
Characterization of βN-Octadecanoyl-5-hydroxytryptamide Anti-Inflammatory Effect

 ,
, 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. C18-5HT Synthesis
2.2. C18-5HT Does Not Affect Cell Viability and Inhibit Nitric Oxide and Cytokines Production
2.3. C18-5HT Also Presents an Effect When Orally Administered to Mice
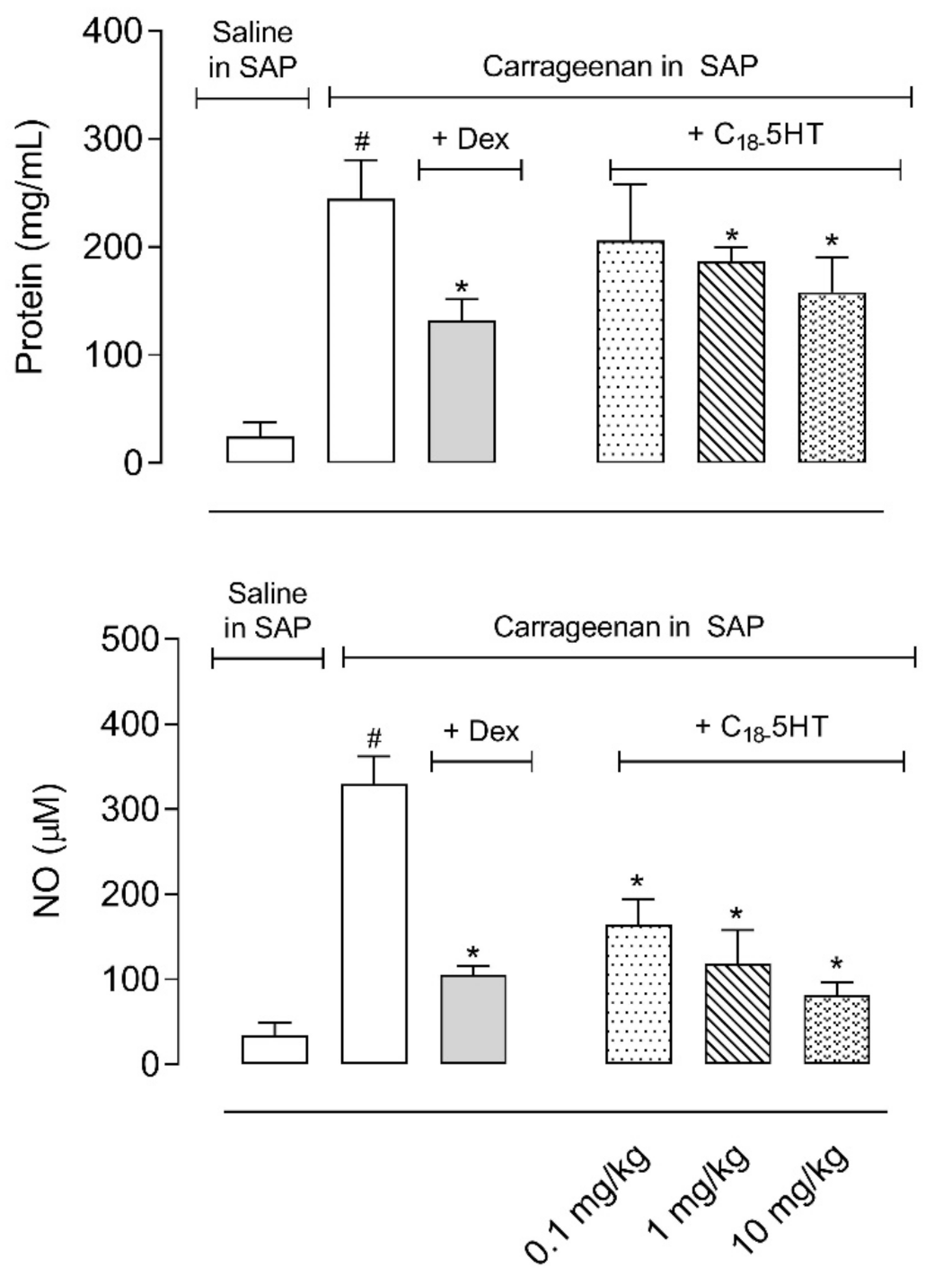
2.4. Pretreatment of Mice with C18-5HT Reduces Protein Extravasation, Nitric Oxide, and Cytokines Production
2.5. C18-5HT Reduces Reactive Oxygen Species Production by Leukocytes in an Ex Vivo Assay
3. Discussion
4. Materials and Methods
4.1. C18-5HT Synthesis
4.2. Materials and Reagents
4.3. Drugs and C18-5HT Administration
4.4. Cell Culture
4.5. Cell Viability
4.6. Animals
4.7. Carrageenan-Induced Cellular Migration into the Subcutaneous Air Pouch (SAP)
4.8. Nitrate, Nitrite, Protein, and Cytokines Measurement
4.9. Ex Vivo Reactive Oxygen Species (ROS) Measurement
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Meijerink, J.; Balvers, M.; Witkamp, R. N-Acyl amines of docosahexaenoic acid and other n-3 polyunsatured fatty acids-from fishy endocannabinoids to potential leads. Br. J. Pharmcol. 2013, 169, 772–783. [Google Scholar] [CrossRef]
- Di Marzo, V.; Bisogno, T.; De Petrocellis, L. Endocannabinoids and related compounds: Walking back and forth between plant natural products and animal physiology. Chem. Biol. 2007, 14, 741–756. [Google Scholar] [CrossRef]
- Alexander, S.P.H.; Mathie, A.; Peters, J.A. Guide to Receptors and Channels (GRAC), 5th edition. Br. J. Pharmcol. 2011, 164, S1–S324. [Google Scholar] [CrossRef]
- Maione, S.; Petrocellis, L.; Novellis, V.; Moriello, A.S.; Petrosino, S.; Palazzo, E.; Rossi, F.S.; Woodward, D.F.; Marzo, V. Analgesic actions of N -arachidonoyl-serotonin, a fatty acid amide hydrolase inhibitor with antagonistic activity at vanilloid TRPV1 receptors. Br. J. Pharmcol. 2007, 150, 766–781. [Google Scholar] [CrossRef] [PubMed]
- Poland, M.; ten Klooster, J.P.; Wang, Z.; Pieters, R.; Boekschoten, M.; Witkamp, R.; Meijerink, J. Docosahexaenoyl serotonin, an endogenously formed n-3 fatty acid-serotonin conjugate has anti-inflammatory properties by attenuating IL-23–IL-17 signaling in macrophages. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 2020–2028. [Google Scholar] [CrossRef] [PubMed]
- Giorno, T.B.S.; Moreira, I.G.D.S.; Rezende, C.M.; Fernandes, P.D. New βN-octadecanoyl-5-hydroxytryptamide: Antinociceptive effect and possible mechanism of action in mice. Sci. Rep. 2018, 8, 10027. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.L.; Lima, F.A.; Laura, A.; Brand, M.; Cunha, S.; Rezende, C.M.; Fernandes, P.D. Two New β N -Alkanoyl-5-Hydroxytryptamides with Relevant Antinociceptive Activity. Biomedines 2021, 9, 455. [Google Scholar] [CrossRef]
- Reddy, S.T.; Swamy, M.J. Synthesis, physicochemical characterization and membrane interactions of a homologous series of N-acylserotonins: Bioactive, endogenous conjugates of serotonin with fatty acids. Biochim. Biophys. Acta Biomembr. 2015, 1848, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Bardelmeier, I.; Weiss, C.; Rubach, M.; Somoza, V.; Hofmann, T. Quantitation of β N -Alkanoyl-5-hydroxytryptamides in Coffee by Means of LC-MS/MS-SIDA and Assessment of Their Gastric Acid Secretion Potential Using the HGT-1 Cell Assay. J. Agric. Food Chem. 2010, 58, 1593–1602. [Google Scholar] [CrossRef]
- Hunskaar, S.; Hole, K. The formalin test in mice: Dissociation between inflammatory and non-inflammatory pain. Pain 1987, 30, 103–114. [Google Scholar] [CrossRef]
- Wheeler-Aceto, H.; Porreca, F.; Cowan, A. The rat paw formalin test: Comparison of noxious agents. Pain 1990, 40, 229–238. [Google Scholar] [CrossRef]
- Jaffery, G.; Coleman, J.W.; Huntley, J.; Bell, E.B. Mast cell recovery following chronic treatment with compound 48/80. Int. Arch. Allergy Immunol. 1994, 105, 274–280. [Google Scholar] [CrossRef]
- Parada, C.A.; Tambeli, C.H.; Cunha, F.Q.; Ferreira, S.H. The major role of peripheral release of histamine and 5-hydroxytryptamine in formalin-induced nociception. Neuroscience 2001, 102, 937–944. [Google Scholar] [CrossRef]
- Lanni, C.; Becker, E.L. Release of phospholipase A2 activity from rabbit peritoneal neutrophils by f-Met-Leu-Phe. Am. J. Pathol. 1983, 113, 90–94. [Google Scholar] [PubMed]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Malech, H.L.; Gallin, J.I. Current concepts: Immunology. Neutrophils in human diseases. N. Engl. J. Med. 1987, 317, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleucin-1. In The Cytokine Handbook; Thompson, A.W., Ed.; Academic Press: San Diego, CA, USA, 1994; pp. 31–36. [Google Scholar]
- Wagner, J.G.; Roth, R.A. Neutrophil migration mechanisms, with an emphasis on the pulmonary vasculature. Pharmcol. Rev. 2000, 52, 349–374. [Google Scholar]
- Kumar, V.L.; Shivkar, Y.M. Involvement of prostaglandins in inflammation induced by latex of Calotropis procera. Mediat. Inflamm. 2004, 13, 151–155. [Google Scholar] [CrossRef]
- Yamashiro, S.; Kamohara, H.; Wang, J.M.; Yang, D.; Gong, W.H.; Yoshimura, T. Phenotypic and functional change of cytokine-activated neutrophils: Inflammatory neutrophils are heterogeneous and enhance adaptive immune responses. J. Leukoc. Biol. 2001, 69, 698–704. [Google Scholar] [PubMed]
- Sedgwick, A.D.; Lees, P. Studies of eicosanoid production in the air pouch model of synovial inflammation. Agents Actions 1986, 18, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Dal Secco, D.; Paron, J.A.; de Oliveira, S.H.P.; Ferreira, S.H.; Silva, J.S.; de Cunha, F.Q. Neutrophil migration in inflammation: Nitric oxide inhibits rolling, adhesion and induces apoptosis. Nitric Oxide Biol. Chem. 2003, 9, 153–164. [Google Scholar] [CrossRef]
- Ferrándiz, M.L.; Gil, B.; Sanz, M.J.; Ubeda, A.; Erazo, S.; González, E.; Negrete, R.; Pacheco, S.; Payá, M.; Alcaraz, M.J. Effect of bakuchiol on leukocyte functions and some inflammatory responses in mice. J. Pharm. Pharmcol. 1996, 48, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Saleh, T.S.; Calixto, J.B.; Medeiros, Y.S. Anti-inflammatory effects of theophylline, cromolyn and salbutamol in a murine model of pleurisy. Br. J. Pharmcol. 1996, 118, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Fröde, T.S.; Medeiros, Y.S. Myeloperoxidase and adenosine-deaminase levels in the pleural fluid leakage induced by carrageenan in the mouse model of pleurisy. Mediat. Inflamm. 2001, 10, 223–227. [Google Scholar] [CrossRef]
- da Silva, M.B.S.; Farges, R.C.; Fröde, T.S. Involvement of steroids in anti-inflammatory effects of PK11195 in a murine model of pleurisy. Mediat. Inflamm. 2004, 13, 93–103. [Google Scholar] [CrossRef]
- Koo, H.-J.; Lim, K.-H.; Jung, H.-J.; Park, E.-H. Anti-inflammatory evaluation of gardenia extract, geniposide and genipin. J. Ethnopharmacol. 2006, 103, 496–500. [Google Scholar] [CrossRef]
- Rau, B.M.; Krüger, C.M.; Hasel, C.; Oliveira, V.; Rubie, C.; Beger, H.G.; Schilling, M.K. Effects of immunosuppressive and immunostimulative treatment on pancreatic injury and mortality in severe acute experimental pancreatitis. Pancreas 2006, 33, 174–183. [Google Scholar] [CrossRef]
- Daxecker, H.; Raab, M.; Markovic, S.; Karimi, A.; Griesmacher, A.; Mueller, M.M. Endothelial adhesion molecule expression in an in vitro model of inflammation. Clin. Chim. Acta 2002, 325, 171–175. [Google Scholar] [CrossRef]
- Aller, M.-A.; Arias, J.-L.; Sánchez-Patán, F.; Arias, J. The inflammatory response: An efficient way of life. Med. Sci. Monit. 2006, 12, RA225–RA234. [Google Scholar] [PubMed]
- Schmid-Schönbein, G.W. Analysis of inflammation. Annu. Rev. Biomed. Eng. 2006, 8, 93–131. [Google Scholar] [CrossRef] [PubMed]
- Chichorro, J.G.; Lorenzetti, B.B.; Zampronio, A.R. Involvement of bradykinin, cytokines, sympathetic amines and prostaglandins in formalin-induced orofacial nociception in rats. Br. J. Pharmcol. 2004, 141, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Tassorelli, C.; Greco, R.; Wang, D.; Sandrini, G.; Nappi, G. Prostaglandins, glutamate and nitric oxide synthase mediate nitroglycerin-induced hyperalgesia in the formalin test. Eur. J. Pharmcol. 2006, 534, 103–107. [Google Scholar] [CrossRef]
- Farsky, S.H.P.; Borelli, P.; Fock, R.A.; Proto, S.Z.; Ferreira, J.M.C.; Mello, S.B. V Chronic blockade of nitric oxide biosynthesis in rats: Effect on leukocyte endothelial interaction and on leukocyte recruitment. Inflamm. Res. 2004, 53, 442–452. [Google Scholar] [CrossRef]
- Tuñón, M.J.; Sánchez-Campos, S.; Gutiérrez, B.; Culebras, J.M.; González-Gallego, J. Effects of FK506 and rapamycin on generation of reactive oxygen species, nitric oxide production and nuclear factor kappa B activation in rat hepatocytes. Biochem. Pharmcol. 2003, 66, 439–445. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Shirahase, H.; Kunishiro, K.; Ichikawa, A.; Kanda, M.; Uehara, Y. Synergistic effect of nitric oxide synthase and cyclooxygenase inhibitors on carrageenan-induced paw edema in rats. Arzneimittelforschung 2006, 56, 695–699. [Google Scholar] [CrossRef]
- Khaper, N.; Bryan, S.; Dhingra, S.; Singal, R.; Bajaj, A.; Pathak, C.M.; Singal, P.K. Targeting the vicious inflammation-oxidative stress cycle for the management of heart failure. Antioxid. Redox Signal. 2010, 13, 1033–1049. [Google Scholar] [CrossRef]
- Noble, E.G.; Shen, G.X. Impact of exercise and metabolic disorders on heat shock proteins and vascular inflammation. Autoimmune Dis. 2012, 2012, 836519. [Google Scholar] [CrossRef]
- Gordon, J.W.; Shaw, J.A.; Kirshenbaum, L.A. Multiple facets of NF-κB in the heart: To be or not to NF-κB. Circ. Res. 2011, 108, 1122–1132. [Google Scholar] [CrossRef]
- Fleetwood, A.J.; Lawrence, T.; Hamilton, J.A.; Cook, A.D. Granulocyte-macrophage colony-stimulating factor (CSF) and macrophage CSF-dependent macrophage phenotypes display differences in cytokine profiles and transcription factor activi-ties: Implications for CSF blockade in inflammation. J. Immunol. 2007, 178, 5245–5252. [Google Scholar] [CrossRef]
- Hsing, C.-H.; Lin, M.-C.; Choi, P.-C.; Huang, W.-C.; Kai, J.-I.; Tsai, C.-C.; Cheng, Y.-L.; Hsieh, C.-Y.; Wang, C.-Y.; Chang, Y.-P.; et al. Anesthetic propofol reduces endotoxic inflammation by inhibiting reactive oxygen species-regulated Akt/IKKβ/NF-κB signaling. PLoS ONE 2011, 6, e17598. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.-P.; Lai, Y.-S.; Wu, C.-J.; Chou, T.-C. Liquid perfluorochemical inhibits inducible nitric oxide synthase expression and nitric oxide formation in lipopolysaccharide-treated RAW 264.7 macrophages. J. Pharm. Sci. 2009, 111, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Effect of endocannabinoids on IgE-mediated allergic response in RBL-2H3 cells. Int. Immunopharmacol. 2013, 17, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Min, A.Y.; Doo, C.N.; Son, E.J.; Sung, N.Y.; Lee, K.J.; Sok, D.E.; Kim, M.R. N-palmitoyl serotonin alleviates scopolamine-induced memory impairment via regulation of cholinergic and antioxidant systems, and expression of BDNF and p-CREB in mice. Chem. Biol. Interact. 2015, 242, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Raymundo, L.J.R.P.; Guilhon, C.C.; Alviano, D.S.; Matheus, M.E.; Antoniolli, A.R.; Cavalcanti, S.C.H.; Alves, P.B.; Alviano, C.S.; Fernandes, P.D. Characterisation of the anti-inflammatory and antinociceptive activities of the Hyptis pectinata (L.) Poit essential oil. J. Ethnopharmacol. 2011, 134, 725–732. [Google Scholar] [CrossRef]
- Bartholomew, B. A rapid method for the assay of nitrate in urine using the nitrate reductase enzyme of Escherichia coli. Food Chem. Toxicol. 1984, 22, 541–543. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wisnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [5N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Cordeiro, N.M.; Freitas, R.H.C.N.; Fraga, C.A.M.; Fernandes, P.D. Discovery of Novel Orally Active Tetrahydro-Naphthyl-N-Acylhydrazones with In Vivo Anti-TNF-α Effect and Remarkable Anti-Inflammatory Properties. PLoS ONE 2016, 11, e0156271. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorno, T.B.S.; Lima, F.A.; Brand, A.L.M.; Oliveira, C.M.d.; Rezende, C.M.; Fernandes, P.D. Characterization of βN-Octadecanoyl-5-hydroxytryptamide Anti-Inflammatory Effect. Molecules 2021, 26, 3709. https://doi.org/10.3390/molecules26123709
Giorno TBS, Lima FA, Brand ALM, Oliveira CMd, Rezende CM, Fernandes PD. Characterization of βN-Octadecanoyl-5-hydroxytryptamide Anti-Inflammatory Effect. Molecules. 2021; 26(12):3709. https://doi.org/10.3390/molecules26123709
Chicago/Turabian StyleGiorno, Thais Biondino Sardella, Fernanda Alves Lima, Ana Laura Macedo Brand, Camila Martins de Oliveira, Claudia Moraes Rezende, and Patricia Dias Fernandes. 2021. "Characterization of βN-Octadecanoyl-5-hydroxytryptamide Anti-Inflammatory Effect" Molecules 26, no. 12: 3709. https://doi.org/10.3390/molecules26123709
APA StyleGiorno, T. B. S., Lima, F. A., Brand, A. L. M., Oliveira, C. M. d., Rezende, C. M., & Fernandes, P. D. (2021). Characterization of βN-Octadecanoyl-5-hydroxytryptamide Anti-Inflammatory Effect. Molecules, 26(12), 3709. https://doi.org/10.3390/molecules26123709