
Curcumin as a Natural Remedy for Atherosclerosis: A Pharmacological Review



Abstract
:1. Introduction
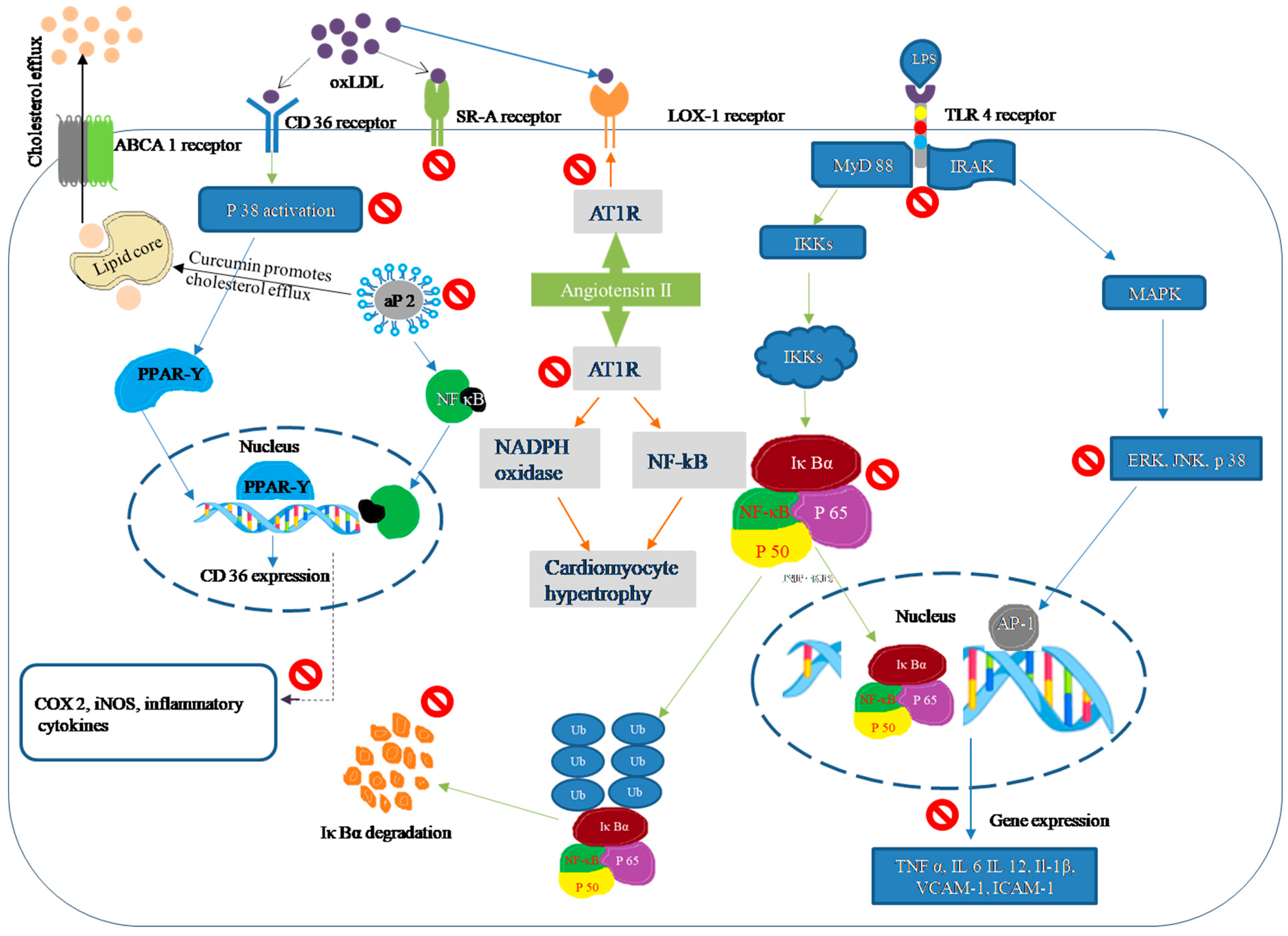
2. Atheroprotective Effects of Curcumin In Vitro
3. Atheroprotective Effects of Curcumin In Vivo
4. Clinical Studies of Curcumin
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hannawi, S.; Hannawi, H.; Al Salmi, I. Cardiovascular disease and subclinical atherosclerosis in rheumatoid arthritis. Hypertens. Res. 2020, 43, 982–984. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.; Fu, Y.; Li, B. Brain-derived neurotrophic factor alleviates diabetes mellitus-accelerated atherosclerosis by promoting M2 polarization of macrophages through repressing the STAT3 pathway. Cell. Signal. 2020, 70, 109569. [Google Scholar] [CrossRef]
- Ito, K.; Maeda, T.; Tada, K.; Takahashi, K.; Yasuno, T.; Masutani, K.; Mukoubara, S.; Arima, H.; Nakashima, H. The role of cigarette smoking on new-onset of chronic kidney disease in a Japanese population without prior chronic kidney disease: Iki epidemiological study of atherosclerosis and chronic kidney disease (ISSA-CKD). Clin. Exp. Nephrol. 2020, 24, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Zang, X.; Cui, X.; Zhang, J. Autophagy, Hyperlipidemia, and Atherosclerosis. Adv. Exp. Med. Biol. 2020, 1207, 237–264. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Wang, Y.; Wang, Z.; Liu, L.; Yang, Z.; Wang, M.; Xu, Y.; Ye, D.; Zhang, J.; Zhou, Q.; et al. The Expression of IL-12 Family Members in Patients with Hypertension and Its Association with the Occurrence of Carotid Atherosclerosis. Mediat. Inflamm. 2020, 2020, 2369279. [Google Scholar] [CrossRef] [Green Version]
- Ntaios, G.; Pearce, L.A.; Meseguer, E.; Endres, M.; Amarenco, P.; Ozturk, S.; Lang, W.; Bornstein, N.M.; Molina, C.A.; Pagola, J.; et al. Aortic Arch Atherosclerosis in Patients with Embolic Stroke of Undetermined Source: An Exploratory Analysis of the NAVIGATE ESUS Trial. Stroke 2019, 50, 3184–3190. [Google Scholar] [CrossRef]
- Pomozova, T.P.; Lykov, Y.V.; Komarova, I.S.; Dyatlov, N.V.; Zhelnov, V.V. Clinical and laboratory features of primary acute myocardial infarction in patients with obstructive and non-obstructive coronary atherosclerosis. Kardiologiia 2019, 59, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, K.S. Effects of Panax ginseng extract on lipid metabolism in humans. Pharmacol. Res. 2003, 48, 511–513. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, J.; Shi, Y.; Grimsgaard, S.; Alraek, T.; Fønnebø, V. Chinese red yeast rice (Monascus purpureus) for primary hyperlipidemia: A meta-analysis of randomized controlled trials. Chin. Med. 2006, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Tortosa, M.C.; Mesa, M.D.; Aguilera, M.C.; Quiles, J.L.; Baró, L.; Ramirez-Tortosa, C.L.; Martinez-Victoria, E.; Gil, A. Oral administration of a turmeric extract inhibits LDL oxidation and has hypocholesterolemic effects in rabbits with experimental atherosclerosis. Atherosclerosis 1999, 147, 371–378. [Google Scholar] [CrossRef]
- Chan, G.H.-H.; Law, B.Y.-K.; Chu, J.M.-T.; Yue, K.K.-M.; Jiang, Z.-H.; Lau, C.-W.; Huang, Y.; Chan, S.-W.; Ying-Kit, Y.P.; Wong, R.N.-S. Ginseng extracts restore high-glucose induced vascular dysfunctions by altering triglyceride metabolism and downregulation of atherosclerosis-related genes. Evid. Based. Complement. Altern. Med. 2013, 2013, 797310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Yang, H.J.; Kim, Y.J.; Park, S.; Lee, O.H.; Kim, K.S.; Kim, M.J. Korean turmeric is effective for dyslipidemia in human intervention study. J. Ethn. Foods 2016, 3, 213–221. [Google Scholar] [CrossRef] [Green Version]
- González-Ortega, L.A.; Acosta-Osorio, A.A.; Grube-Pagola, P.; Palmeros-Exsome, C.; Cano-Sarmiento, C.; García-Varela, R.; García, H.S. Anti-inflammatory Activity of Curcumin in Gel Carriers on Mice with Atrial Edema. J. Oleo Sci. 2020, 69, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Huang, L.; Gong, J.; Shen, S.; Huang, J.; Ren, H.; Hu, H. Efficacy and safety of turmeric and curcumin in lowering blood lipid levels in patients with cardiovascular risk factors: A meta-analysis of randomized controlled trials. Nutr. J. 2017, 16, 68. [Google Scholar] [CrossRef] [PubMed]
- Song, H.-C.; Chen, Y.; Chen, Y.; Park, J.; Zheng, M.; Surh, Y.-J.; Kim, U.-H.; Park, J.W.; Yu, R.; Chung, H.T.; et al. GSK-3β inhibition by curcumin mitigates amyloidogenesis via TFEB activation and anti-oxidative activity in human neuroblastoma cells. Free Radic. Res. 2020, 1–13. [Google Scholar] [CrossRef]
- Zhang, S.; Zou, J.; Li, P.; Zheng, X.; Feng, D. Curcumin Protects against Atherosclerosis in Apolipoprotein E-Knockout Mice by Inhibiting Toll-like Receptor 4 Expression. J. Agric. Food Chem. 2018, 66, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Victor, V.M.; Mauricio, M.D. Targeting Early Atherosclerosis: A Focus on Oxidative Stress and Inflammation. Oxidative Med. Cell. Longev. 2019, 1, 32. [Google Scholar] [CrossRef]
- Chen, Y.-Q.; Chai, Y.-S.; Xie, K.; Yu, F.; Wang, C.-J.; Lin, S.-H.; Yang, Y.-Z.; Xu, F. Curcumin Promotes the Expression of IL-35 by Regulating Regulatory T Cell Differentiation and Restrains Uncontrolled Inflammation and Lung Injury in Mice. Inflammation 2020, 43, 1913–1924. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, X.; Wang, X.; Liu, B.; Yuan, Y.; Zuo, X. Curcumin attenuates inflammation and cell apoptosis through regulating NF-κB and JAK2/STAT3 signaling pathway against acute kidney injury. Cell Cycle 2020, 19, 1941–1951. [Google Scholar] [CrossRef]
- Olszanecki, R.; Jawień, J.; Gajda, M.; Mateuszuk, L.; Gebska, A.; Korabiowska, M.; Chłopicki, S.; Korbut, R. Effect of Curcumin on atherosclerosis in apoE/LDLR-double knockout mice. J. Physiol. Pharmacol. 2005, 56, 627–635. [Google Scholar]
- Xie, M.; Tang, Q.; Nie, J.; Zhang, C.; Zhou, X.; Yu, S.; Sun, J.; Cheng, X.; Dong, N.; Hu, Y.; et al. BMAL1-Downregulation Aggravates Porphyromonas Gingivalis-Induced Atherosclerosis by Encouraging Oxidative Stress. Circ. Res. 2020, 126, e15–e29. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–327. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Goel, A.; Mehta, J.L. LOX-1: Regulation, Signaling and Its Role in Atherosclerosis. Antioxidants 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Boonla, O.; Kukongviriyapan, U.; Pakdeechote, P.; Kukongviriyapan, V.; Pannangpetch, P.; Prachaney, P.; Greenwald, S.E. Curcumin improves endothelial dysfunction and vascular remodeling in 2K-1C hypertensive rats by raising nitric oxide availability and reducing oxidative stress. Nitric Oxide Biol. Chem. 2014, 42, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Kapakos, G.; Youreva, V.; Srivastava, A. Cardiovascular protection by curcumin: Molecular aspects. Indian J. Biochem. Biophys. 2012, 49, 306–315. [Google Scholar] [PubMed]
- Han, Y.; Sun, H.-J.; Tong, Y.; Chen, Y.-Z.; Ye, C.; Qiu, Y.; Zhang, F.; Chen, A.-D.; Qi, X.-H.; Chen, Q.; et al. Curcumin attenuates migration of vascular smooth muscle cells via inhibiting NFκB-mediated NLRP3 expression in spontaneously hypertensive rats. J. Nutr. Biochem. 2019, 72, 108212. [Google Scholar] [CrossRef]
- Horváth, R.; Cerný, J.; Benedík, J.J.; Hökl, J.; Jelínková, I.; Benedík, J. The possible role of human cytomegalovirus (HCMV) in the origin of atherosclerosis. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2000, 16, 17–24. [Google Scholar] [CrossRef]
- Shen, K.; Xu, L.; Chen, D.; Tang, W.; Huang, Y. Human cytomegalovirus-encoded miR-UL112 contributes to HCMV-mediated vascular diseases by inducing vascular endothelial cell dysfunction. Virus Genes 2018, 54, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.-L.; Jia, Y.; Wan, Z.; An, Z.-L.; Yang, S.; Han, F.-F.; Gong, L.-L.; Xuan, L.-L.; Ren, L.-L.; Zhang, W.; et al. Curcumin inhibits the formation of atherosclerosis in ApoE(-/-) mice by suppressing cytomegalovirus activity in endothelial cells. Life Sci. 2020, 257, 117658. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713. [Google Scholar] [CrossRef]
- Di Pietro, N.; Formoso, G.; Pandolfi, A. Physiology and pathophysiology of oxLDL uptake by vascular wall cells in atherosclerosis. Vasc. Pharmacol. 2016, 84, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Khan, H.; Mirzaei, H. Mechanics insights of Curcumin in myocardial ischemia: Where are we standing? Eur. J. Med. Chem. 2019, 183, 111658. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Miao, X.; Li, F.; Adhikari, B.K.; Liu, Y.; Sun, J.; Zhang, R.; Cai, L.; Liu, Q.; Wang, Y. Curcuminoids: Implication for inflammation and oxidative stress in cardiovascular diseases. The clinical studies of Curcumin should be summarized and added in the last chapter. Phytother. Res. 2019, 33, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Hu, Y.; Zhao, Y.; Cheng, M.; Qin, H.; Cheng, T.; Wang, Q.; Peng, X.; Zhang, X. Curcumin Attenuates Titanium Particle-Induced Inflammation by Regulating Macrophage Polarization In Vitro and In Vivo. Front. Immunol. 2017, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momtazi-Borojeni, A.A.; Banach, M.; Majeed, M.; Sahebkar, A. P5330 Evaluating lipid-lowering and anti-atherogenic effect of injectable Curcumin in a rabbit model of atherosclerosis. Eur. Heart J. 2019, 40, ehz746-0299. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, Y.; Wang, F.; Tian, N.; Fan, C. Effect of Curcumin on down-expression of thrombospondin-4 induced by oxidized low-density lipoprotein in mouse macrophages. Biomed. Mater. Eng. 2014, 24, 181–189. [Google Scholar] [CrossRef]
- Chen, F.-Y.; Zhou, J.; Guo, N.; Ma, W.-G.; Huang, X.; Wang, H.; Yuan, Z.-Y. Curcumin retunes cholesterol transport homeostasis and inflammation response in M1 macrophage to prevent atherosclerosis. Biochem. Biophys. Res. Commun. 2015, 467, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Liu, M.-H.; Hu, H.-J.; Feng, H.; Fan, X.-J.; Zou, W.; Pan, Y.; Hu, X.; Wang, Z. Curcumin enhanced cholesterol efflux by upregulating ABCA1 expression through AMPK-SIRT1-LXRα signaling in THP-1 macrophage-derived foam cells. DNA Cell Biol. 2015, 34, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Sreedhar, R.; Giridharan, V.V.; Afrin, R.; Harima, M.; Miyashita, S.; Hara, M.; Suzuki, K.; et al. Curcumin alleviates renal dysfunction and suppresses inflammation by shifting from M1 to M2 macrophage polarization in daunorubicin induced nephrotoxicity in rats. Cytokine 2016, 84, 1–9. [Google Scholar] [CrossRef]
- Gao, S.; Zhou, J.; Liu, N.; Wang, L.; Gao, Q.; Wu, Y.; Zhao, Q.; Liu, P.; Wang, S.; Liu, Y.; et al. Curcumin induces M2 macrophage polarization by secretion IL-4 and/or IL-13. J. Mol. Cell. Cardiol. 2015, 85, 131–139. [Google Scholar] [CrossRef]
- Karimian, M.S.; Pirro, M.; Majeed, M.; Sahebkar, A. Curcumin as a natural regulator of monocyte chemoattractant protein-1. Cytokine Growth Factor Rev. 2017, 63, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tian, L.; Sun, D.; Yin, D. Curcumin ameliorates atherosclerosis through upregulation of miR-126. J. Cell. Physiol. 2019, 234, 21049–21059. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, T.; Wang, X.; Wei, X.; Chen, Y.; Guo, L.; Zhang, J.; Wang, C. Curcumin Modulates Macrophage Polarization Through the Inhibition of the Toll-Like Receptor 4 Expression and its Signaling Pathways. Cell. Physiol. Biochem. 2015, 36, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Rains, J.; Croad, J.; Larson, B.; Jones, K. Curcumin supplementation lowers TNF-alpha, IL-6, IL-8, and MCP-1 secretion in high glucose-treated cultured monocytes and blood levels of TNF-alpha, IL-6, MCP-1, glucose, and glycosylated hemoglobin in diabetic rats. Antioxid. Redox Signal. 2009, 11, 241–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.T.; Zingg, J.-M.; Kwan, P.; Noble, T.; Smith, D.; Meydani, M. Curcumin modulation of high fat diet-induced atherosclerosis and steatohepatosis in LDL receptor deficient mice. Atherosclerosis 2014, 232, 40–51. [Google Scholar] [CrossRef]
- Youn, H.S.; Saitoh, S.I.; Miyake, K.; Hwang, D.H. Inhibition of homodimerization of Toll-like receptor 4 by curcumin. Biochem. Pharmacol. 2006, 72, 62–69. [Google Scholar] [CrossRef]
- Chen, F.; Guo, N.; Cao, G.; Zhou, J.; Yuan, Z. Molecular analysis of curcumin-induced polarization of murine RAW264.7 macrophages. J. Cardiovasc. Pharmacol. 2014, 63, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kang, Y.-X.; Pan, W.; Lei, W.; Feng, B.; Wang, X.-J. Enhancement of Anti-Inflammatory Activity of Curcumin Using Phosphatidylserine-Containing Nanoparticles in Cultured Macrophages. Int. J. Mol. Sci. 2016, 17, 969. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Yao, Y.-H.; Zhang, Z.-M.; Liu, J.-S.; Xiang, H. Curcumin inhibits hypoxia inducible factor-1α-induced inflammation and apoptosis in macrophages through an ERK dependent pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1816–1825. [Google Scholar] [CrossRef]
- Keshavarz, Z.; Kheirollah, A.; Ghaffari, M.-A.; Babaahmadi-Rezaei, H. Curcumin Inhibited Endothelin-1 mRNA Expression Induced by TGF-β in Bovine Aortic Endothelial Cell. Jundishapur J. Nat. Pharm. Prod. 2019, 14. [Google Scholar] [CrossRef]
- Ameruoso, A.; Palomba, R.; Palange, A.L.; Cervadoro, A.; Lee, A.; Di Mascolo, D.; Decuzzi, P. Ameliorating Amyloid-β Fibrils Triggered Inflammation via Curcumin-Loaded Polymeric Nanoconstructs. Front. Immunol. 2017, 8, 1411. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Ye, B.; Lin, L.; Tian, L.; Yang, H.; Wang, C.; Huang, W.; Huang, Z. Curcumin Alleviates oxLDL Induced MMP-9 and EMMPRIN Expression through the Inhibition of NF-κB and MAPK Pathways in Macrophages. Front. Pharmacol. 2017, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Han, Z.; Tian, L.; Chen, K.; Fan, Y.; Ye, B.; Huang, W.; Wang, C.; Huang, Z. Curcumin inhibits EMMPRIN and MMP-9 expression through AMPK-MAPK and PKC signaling in PMA induced macrophages. J. Transl. Med. 2014, 12, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-L.; Chen, P.-Y.; Wu, M.-J.; Tai, M.-H.; Ho, C.-T.; Yen, J.-H. Curcuminoids Modulate the PKCδ/NADPH Oxidase/Reactive Oxygen Species Signaling Pathway and Suppress Matrix Invasion during Monocyte-Macrophage Differentiation. J. Agric. Food Chem. 2015, 63, 8838–8848. [Google Scholar] [CrossRef]
- Kong, F.; Ye, B.; Cao, J.; Cai, X.; Lin, L.; Huang, S.; Huang, W.; Huang, Z. Curcumin Represses NLRP3 Inflammasome Activation via TLR4/MyD88/NF-κB and P2X7R Signaling in PMA-Induced Macrophages. Front. Pharmacol. 2016, 7, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Sun, X.; Zhu, X.; Lv, F.; Zhong, Z.; Zhang, F.; Guo, W.; Cao, W.; Yang, L.; Tian, Y. Apoptosis of THP-1 derived macrophages induced by sonodynamic therapy using a new sonosensitizer hydroxyl acetylated curcumin. PLoS ONE 2014, 9, e93133. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Liu, T.; Guo, Z. Curcumin inhibits ox-LDL-induced MCP-1 expression by suppressing the p38MAPK and NF-κB pathways in rat vascular smooth muscle cells. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2012, 61, 61–67. [Google Scholar] [CrossRef]
- Hosseinzadeh, L.; Behravan, J.; Mosaffa, F.; Bahrami, G.; Bahrami, A.; Karimi, G. Curcumin potentiates doxorubicin-induced apoptosis in H9c2 cardiac muscle cells through generation of reactive oxygen species. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 1102–1109. [Google Scholar] [CrossRef]
- Meng, Z.; Yan, C.; Deng, Q.; Gao, D.; Niu, X. Curcumin inhibits LPS-induced inflammation in rat vascular smooth muscle cells in vitro via ROS-relative TLR4-MAPK/NF-κB pathways. Acta Pharmacol. Sin. 2013, 34, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Li, C.; Sun, H.; Luo, T.; Tan, Y.; Tian, D.; Guo, Z. Curcumin inhibits monocyte chemoattractant protein-1 expression and enhances cholesterol efflux by suppressing the c-Jun N-terminal kinase pathway in macrophage. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2014, 63, 841–850. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Curcumin-induced suppression of adipogenic differentiation is accompanied by activation of Wnt/beta-catenin signaling. Am. J. Physiol. Cell Physiol. 2010, 298, C1510-6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, J.; Pang, X.; Zhao, J.; Xu, S. Curcumin Suppresses Aldosterone-Induced CRP Generation in Rat Vascular Smooth Muscle Cells via Interfering with the ROS-ERK1/2 Signaling Pathway. Evid. Based. Complement. Altern. Med. 2020, 2020, 3245653. [Google Scholar] [CrossRef]
- Wu, T.; Xiang, Y.; Lv, Y.; Li, D.; Yu, L.; Guo, R. miR-590-3p mediates the protective effect of Curcumin on injured endothelial cells induced by angiotensin II. Am. J. Transl. Res. 2017, 9, 289–300. [Google Scholar] [PubMed]
- Ramaswami, G.; Chai, H.; Yao, Q.; Lin, P.H.; Lumsden, A.B.; Chen, C. Curcumin blocks homocysteine-induced endothelial dysfunction in porcine coronary arteries. J. Vasc. Surg. 2004, 40, 1216–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel-Dávalos, A.; Silva Sánchez, G.J.; Huerta-García, E.; Rueda-Romero, C.; Soca Chafre, G.; Mitre-Aguilar, I.B.; Alfaro-Moreno, E.; Pedraza-Chaverri, J.; López-Marure, R. Curcumin inhibits activation induced by urban particulate material or titanium dioxide nanoparticles in primary human endothelial cells. PLoS ONE 2017, 12, e0188169. [Google Scholar] [CrossRef]
- Lee, S.E.; Park, H.R.; Jeon, S.; Han, D.; Park, Y.S. Curcumin Attenuates Acrolein-induced COX-2 Expression and Prostaglandin Production in Human Umbilical Vein Endothelial Cells. J. Lipid Atheroscler. 2020, 9, 184–194. [Google Scholar] [CrossRef]
- Lee, H.-S.; Lee, M.-J.; Kim, H.; Choi, S.-K.; Kim, J.-E.; Moon, H.-I.; Park, W.-H. Curcumin inhibits TNFalpha-induced lectin-like oxidised LDL receptor-1 (LOX-1) expression and suppresses the inflammatory response in human umbilical vein endothelial cells (HUVECs) by an antioxidant mechanism. J. Enzym. Inhib. Med. Chem. 2010, 25, 720–729. [Google Scholar] [CrossRef]
- Kim, D.-C.; Ku, S.-K.; Lee, W.; Bae, J.-S. Barrier protective activities of Curcumin and its derivative. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2012, 61, 437–444. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ahn, Y.; Hong, M.H.; Joo, S.Y.; Kim, K.H.; Sohn, I.S.; Park, H.W.; Hong, Y.J.; Kim, J.H.; Kim, W.; et al. Curcumin attenuates inflammatory responses of TNF-alpha-stimulated human endothelial cells. J. Cardiovasc. Pharmacol. 2007, 50, 41–49. [Google Scholar] [CrossRef]
- Zhong, Y.; Feng, J.; Li, J.; Fan, Z. Curcumin prevents lipopolysaccharide-induced matrix metalloproteinase-2 activity via the Ras/MEK1/2 signaling pathway in rat vascular smooth muscle cells. Mol. Med. Rep. 2017, 16, 4315–4319. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Xie, H.; Chen, J.; Liu, S. Curcumin inhibits proliferation, migration and neointimal formation of vascular smooth muscle via activating miR-22. Pharm. Biol. 2020, 58, 610–619. [Google Scholar] [CrossRef]
- Yuan, H.-Y.; Kuang, S.-Y.; Zheng, X.; Ling, H.-Y.; Yang, Y.-B.; Yan, P.-K.; Li, K.; Liao, D.-F. Curcumin inhibits cellular cholesterol accumulation by regulating SREBP-1/caveolin-1 signaling pathway in vascular smooth muscle cells. Acta Pharmacol. Sin. 2008, 29, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Den Dekker, W.K.; Cheng, C.; Pasterkamp, G.; Duckers, H.J. Toll like receptor 4 in atherosclerosis and plaque destabilization. Atherosclerosis 2010, 209, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Lee, E.N.; Park, J.K.; Lee, J.-R.; Kim, J.-H.; Choi, H.-J.; Kim, B.-S.; Lee, H.-W.; Lee, K.-S.; Yoon, S. Curcumin attenuates TNF-α-induced expression of intercellular adhesion molecule-1, vascular cell adhesion molecule-1 and proinflammatory cytokines in human endometriotic stromal cells. Phytother. Res. 2012, 26, 1037–1047. [Google Scholar] [CrossRef]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Ogura, S.; Little, P.J.; Xu, S.; Sawamura, T. Targeting LOX-1 in atherosclerosis and vasculopathy: Current knowledge and future perspectives. Ann. N.Y. Acad. Sci. 2019, 1443, 34–53. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Um, H.J.; Cho, K.-H.; Kwon, T.K. Curcumin inhibits oxLDL-induced CD36 expression and foam cell formation through the inhibition of p38 MAPK phosphorylation. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 58, 77–85. [Google Scholar] [CrossRef]
- Tontonoz, P.; Nagy, L.; Alvarez, J.G.A.; Thomazy, V.A.; Evans, R.M. PPARγ Promotes Monocyte/Macrophage Differentiation and Uptake of Oxidized LDL. Cell 1998, 93, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Chistiakov, D.A.; Bobryshev, Y.V.; Orekhov, A.N. Macrophage-mediated cholesterol handling in atherosclerosis. J. Cell. Mol. Med. 2016, 20, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Makowski, L.; Brittingham, K.C.; Reynolds, J.M.; Suttles, J.; Hotamisligil, G.S. The fatty acid-binding protein, aP2, coordinates macrophage cholesterol trafficking and inflammatory activity. Macrophage expression of aP2 impacts peroxisome proliferator-activated receptor gamma and IkappaB kinase activities. J. Biol. Chem. 2005, 280, 12888–12895. [Google Scholar] [CrossRef] [Green Version]
- Wessel, J.; Topol, E.J.; Ji, M.; Meyer, J.; McCarthy, J.J. Replication of the association between the thrombospondin-4 A387P polymorphism and myocardial infarction. Am. Heart J. 2004, 147, 905–909. [Google Scholar]
- Changal, K.H.; Khan, M.S.; Bashir, R.; Sheikh, M.A. Curcumin Preparations Can Improve Flow-Mediated Dilation and Endothelial Function: A Meta-Analysis. Complement. Med. Res. 2020, 27, 272–281. [Google Scholar] [CrossRef]
- Li, X.; Lu, Y.; Sun, Y.; Zhang, Q. Effect of Curcumin on permeability of coronary artery and expression of related proteins in rat coronary atherosclerosis heart disease model. Int. J. Clin. Exp. Pathol. 2015, 8, 7247–7253. [Google Scholar]
- Gao, S.; Zhang, W.; Zhao, Q.; Zhou, J.; Wu, Y.; Liu, Y.; Yuan, Z.; Wang, L. Curcumin ameliorates atherosclerosis in apolipoprotein E deficient asthmatic mice by regulating the balance of Th2/Treg cells. Phytomedicine 2019, 52, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.L.; Ghafil, F.A.; Fatima, G.; Hadi, N.R.; Mahdi, H.F. Anti-Atherosclerotic and Anti-Inflammatory Effects of Curcumin on Hypercholesterolemic Male Rabbits. Indian J. Clin. Biochem. 2021, 36, 74–80. [Google Scholar] [CrossRef]
- Amato, A.; Caldara, G.F.; Nuzzo, D.; Baldassano, S.; Picone, P.; Rizzo, M.; Mulè, F.; Di Carlo, M. NAFLD and atherosclerosis are prevented by a natural dietary supplement containing curcumin, silymarin, guggul, chlorogenic acid and inulin in mice fed a high-fat diet. Nutrients 2017, 9, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coban, D.; Milenkovic, D.; Chanet, A.; Khallou-Laschet, J.; Sabbe, L.; Palagani, A.; Vanden Berghe, W.; Mazur, A.; Morand, C. Dietary curcumin inhibits atherosclerosis by affecting the expression of genes involved in leukocyte adhesion and transendothelial migration. Mol. Nutr. Food Res. 2012, 56, 1270–1281. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Bie, J.; Wang, J.; Ghosh, S. Oral supplementation with non-absorbable antibiotics or Curcumin attenuates western diet-induced atherosclerosis and glucose intolerance in LDLR−/− mice–role of intestinal permeability and macrophage activation. PLoS ONE 2014, 9, e108577. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Wang, Y.; Cai, L.; Cai, Y.; Hu, J.; Yu, C.; Li, J.; Feng, Z.; Yang, S.; Li, X.; et al. Inhibition of high glucose-induced inflammatory response and macrophage infiltration by a novel Curcumin derivative prevents renal injury in diabetic rats. Br. J. Pharmacol. 2012, 166, 1169–1182. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Hong, J.H.; Jung, S.H.; Cho, K.H. Turmeric and laurel aqueous extracts exhibit in vitro anti-atherosclerotic activity and in vivo hypolipidemic effects in a zebrafish model. J. Med. Food 2011, 14, 247–256. [Google Scholar] [CrossRef]
- Shin, S.K.; Ha, T.Y.; McGregor, R.A.; Choi, M.S. Long-term Curcumin administration protects against atherosclerosis via hepatic regulation of lipoprotein cholesterol metabolism. Mol. Nutr. Food Res. 2011, 55, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Liu, Z.Y.; Yang, Y.P.; Liu, S.M. Effect of Curcumin on inhibiting atherogenesis by down-regulating lipocalin-2 expression in apolipoprotein E knockout mice. Bio-Med. Mater. Eng. 2016, 27, 577–587. [Google Scholar] [CrossRef]
- Zheng, B.; Yang, L.; Wen, C.; Huang, X.; Xu, C.; Lee, K.H.; Xu, J. Curcumin analog L3 alleviates diabetic atherosclerosis by multiple effects. Eur. J. Pharmacol. 2016, 775, 22–34. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, S.; Li, P.; Zheng, X.; Feng, D. Supplementation with Curcumin inhibits intestinal cholesterol absorption and prevents atherosclerosis in high-fat diet–fed apolipoprotein E knockout mice. Nutr. Res. 2018, 56, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Khalili, N.; Sahebi, E.; Namazi, S.; Reiner, Ž.; Majeed, M.; Sahebkar, A. Curcuminoids modify lipid profile in type 2 diabetes mellitus: A randomized controlled trial. Complementary Ther. Med. 2017, 33, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Kianpour, P.; Mohtashami, R.; Jafari, R.; Simental-Mendía, L.E.; Sahebkar, A. Curcumin lowers serum lipids and uric acid in subjects with nonalcoholic fatty liver disease: A randomized controlled trial. J. Cardiovasc. Pharmacol. 2016, 68, 223–229. [Google Scholar] [CrossRef]
- Ramırez-Boscá, A.; Soler, A.; Carrion, M.A.; Dıaz-Alperi, J.; Bernd, A.; Quintanilla, C.; Almagro, E.Q.; Miquel, J. An hydroalcoholic extract of Curcuma longa lowers the apo B/apo A ratio: Implications for atherogenesis prevention. Mech. Ageing Dev. 2000, 119, 41–47. [Google Scholar] [CrossRef]
- Wongcharoen, W.; Jai-Aue, S.; Phrommintikul, A.; Nawarawong, W.; Woragidpoonpol, S.; Tepsuwan, T.; Sukonthasarn, A.; Apaijai, N.; Chattipakorn, N. Effects of curcuminoids on frequency of acute myocardial infarction after coronary artery bypass grafting. Am. J. Cardiol. 2012, 110, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Funamoto, M.; Sunagawa, Y.; Katanasaka, Y.; Miyazaki, Y.; Imaizumi, A.; Kakeya, H.; Yamakage, H.; Satoh-Asahara, N.; Komiyama, M.; Wada, H.; et al. Highly absorptive Curcumin reduces serum atherosclerotic low-density lipoprotein levels in patients with mild COPD. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 2029. [Google Scholar]
- DiSilvestro, R.A.; Joseph, E.; Zhao, S.; Bomser, J. Diverse effects of a low dose supplement of lipidated Curcumin in healthy middle aged people. Nutr. J. 2012, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.M.; Stoner, L.; Rowlands, D.S.; Caldwell, A.R.; Sanders, E.; Kreutzer, A.; Mitchell, J.B.; Purpura, M.; Jäger, R. Novel form of Curcumin improves endothelial function in young, healthy individuals: A double-blind placebo controlled study. J. Nutr. Metab. 2016, 2016, 1089653. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.S.; Berrones, A.J.; Krishnakumar, I.M.; Charnigo, R.J.; Westgate, P.M.; Fleenor, B.S. Responsiveness to Curcumin intervention is associated with reduced aortic stiffness in young, obese men with higher initial stiffness. J. Funct. Foods 2017, 29, 154–160. [Google Scholar] [CrossRef]
- Zhou, Y.; Little, P.J.; Xu, S.; Kamato, D. Curcumin Inhibits Lysophosphatidic Acid Mediated MCP-1 Expression via Blocking ROCK Signalling. Molecules 2021, 26, 2320. [Google Scholar] [CrossRef] [PubMed]
- Helli, B.; Gerami, H.; Kavianpour, M.; Heybar, H.; Hosseini, S.K.; Haghighian, H.K. Curcumin Nanomicelle Improves Lipid Profile, Stress Oxidative Factors and Inflammatory Markers in Patients Undergoing Coronary Elective Angioplasty; A Randomized Clinical Trial. Endocr. Metab. Immune Disord. Drug Targets 2021. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, X.; Pi, C.; Yang, H.; Zheng, X.; Zhao, L.; Wei, Y. Review of Curcumin Physicochemical Targeting Delivery System. Int. J. Nanomed. 2020, 15, 9799–9821. [Google Scholar] [CrossRef]
- Pechanova, O.; Dayar, E.; Cebova, M. Therapeutic Potential of Polyphenols-Loaded Polymeric Nanoparticles in Cardiovascular System. Molecules 2020, 25, 3322. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Experimental Model | Concentration Used | Outcomes and Possible Mechanisms of Action | References |
|---|---|---|---|
| U937 monocytes | 0.01–1 µM |
| [44] |
| HMEC-1 cells | 0.1–10 μM |
| [42] |
| ANA-1 mouse macrophage cell line | 5–25 μM |
| [36] |
| RAW 264.7 macrophages |
| [34] | |
| H9c2 rat cardiac myoblasts | 5–40 μM |
| [36] |
| Human monocytic THP-1 cells | 7.5–30 μM |
| [43] |
| Human monocytic THP-1 cells | 5–20 μM |
| [45] |
| RAW264.7 macrophage | 6.25 and 12.5 μM |
| [37] |
| Ba/F3 cells | 10–20 μM |
| [46] |
| RAW264.7 macrophage | 6.25–25 μM |
| [47] |
| RAW264.7 macrophage | 6.25–50 μM |
| [40] |
| RAW264.7 macrophage | 6.25, and 25 nM |
| [34] |
| RAW264.7 macrophage | 8–128 μM |
| [48] |
| Mouse peritoneal macrophages | 10–50 μM |
| [16] |
| Human monocytic THP-1 cells | 20–40 μM |
| [49] |
| Bovine aortic endothelial cells (BAECs) | 5–15 μM |
| [50] |
| RAW264.7 macrophage | 0.1–30 μM |
| [51] |
| Human monocytic THP-1 cells | 0–50 μM |
| [52] |
| Human monocytic THP-1 cells | 0 to 100 μM |
| [53] |
| Human monocytic THP-1 cells | 10−20 μM |
| [54] |
| Human monocytic THP-1 cells | 0–50 μM |
| [55] |
| THP1-derived macrophage foam cells | 0–80 μM |
| [38] |
| Human monocytic THP-1 cells | 5.0 µg/mL |
| [56] |
| VSMCs | 5–30 μM |
| [57] |
| H9c2 embryonic rat heart derived cells | 5–15 μM |
| [58] |
| VSMCs | 5–30 μM |
| [59] |
| RAW264.7 macrophage | 0–40 μM |
| [60] |
| 3T3-L1 fibroblast cells | 0–30 μM |
| [61] |
| VSMCs | 1.25–5 μM |
| [62] |
| Endothelial cells | 10−5 M |
| [63] |
| Cultured porcine coronary artery rings | 5 μM |
| [64] |
| HUVEC cells | 1, 10,100 μM |
| [65] |
| HUVEC cells | 25 μM |
| [66] |
| HUVEC cells | 1–25 μM |
| [67] |
| HUVEC cells | 2.5–100 μM |
| [68] |
| HUVEC cells | 3–30 μM |
| [69] |
| VSMCs | 20–40 μM |
| [70] |
| VSMCs | - |
| [71] |
| VSMCs | 12.5–50 μM |
| [72] |
| HUVEC cells | 0.5–2 μM |
| [29] |
| VSMCs | 10–20 μM |
| [34] |
| VSMCs | 20 μM |
| [26] |
| In Vivo Experimental Model | Curcumin Concentration | Outcomes and Possible Mechanisms of Action | References |
|---|---|---|---|
| ApoE−/− mice | 0.1% w/w |
| [16] |
| Male New-Zealand rabbits | 1.66 mg/kg body weight |
| [10] |
| New Zealand white male rabbits | 10 mg/kg/week |
| [35] |
| Ldlr−/− mice | 500–1500 mg/kg |
| [45] |
| Male Wistar rats | 100 mg/(kg/d) curcumin |
| [83] |
| ApoE−/− mice | 200 mg/kg/d |
| [84] |
| Male Rabbits | 0.2% |
| [85] |
| ApoE/LDLR—doubleknockout mice | 0.3 mg/perday |
| [20] |
| Male C57BL/6J (B6) mice | 0.09 mg |
| [86] |
| ApoE−/− mice | 0.2% |
| [87] |
| LDLR−/− mice | 100 mg/kg |
| [88] |
| Sprague-Dawley rats | 100 mg/kg body weight |
| [44] |
| Sprague-Dawley rats | 0.2–5.0 mg/kg |
| [89] |
| Zebrafish | 10% wt/wt |
| [90] |
| ApoE−/− mice | 15–25mg/kg/d |
| [29] |
| ApoE−/− mice | 10 mg/kg |
| [59] |
| LDLR−/− mice | 0.02%w/w |
| [91] |
| ApoE−/− mice | 40, 60, and 80 mg/kg/d curcumin |
| [92] |
| Male ICR mice | 1–2mmol/kg/day |
| [93] |
| ApoE−/− mice | 0.1% w/w |
| [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, L.; Sharma, S.; Xu, S.; Tewari, D.; Fang, J. Curcumin as a Natural Remedy for Atherosclerosis: A Pharmacological Review. Molecules 2021, 26, 4036. https://doi.org/10.3390/molecules26134036
Singh L, Sharma S, Xu S, Tewari D, Fang J. Curcumin as a Natural Remedy for Atherosclerosis: A Pharmacological Review. Molecules. 2021; 26(13):4036. https://doi.org/10.3390/molecules26134036
Chicago/Turabian StyleSingh, Laxman, Shikha Sharma, Suowen Xu, Devesh Tewari, and Jian Fang. 2021. "Curcumin as a Natural Remedy for Atherosclerosis: A Pharmacological Review" Molecules 26, no. 13: 4036. https://doi.org/10.3390/molecules26134036
APA StyleSingh, L., Sharma, S., Xu, S., Tewari, D., & Fang, J. (2021). Curcumin as a Natural Remedy for Atherosclerosis: A Pharmacological Review. Molecules, 26(13), 4036. https://doi.org/10.3390/molecules26134036