules, 26(17), 5311. https://doi.org/10.3390/molecules26175311