Binding Characteristics Study of DNA based Aptamers for E. coli O157:H7


Abstract
1. Introduction
2. Results and Discussion
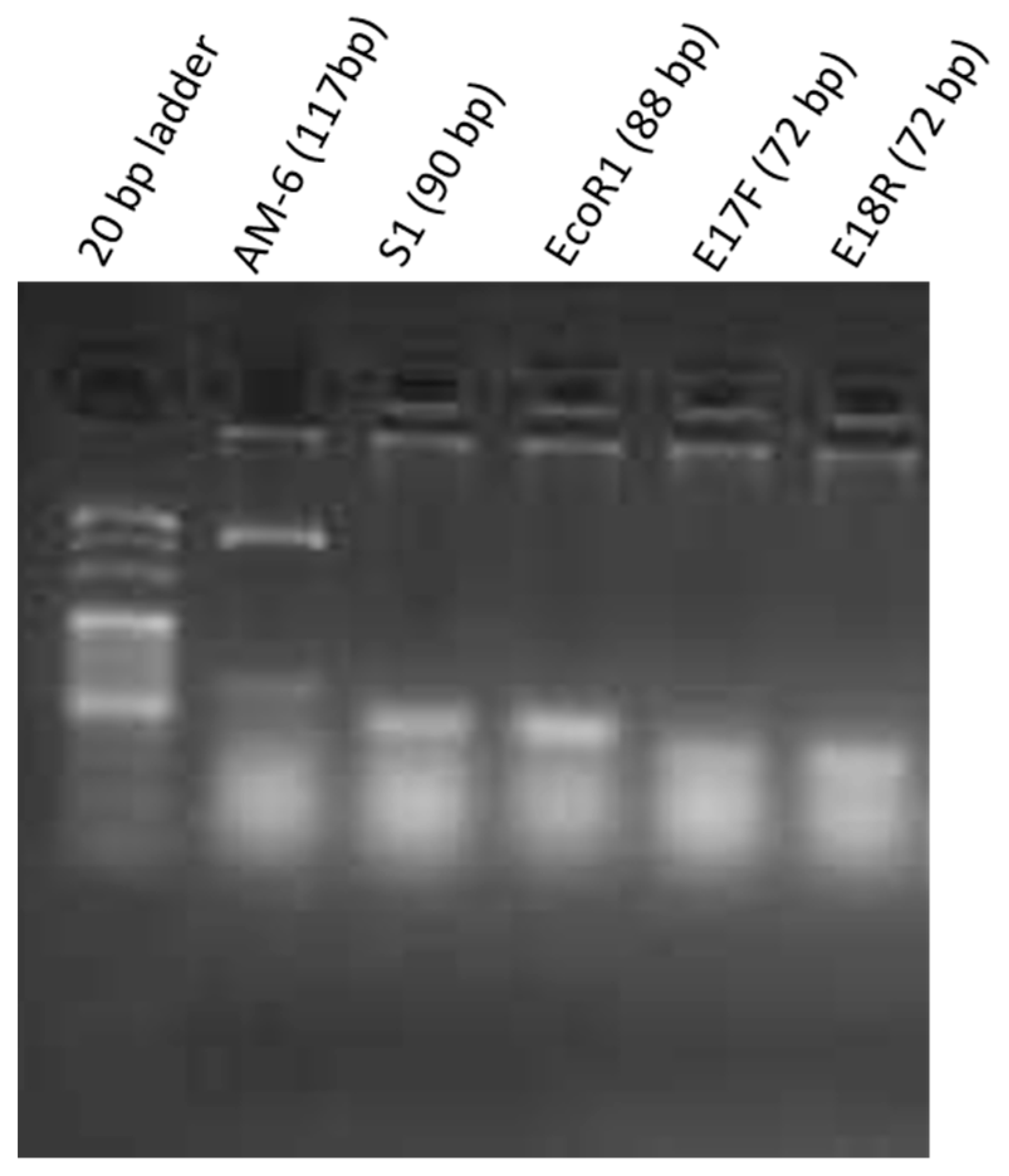
2.1. Qualitative Analysis
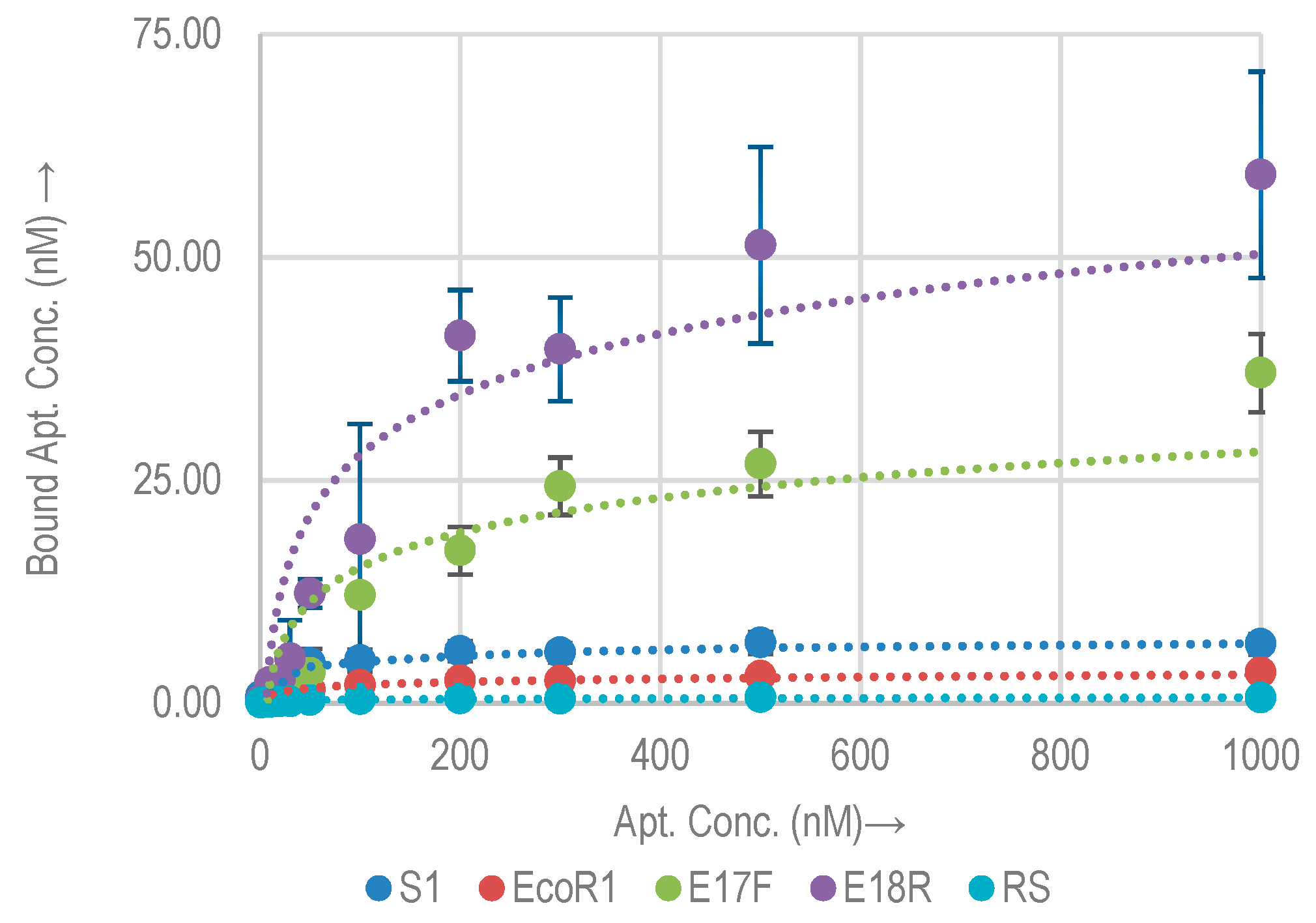
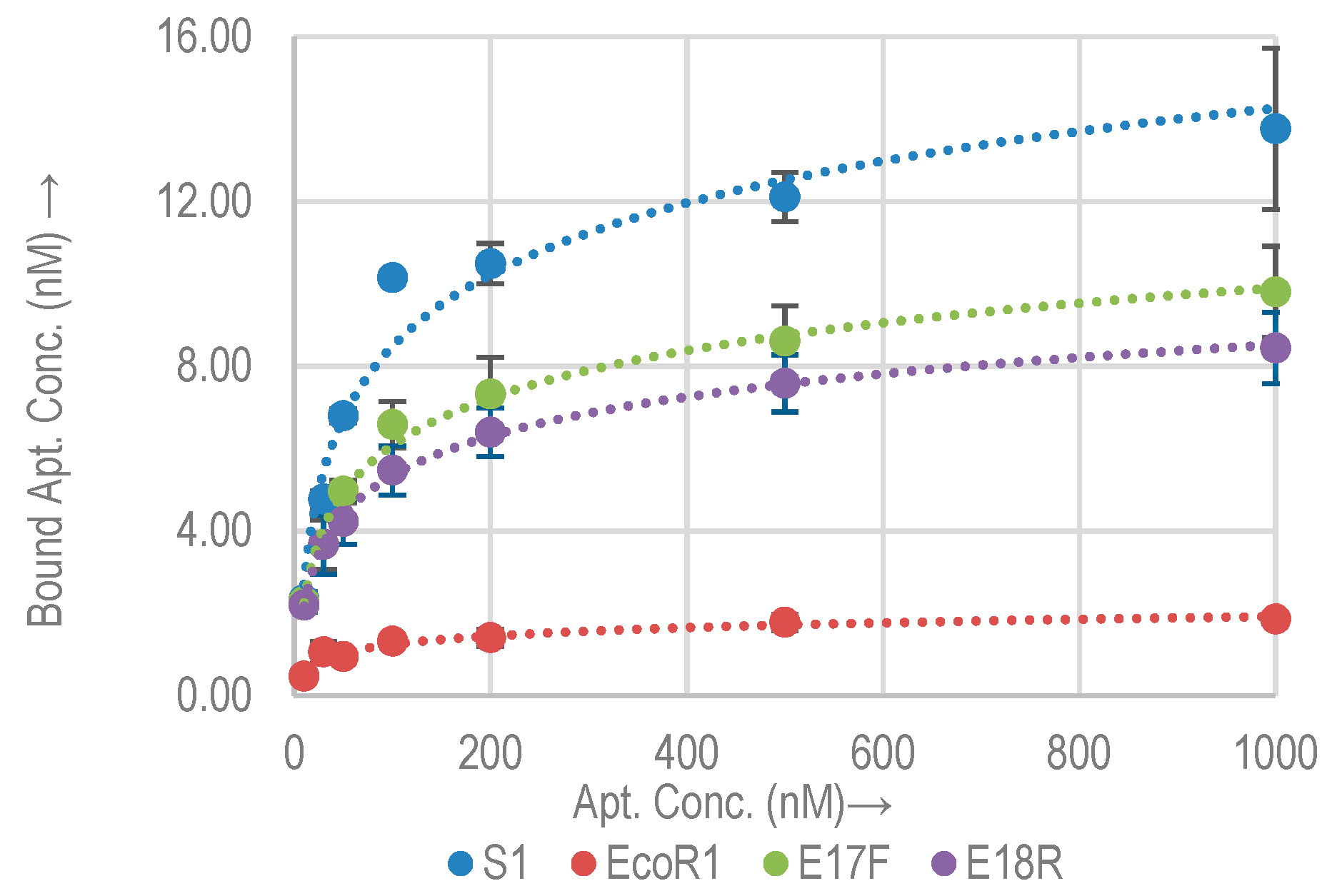
2.2. Quantitative Analysis (Kd Analysis by qPCR)
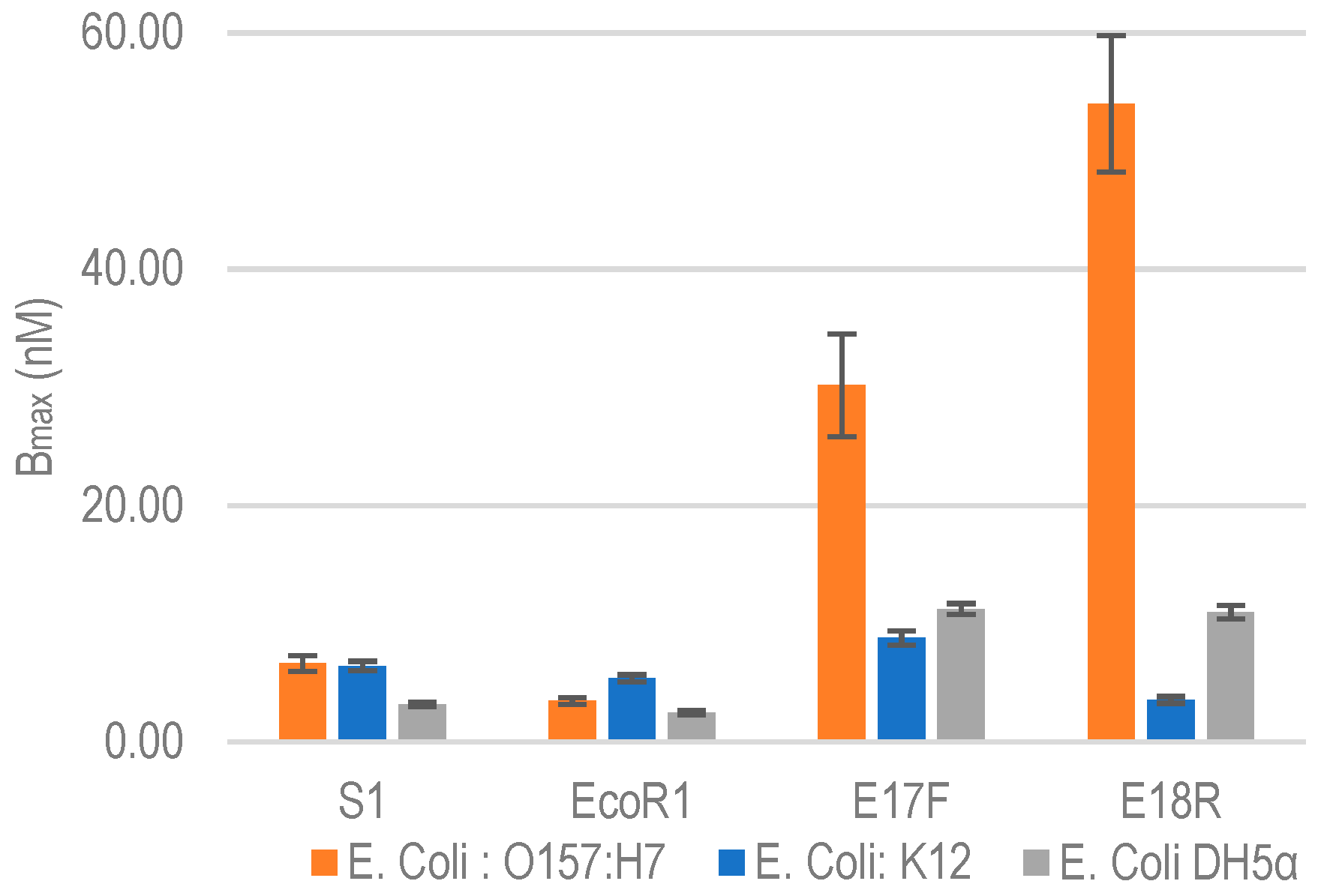
2.2.1. Quantitative Binding Analysis with E. coli O157:H7
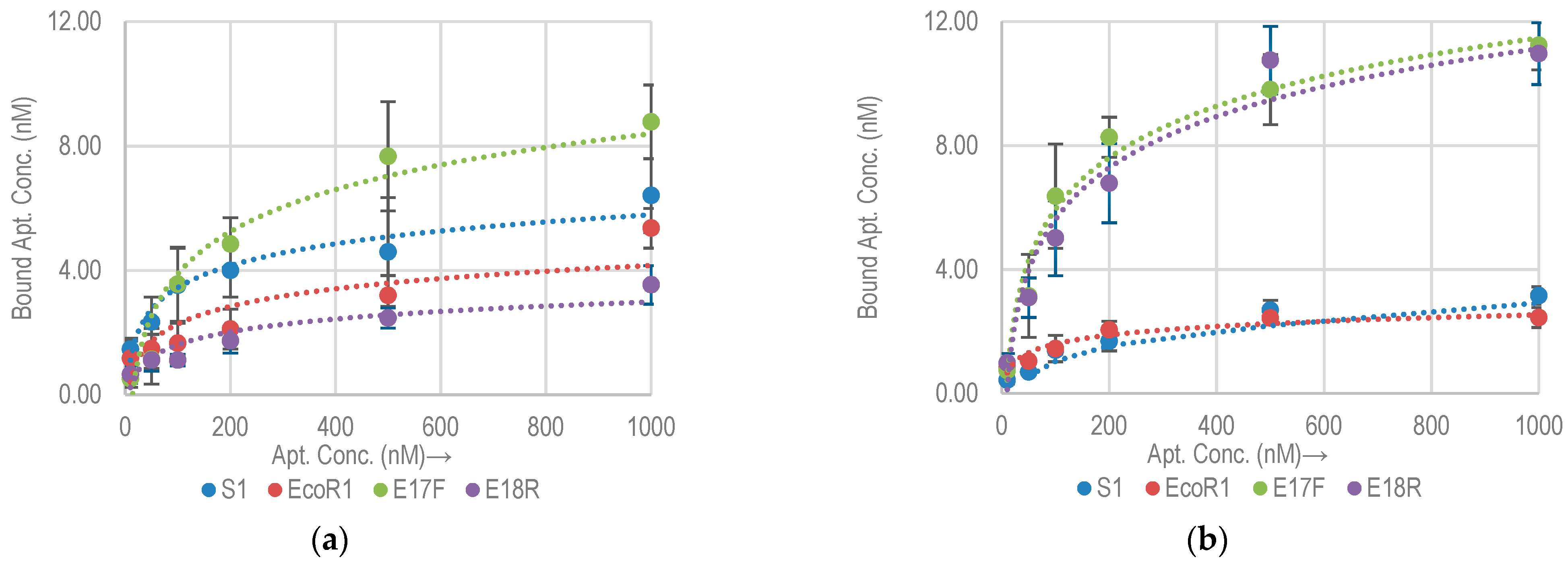
2.2.2. Non-Specific Aptamer Binding Analysis with E. coli K12 and DH5α
2.3. Effect of Biotin Tag on Aptamer Binding to E. coli O157:H7
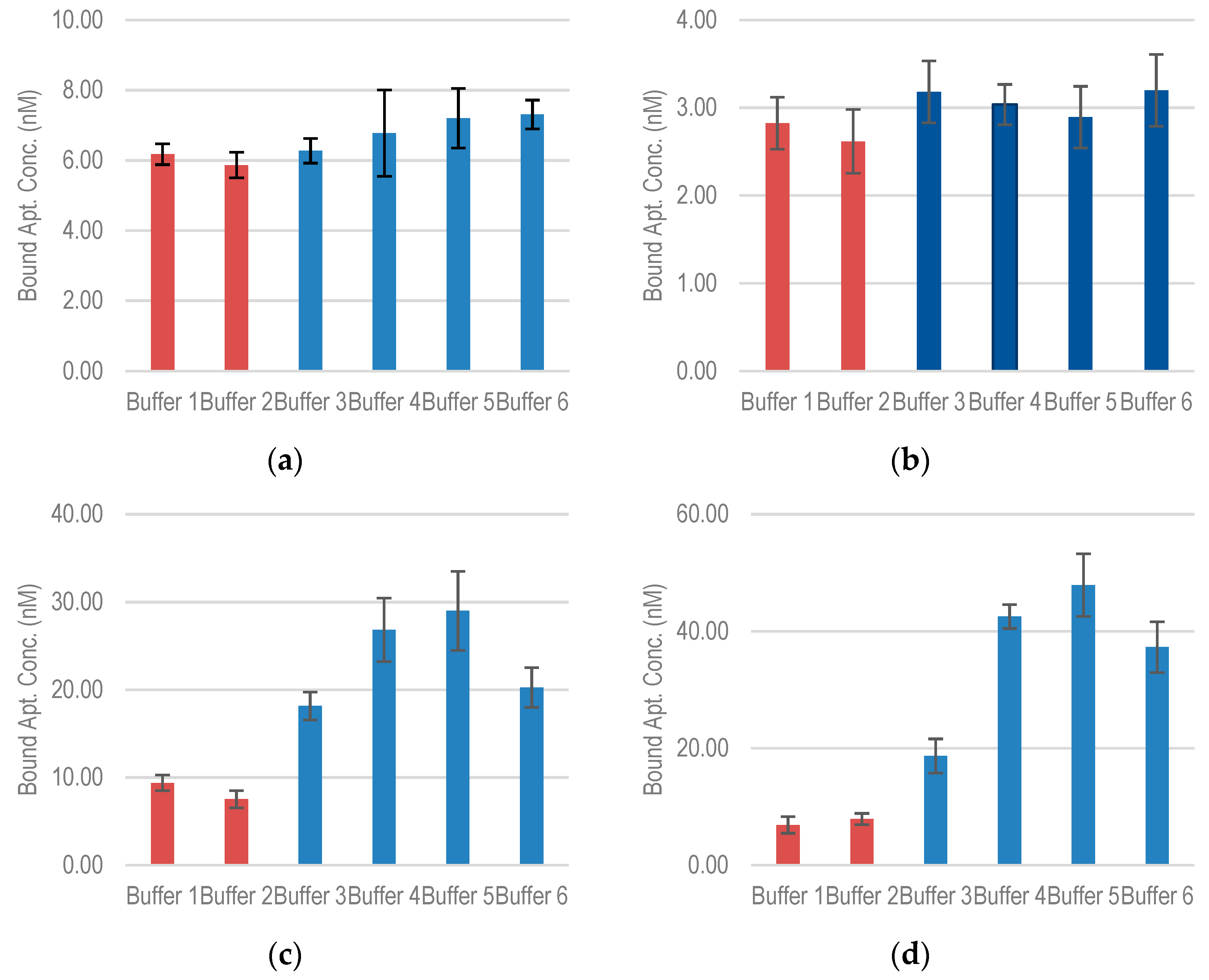
2.4. Effect of Ionic Strength on Aptamer-Target Binding
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. Aptamers and Primers
3.3. Aptamer–E. coli Binding Assay
3.4. Qualitative Aptamer-Target Binding Analysis
3.5. Quantitative Aptamer-Target Binding Analysis (Kd Analysis)
3.6. Specificity of Aptamers
3.7. Effect of Biotin-Tag Modification on the Binding Efficiency of Aptamers
3.8. Effect of Ionic Strength on the Aptamer-Target Binding
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorbach, S.L. Microbiology of the gastrointestinal tract. In Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Glaveston, TX, USA, 1996. [Google Scholar]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, V.; Nguyen, Y. Enterohemorrhagic E. coli (EHEC) pathogenesis. Front. Cell. Infect. Microbiol. 2012, 2, 90. [Google Scholar]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coliO157:H7 Outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Adams, M.C.; Naik, R.R.; Milam, V.T. Analyzing Secondary Structure Patterns in DNA Aptamers Identified via CompELS. Molecules 2019, 24, 1572. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- McKeague, M.; DeRosa, M.C. Challenges and Opportunities for Small Molecule Aptamer Development. J. Nucleic Acids 2012, 2012, 1–20. [Google Scholar] [CrossRef]
- Kong, H.Y.; Byun, J. Nucleic Acid Aptamers: New Methods for Selection, Stabilization, and Application in Biomedical Science. Biomol. Ther. 2013, 21, 423–434. [Google Scholar] [CrossRef]
- Turek, D.; Van Simaeys, D.; Johnson, J.; Ocsoy, I.; Tan, W. Molecular recognition of live methicillin-resistant staphylococcus aureus cells using DNA aptamers. World J. Transl. Med. 2013, 2, 67–74. [Google Scholar] [CrossRef]
- Dwivedi, H.P.; Smiley, R.D.; Jaykus, L.-A. Selection of DNA aptamers for capture and detection of Salmonella Typhimurium using a whole-cell SELEX approach in conjunction with cell sorting. Appl. Microbiol. Biotechnol. 2013, 97, 3677–3686. [Google Scholar] [CrossRef]
- Dwivedi, H.P.; Smiley, R.D.; Jaykus, L.-A. Selection and characterization of DNA aptamers with binding selectivity to Campylobacter jejuni using whole-cell SELEX. Appl. Microbiol. Biotechnol. 2010, 87, 2323–2334. [Google Scholar] [CrossRef] [PubMed]
- Amraee, M.; Oloomi, M.; Yavari, A.; Bouzari, S. DNA aptamer identification and characterization for E. coli O157 detection using cell based SELEX method. Anal. Biochem. 2017, 536, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chen, F.; Wang, R.; Li, Y. Whole-bacterium SELEX of DNA aptamers for rapid detection of E.coli O157:H7 using a QCM sensor. J. Biotechnol. 2018, 266, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Renuka, R.M.; Achuth, J.; Chandan, H.R.; Venkataramana, M.; Kadirvelu, K. A fluorescent dual aptasensor for the rapid and sensitive onsite detection of E. coli O157:H7 and its validation in various food matrices. New J. Chem. 2018, 42, 10807–10817. [Google Scholar] [CrossRef]
- Hein, P.; Michel, M.C.; Leineweber, K.; Wieland, T.; Wettschureck, N.; Offermanns, S. Receptor and Binding Studies. In Practical Methods in Cardiovascular Research; Springer: Berlin, Germany, 2006; pp. 723–783. [Google Scholar]
- Marton, S.; Cleto, F.; Krieger, M.A.; Cardoso, J. Isolation of an Aptamer that Binds Specifically to E. coli. PLoS ONE 2016, 11, e0153637. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Song, M.Y.; Jurng, J.; Kim, B.C. Isolation and characterization of DNA aptamers against Escherichia coli using a bacterial cell–systematic evolution of ligands by exponential enrichment approach. Anal. Biochem. 2013, 436, 22–28. [Google Scholar] [CrossRef]
- Moreno, M.; Fernandez-Algar, M.; Fernandez-Chamorro, J.; Ramajo, J.; Martinez-Salas, E.; Briones, C. A combined ELONA-(RT) qPCR approach for characterizing DNA and RNA aptamers selected against PCBP-2. Molecules 2019, 24, 1213. [Google Scholar] [CrossRef]
- Cho, E.J.; Lee, J.-W.; Ellington, A.D. Applications of Aptamers as Sensors. Annu. Rev. Anal. Chem. 2009, 2, 241–264. [Google Scholar] [CrossRef]
- Girardot, M.; Gareil, P.; Varenne, A. Interaction study of a lysozyme-binding aptamer with mono- and divalent cations by ACE. Electrophoresis 2010, 31, 546–555. [Google Scholar] [CrossRef]
- Noeske, J.; Buck, J.; Fürtig, B.; Nasiri, H.R.; Schwalbe, H.; Wöhnert, J. Interplay of ‘induced fit’ and preorganization in the ligand induced folding of the aptamer domain of the guanine binding riboswitch. Nucleic Acids Res. 2006, 35, 572–583. [Google Scholar] [CrossRef]
- Meyer, M. Applications of Aptamers in Flow Cytometry Assays; Gottfried Wilhelm Leibniz Universität Hannover: Hannover, Germany, 2014. [Google Scholar]
- Stoltenburg, R.; Schubert, T.; Strehlitz, B. In vitro Selection and Interaction Studies of a DNA Aptamer Targeting Protein A. PLoS ONE 2015, 10, e0134403. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.G.; Chanpong, J. Methods of Producing Competitive Aptamer Fret Reagents and Assays. US20090186342A1, 23 July 2009. [Google Scholar]
- Wu, W.; Zhang, J.; Zheng, M.; Zhong, Y.; Yang, J. An aptamer-based biosensor for colorimetric detection of Escherichia coli O157: H7. PLoS ONE 2012, 8, e004899. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zou, S.; Cao, X. A simple dendrimer-aptamer based microfluidic platform for E. coli O157:H7 detection and signal intensification by rolling circle amplification. Sensors Actuators B Chem. 2017, 251, 976–984. [Google Scholar] [CrossRef]
- Wu, W.; Zhao, S.; Mao, Y.; Fang, Z.; Lu, X.; Zeng, L. A sensitive lateral flow biosensor for Escherichia coli O157:H7 detection based on aptamer mediated strand displacement amplification. Anal. Chim. Acta 2015, 861, 62–68. [Google Scholar] [CrossRef]
- Pabinger, S.; Rödiger, S.; Kriegner, A.; Vierlinger, K.; Weinhäusel, A. A survey of tools for the analysis of quantitative PCR (qPCR) data. Biomol. Detect. Quantif. 2014, 1, 23–33. [Google Scholar] [CrossRef]
- Pedersen, J.; Lindup, W. Interpretation and analysis of receptor binding experiments which yield non-linear Scatchard plots and binding constants dependent upon receptor concentration. Biochem. Pharmacol. 1994, 47, 179–185. [Google Scholar] [CrossRef]
- Dolot, R.; Lam, C.H.; Sierant, M.; Zhao, Q.; Liu, F.-W.; Nawrot, B.; Egli, M.; Yang, X. Crystal structures of thrombin in complex with chemically modified thrombin DNA aptamers reveal the origins of enhanced affinity. Nucleic Acids Res. 2018, 46, 4819–4830. [Google Scholar] [CrossRef]
- Gemmill, D.; D’Souza, S.; Meier-Stephenson, V.; Patel, T.R. Current approaches for RNA-labelling to identify RNA-binding proteins. Biochem. Cell Biol. 2020, 98, 31–41. [Google Scholar] [CrossRef]
- Dua, P.; Ren, S.; Lee, S.W.; Kim, J.-K.; Shin, H.-S.; Jeong, O.-C.; Kim, S.; Lee, A.D.-K. Cell-SELEX Based Identification of an RNA Aptamer for Escherichia coli and Its Use in Various Detection Formats. Mol. Cells 2016, 39, 807–813. [Google Scholar] [CrossRef]
- Lipfert, J.; Sim, A.Y.; Herschlag, D.; Doniach, S. Dissecting electrostatic screening, specific ion binding, and ligand binding in an energetic model for glycine riboswitch folding. RNA 2010, 16, 708–719. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zavyalova, E.; Tagiltsev, G.; Reshetnikov, R.; Arutyunyan, A.; Kopylov, A. Cation Coordination Alters the Conformation of a Thrombin-Binding G-Quadruplex DNA Aptamer That Affects Inhibition of Thrombin. Nucleic Acid Ther. 2016, 26, 299–308. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| bp | Reported Kd | Sequence | Primers | |
|---|---|---|---|---|
| AM6 [13] | 117 | 107.6 ± 67.8 pmol | CGTGATGATGTTGAGTTGGGGTGATGGGTGCATGTGATGAAAGGGGTTCGTGCTATGCTGTTTTGTCTAATAATACTAGTCCTTGCCAAGGTTTATTCCAGTAATGCCAACCAATCT | FP- |
| CGTGATGATGTTGAGTTG RP- | ||||
| AGATTGGTTGGCATTACTG | ||||
| S1 [14] | 90 | 10.30 nM | CAGTCCAGGACAGATTCGCGAGTGGTCGTGGTGAGGTGCGTGTATGGGTGGTGGATGAGTGTGTGGCCACGTGGATTTCATTCAGCGATT | FP-CAGTCCAGGACAGATTCGCGAG |
| RP-AATCGCTGAATGAAATCCACGTG | ||||
| EcoR1 [15] | 88 | 41 ± 2 nM | ATTAGTCAAGAGGTAGACGCACATATCATCACAGCCGCAGCCGCCCCTTCCATTCACATGCCAGCTTCTGGTCGTCGTGACTCCTATA | FP-ATAGGAGTCACGACGACCAGAA RP-ATTAGTCAAGAGGTAGACGCACATA |
| E17F [25] | 72 | ATCCGTCACACCTGCTCTATCAAATGTGCAGATATCAAGACGATTTGTACAAGATGGTGTTGGCTCCCGTAT | FP- | |
| ATCCGTCACACCTGCTCT RP- | ||||
| ATACGGGAGCCAACACCATC | ||||
| E18R [25] | 72 | ATACGGGAGCCAACACCATTCTATCGTTCCGGACGCTTATGCCTTGCCATCTACAGAGCAGGTGTGACGGAT | FP- | |
| ATACGGGAGCCAACACCA RP- | ||||
| ATCCGTCACACCTGCTCT | ||||
| RS | 80 | ATCCAGAGTGACGCAGCATGCTTAAGGGGGGGGCGGGTTAAGGGAGTGGGGAGGGAGCTGGTGTGGACACGGTGGCTTAGT | FP- | |
| ATCCAGAGTGACGCAGCA RP- | ||||
| ACTAAGCCACCGTGTCCA |
| Aptamer | Kd (nM) | Bmax (nM) | Binding Sites Per Bacteria | Bmax Compared to RS |
|---|---|---|---|---|
| S1 | 25.7 ± 12 | 6.61 ± 1.1 | 1980 | 10× |
| EcoR1 | 45.5 ± 17 | 3.42 ± 0.5 | 1030 | 5× |
| E17F | 135 ± 72 | 30.17 ± 4.4 | 9050 | 47× |
| E18R | 151 ± 80 | 54.30 ± 10 | 16,290 | 85.64× |
| Aptamer | E. coli K12 | E. coli DH5α | ||||
|---|---|---|---|---|---|---|
| Kd (nM) | Bmax (nM) | Approx. Binding Sites/ Bacteria | Kd (nM) | Bmax (nM) | Approx. Binding Sites/ Bacteria | |
| S1 | 62.9 ± 27 | 6.41 ± 0.1 | 1920 | 114.2 ± 23 | 3.16 ± 0.1 | 940 |
| EcoR1 | 75.2 ± 28 | 5.36 ± 0.1 | 1600 | 48.3 ± 24 | 2.45 ± 0.1 | 730 |
| E17F | 202 ± 116 | 8.78 ± 0.6 | 2630 | 87.7 ± 14 | 11.2 ± 0.4 | 3360 |
| E18R | 111.3 ± 49 | 3.54 ± 0.2 | 1060 | 95.5 ± 24 | 11.0 ± 0.5 | 3300 |
| Aptamer | with Biotin | without Biotin | ||
|---|---|---|---|---|
| Kd (nM) | Bmax (nM) | Kd (nM) | Bmax (nM) | |
| S1 | 25.7 ± 12 | 13.76 ± 1.94 | 57.6 ± 10.6 | 6.61 ± 1.1 |
| EcoR1 | 45.5 ± 17 | 1.85 ± 0.03 | 33.7 ± 2.5 | 3.42 ± 0.5 |
| E17F | 135 ± 72 | 9.8 ± 0.9 | 51.2 ± 5.3 | 30.17 ± 4.4 |
| E18R | 151 ± 80 | 8.44 ± 0.5 | 45.5 ± 3.7 | 54.30 ± 10 |
| Binding Buffer without Dibasic Salt Ions |
|---|
| Buffer 1—50 mM Tris-HCl Ph 7.5, 50 mM NaCl. |
| Buffer 2—50 mM Tris-HCl pH 7.5, 50 mM NaCl, 5 mM KCl. |
| Binding Buffer with Dibasic Salt Ions |
| Buffer 3—50 mM Tris-HCl pH 7.5, 50 mM NaCl, 5 mM KCl, 0.5 mM MgCl2. |
| Buffer 4—50 mM Tris-HCl pH 7.5, 50 mM NaCl, 5 mM KCl, 1 mM MgCl2. |
| Buffer 5—50 mM Tris-HCl pH 7.5, 50 mM NaCl, 5 mM KCl, 5 mM MgCl2. |
| Buffer 6—50 mM Tris-HCl pH 7.5, 50 mM NaCl, 5 mM KCl, 10 mM MgCl2. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqui, S.; Yuan, J. Binding Characteristics Study of DNA based Aptamers for E. coli O157:H7. Molecules 2021, 26, 204. https://doi.org/10.3390/molecules26010204
Siddiqui S, Yuan J. Binding Characteristics Study of DNA based Aptamers for E. coli O157:H7. Molecules. 2021; 26(1):204. https://doi.org/10.3390/molecules26010204
Chicago/Turabian StyleSiddiqui, Saika, and Jie Yuan. 2021. "Binding Characteristics Study of DNA based Aptamers for E. coli O157:H7" Molecules 26, no. 1: 204. https://doi.org/10.3390/molecules26010204
APA StyleSiddiqui, S., & Yuan, J. (2021). Binding Characteristics Study of DNA based Aptamers for E. coli O157:H7. Molecules, 26(1), 204. https://doi.org/10.3390/molecules26010204