Profiling of Individual Desulfo-Glucosinolate Content in Cabbage Head (Brassica oleracea var. capitata) Germplasm

 ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Variation in Agronomic Characteristics
2.2. Identification and Quantification of Individual GSLs Profile in Cabbage
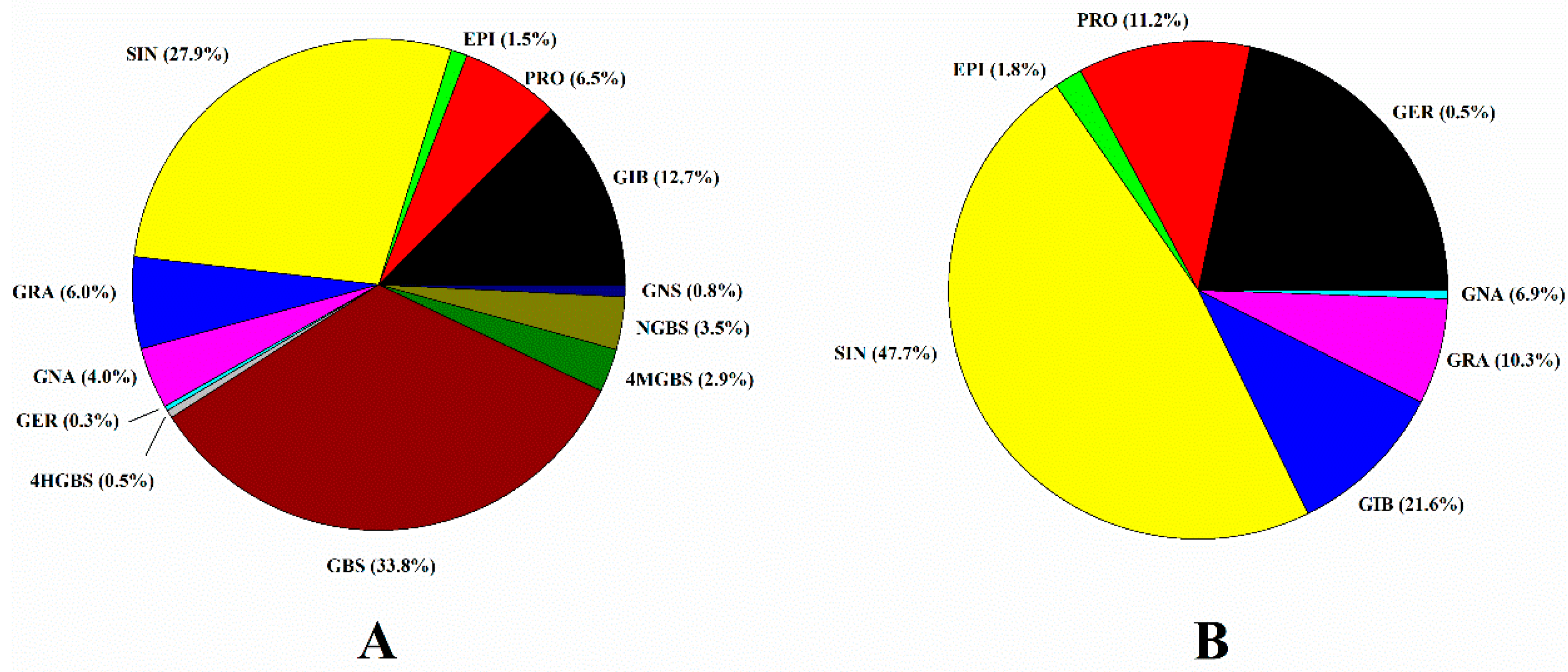
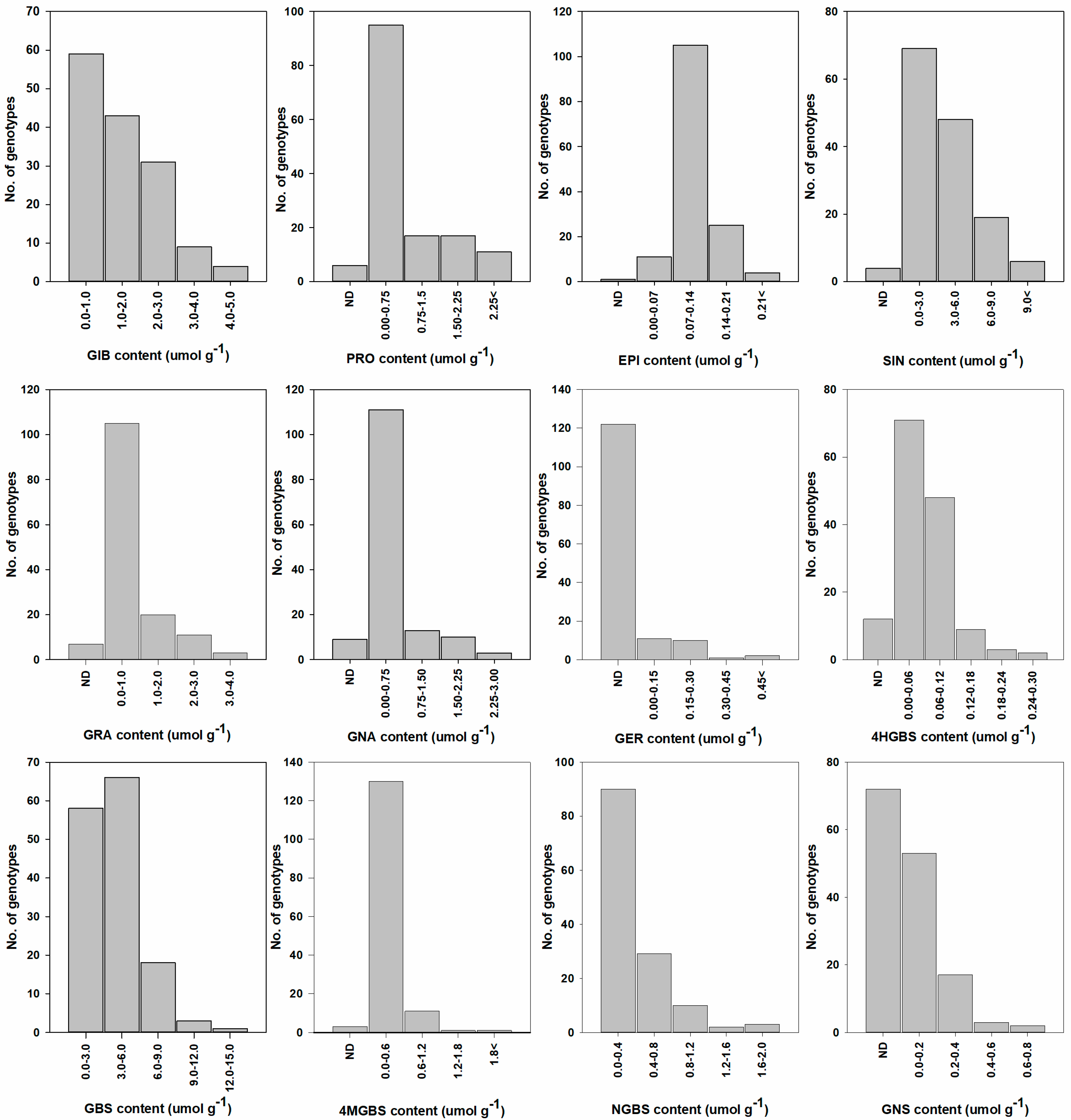
2.3. Variation in Individual and Total GSL Content among 146 Cabbage Genotypes
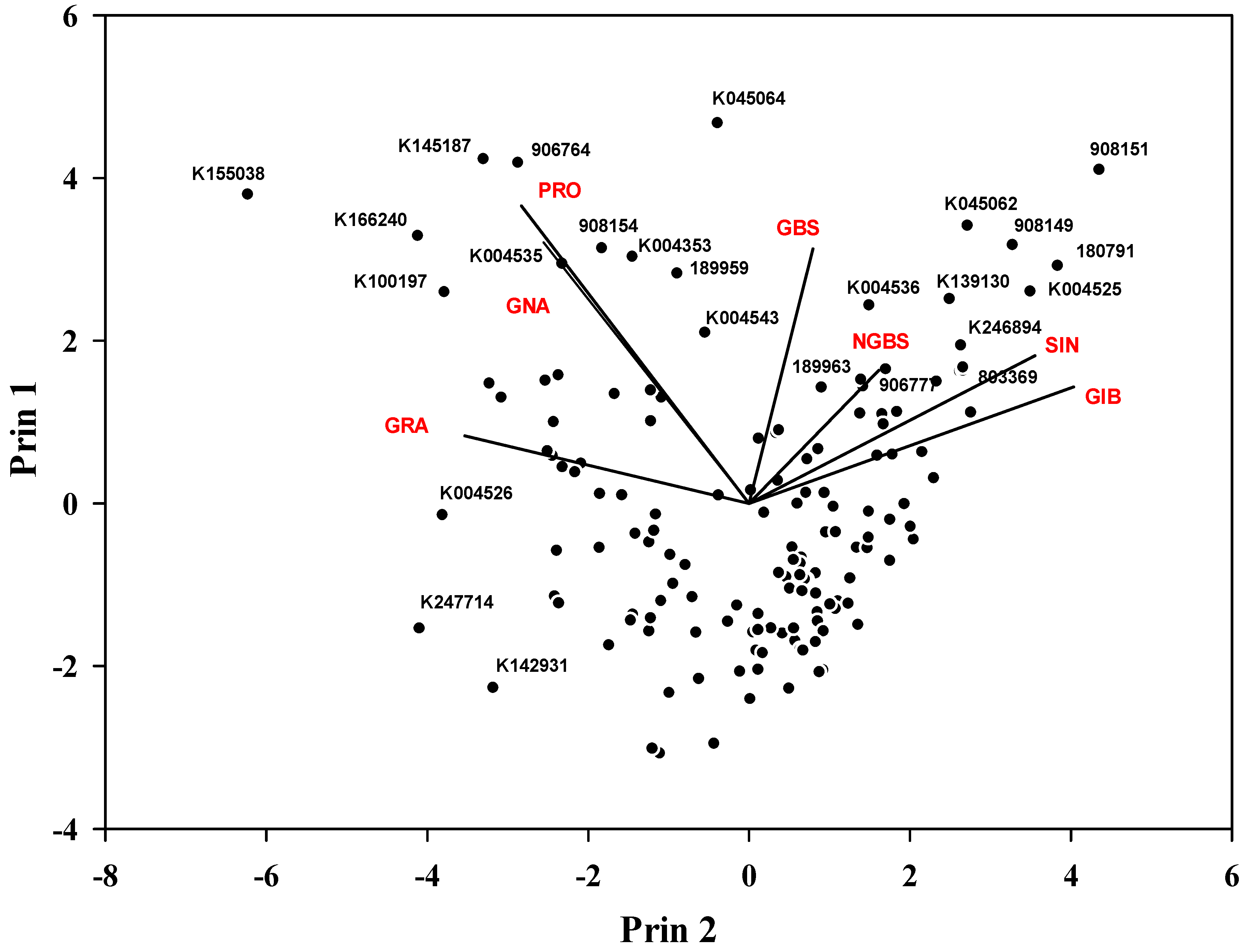
2.4. Selection of Candidate Genotypes for the Breeding Program
2.5. Correlation Analysis among the GSLs
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material and Cultivation
3.3. Extraction of Intact Glucosinolates (GSLs) and Their Desulfation
3.4. Separation and Identification of Individual GSLs Using High Performance Liquid Chromatography (HPLC)
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2019, 169, 112100. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant. Boil. 2006, 57, 303–333. [Google Scholar] [CrossRef]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310. [Google Scholar] [CrossRef]
- Sønderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of glucosinolates – gene discovery and beyond. Trends Plant. Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef]
- Agerbirk, N.; De Vos, M.; Kim, J.H.; Jander, G. Indole glucosinolate breakdown and its biological effects. Phytochem. Rev. 2008, 8, 101–120. [Google Scholar] [CrossRef]
- Agnihotri, A.R.; Hulagabali, C.V.; Adhav, A.S.; Joshi, R.S. Mechanistic insight in potential dual role of sinigrin against Helicoverpa armigera. Phytochemistry 2018, 145, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2007, 7, 213–229. [Google Scholar] [CrossRef]
- Pappa, G.; Lichtenberg, M.; Iori, R.; Barillari, J.; Bartsch, H.; Gerhauser, C. Comparison of growth inhibition profiles and mechanisms of apoptosis induction in human colon cancer cell lines by isothiocyanates and indoles from Brassicaceae. Mutat. Res. Mol. Mech. Mutagen. 2006, 599, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Traka, M.; Mithen, R. Glucosinolates, isothiocyanates and human health. Phytochem. Rev. 2008, 8, 269–282. [Google Scholar] [CrossRef]
- Avato, P.; Argentieri, M. Brassicaceae: A rich source of health improving phytochemicals. Phytochem. Rev. 2015, 14, 1019–1033. [Google Scholar] [CrossRef]
- Hong, E.; Kim, G.-H. Anticancer and antimicrobial activities of β-phenylethyl isothiocyanate in Brassica rapa L. Food Sci. Technol. Res. 2008, 14, 377–382. [Google Scholar] [CrossRef]
- James, D.; Devaraj, S.; Bellur, P.; Lakkanna, S.; Vicini, J.; Boddupalli, S. Novel concepts of broccoli sulforaphanes and disease: Induction of phase II antioxidant and detoxification enzymes by enhanced-glucoraphanin broccoli. Nutr. Rev. 2012, 70, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Staack, R.; Kingston, S.; Wallig, M.A.; Jeffery, E.H. A comparison of the individual and collective effects of four glucosinolate breakdown products from brussels sprouts on induction of detoxification enzymes. Toxicol. Appl. Pharmacol. 1998, 149, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Bommareddy, A.; Hahm, E.-R.; Xiao, N.; Powolny, A.A.; Fisher, A.; Jiang, Y.; Singh, S.V. Atg5 regulates phenethyl isothiocyanate-induced autophagic and apoptotic cell death in human prostate cancer cells. Cancer Res. 2009, 69, 3704–3712. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.I.; Kirkegaard, J.; Wong, P.; Desmarchelier, J. Biofumigation potential of brassicas. Plant. Soil 1998, 201, 103–112. [Google Scholar] [CrossRef]
- Avato, P.; D’Addabbo, T.; Leonetti, P.; Argentieri, M. Nematicidal potential of Brassicaceae. Phytochem. Rev. 2013, 12, 791–802. [Google Scholar] [CrossRef]
- Baik, H.-Y.; Juvik, J.; Jeffery, E.H.; Wallig, M.A.; Kushad, M.; Klein, B. Relating glucosinolate content and flavor of broccoli cultivars. J. Food Sci. 2003, 68, 1043–1050. [Google Scholar] [CrossRef]
- Choi, S.-H.; Park, S.; Lim, Y.P.; Kim, J.B.; Park, J.-T.; An, G. Metabolite profiles of glucosinolates in cabbage varieties (Brassica oleracea var. capitata) by season, color, and tissue position. Hortic. Environ. Biotechnol. 2014, 55, 237–247. [Google Scholar] [CrossRef]
- Park, S.; Arasu, M.V.; Lee, M.-K.; Chun, J.-H.; Seo, J.M.; Lee, S.-W.; Al-Dhabi, N.A.; Kim, J.B. Quantification of glucosinolates, anthocyanins, free amino acids, and vitamin C in inbred lines of cabbage (Brassica oleracea L.). Food Chem. 2014, 145, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Arasu, M.V.; Lee, M.-K.; Chun, J.-H.; Seo, J.M.; Al-Dhabi, N.A.; Kim, J.B. Analysis and metabolite profiling of glucosinolates, anthocyanins and free amino acids in inbred lines of green and red cabbage (Brassica oleracea L.). LWT-Food Sci. Technol. 2014, 58, 203–213. [Google Scholar] [CrossRef]
- Šamec, D.; Pavlović, I.; Sondi, B.S. White cabbage (Brassica oleracea var. capitata f. alba): Botanical, phytochemical and pharmacological overview. Phytochem. Rev. 2016, 16, 117–135. [Google Scholar] [CrossRef]
- Vale, A.; Santos, J.; Brito, A.; Fernandes, D.; Rosa, E.; Oliveira, M.B.P.P. Evaluating the impact of sprouting conditions on the glucosinolate content of Brassica oleracea sprouts. Phytochemistry 2015, 115, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yue, Z.; Zhong, X.; Lei, J.; Tao, P.; Li, B. Distribution of primary and secondary metabolites among the leaf layers of headed cabbage (Brassica oleracea var. capitata). Food Chem. 2019, 312, 126028. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Jo, J.S.; Lee, J.G. Comparison of glucosinolate profiles in different tissues of nine Brassica crops. Molecules 2015, 20, 15827–15841. [Google Scholar] [CrossRef] [PubMed]
- Fritz, V.A.; Justen, V.L.; Bode, A.M.; Schuster, T.; Wang, M. Glucosinolate enhancement in cabbage induced by jasmonic acid application. HortScience 2010, 45, 1188–1191. [Google Scholar] [CrossRef]
- Kołodziejski, D.; Piekarska, A.; Hanschen, F.S.; Pilipczuk, T.; Tietz, F.; Kusznierewicz, B.; Bartoszek, A. Relationship between conversion rate of glucosinolates to isothiocyanates/indoles and genotoxicity of individual parts of Brassica vegetables. Eur. Food Res. Technol. 2018, 245, 383–400. [Google Scholar] [CrossRef]
- Wang, J.; Gu, H.; Yu, H.; Zhao, Z.; Sheng, X.; Zhang, X. Genotypic variation of glucosinolates in broccoli (Brassica oleracea var. italica) florets from China. Food Chem. 2012, 133, 735–741. [Google Scholar] [CrossRef]
- Yi, G.; Lim, S.; Chae, W.B.; Park, J.E.; Park, H.R.; Lee, E.J.; Huh, J.H. Root glucosinolate profiles for screening of radish (Raphanus sativus L.) genetic resources. J. Agric. Food Chem. 2015, 64, 61–70. [Google Scholar] [CrossRef]
- Lee, M.-K.; Chun, J.-H.; Byeon, D.H.; Chung, S.O.; Park, S.; Park, S.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, Y.-P.; Kim, J.B. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp. pekinensis) and their antioxidant activity. LWT-Food Sci. Technol. 2014, 58, 93–101. [Google Scholar] [CrossRef]
- Hasan, M.; Friedt, W.; Pons-Kühnemann, J.; Freitag, N.M.; Link, K.; Snowdon, R.J. Association of gene-linked SSR markers to seed glucosinolate content in oilseed rape (Brassica napus ssp. napus). Theor. Appl. Genet. 2008, 116, 1035–1049. [Google Scholar] [CrossRef] [PubMed]
- Klopsch, R.; Witzel, K.; Börner, A.; Schreiner, M.; Hanschen, F.S. Metabolic profiling of glucosinolates and their hydrolysis products in a germplasm collection of Brassica rapa turnips. Food Res. Int. 2017, 100, 392–403. [Google Scholar] [CrossRef]
- Lee, J.G.; Bonnema, G.; Zhang, N.; Kwak, J.H.; De Vos, R.C.H.; Beekwilder, J. Evaluation of glucosinolate variation in a collection of turnip (Brassica rapa) germplasm by the analysis of intact and desulfo glucosinolates. J. Agric. Food Chem. 2013, 61, 3984–3993. [Google Scholar] [CrossRef] [PubMed]
- Wiesner-Reinhold, M.; Zrenner, R.; Krumbein, A.; Glatt, H.; Schreiner, M. Genotypic variation of the glucosinolate profile in pak choi (Brassica rapa ssp. chinensis). J. Agric. Food Chem. 2013, 61, 1943–1953. [Google Scholar] [CrossRef]
- Radovich, T.J.; Kleinhenz, M.D.; Streeter, J.G.; Miller, A.R.; Scheerens, J.C. Planting date affects total glucosinolate concentrations in six commercial cabbage cultivars. HortScience 2005, 40, 106–110. [Google Scholar] [CrossRef]
- Sarikamis, G.; Balkaya, A.; Yanmaz, R. Glucosinolates within a collection of white head cabbages (Brassica oleracea var. capitata sub. var. alba) from Turkey. Afr. J. Biotechnol. 2009, 8, 5046–5052. [Google Scholar]
- Banerjee, A.; Variyar, P.S.; Chatterjee, S.; Sharma, A. Effect of post harvest radiation processing and storage on the volatile oil composition and glucosinolate profile of cabbage. Food Chem. 2014, 151, 22–30. [Google Scholar] [CrossRef]
- Ciska, E.; Drabińska, N.; Narwojsz, A.; Honke, J. Stability of glucosinolates and glucosinolate degradation products during storage of boiled white cabbage. Food Chem. 2016, 203, 340–347. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P.; Obregón, S.; Padilla, G.; De Haro, A. Seasonal variation in glucosinolate content in Brassica oleracea crops grown in northwestern Spain. Phytochemistry 2008, 69, 403–410. [Google Scholar] [CrossRef]
- Pocock, K.; Heaney, R.K.; Wilkinson, A.P.; Beaumont, J.E.; Vaughan, J.G.; Fenwick, G.R. Changes in myrosinase activity and isoenzyme pattern, glucosinolate content and the cytology of myrosin cells in the leaves of heads of three cultivars of English white cabbage. J. Sci. Food Agric. 1987, 41, 245–257. [Google Scholar] [CrossRef]
- Awasthi, S.; Saraswathi, N. Sinigrin, a major glucosinolate from cruciferous vegetables restrains non-enzymatic glycation of albumin. Int. J. Boil. Macromol. 2016, 83, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Orlikova, B.; Diederich, M. Power from the garden: Plant compounds as inhibitors of the hallmarks of cancer. Curr. Med. Chem. 2012, 19, 2061–2087. [Google Scholar] [CrossRef] [PubMed]
- Vicas, S.; Teusdea, A.C.; Carbunar, M.; Socaci, S.A.; Socaciu, C. Glucosinolates profile and antioxidant capacity of Romanian Brassica vegetables obtained by organic and conventional agricultural practices. Plant. Foods Hum. Nutr. 2013, 68, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Washida, K.; Miyata, M.; Koyama, T.; Yazawa, K.; Nomoto, K. Suppressive effect of yamato-mana (Brassica rapa L. oleifera group) constituent 3-butenyl glucosinolate (gluconapin) on postprandial hypertriglyceridemia in mice. Biosci. Biotechnol. Biochem. 2010, 74, 1286–1289. [Google Scholar] [CrossRef]
- Conaway, C.C.; Yang, Y.-M.; Chung, F.-L. Isothiocyanates as cancer chemopreventive agents: Their biological activities and metabolism in rodents and humans. Curr. Drug Metab. 2002, 3, 233–255. [Google Scholar] [CrossRef]
- Lee, H.-W.; Lee, C.G.; Rhee, D.-K.; Um, S.H.; Pyo, S.; Pyo, S. Sinigrin inhibits production of inflammatory mediators by suppressing NF-κB/MAPK pathways or NLRP3 inflammasome activation in macrophages. Int. Immunopharmacol. 2017, 45, 163–173. [Google Scholar] [CrossRef]
- Zhang, N.-Q.; Ho, S.C.; Mo, X.-F.; Lin, F.-Y.; Huang, W.; Luo, H.; Huang, J.; Zhang, C.-X. Glucosinolate and isothiocyanate intakes are inversely associated with breast cancer risk: A case–control study in China. Br. J. Nutr. 2018, 119, 957–964. [Google Scholar] [CrossRef]
- Pocasap, P.; Weerapreeyakul, N. Sulforaphene and sulforaphane in commonly consumed cruciferous plants contributed to antiproliferation in HCT116 colon cancer cells. Asian Pac. J. Trop. Biomed. 2016, 6, 119–124. [Google Scholar] [CrossRef]
- Tripathi, M.; Mishra, A. Glucosinolates in animal nutrition: A review. Anim. Feed. Sci. Technol. 2007, 132, 1–27. [Google Scholar] [CrossRef]
- Yi, G.-E.; Robin, A.H.K.; Yang, K.; Park, J.-I.; Hwang, B.H.; Nou, I.-S. Exogenous methyl jasmonate and salicylic acid induce subspecies-specific patterns of glucosinolate accumulation and gene expression in Brassica oleracea L. Molecules 2016, 21, 1417. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Hara, M.; Fukino, N.; Kakizaki, T.; Morimitsu, Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed. Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of each cabbage germplasm is available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SN | Retention Time (min) | Common Name | Semisystematic Name of R-Group | Abbreviation | Group | Linearity Curve | R2 |
|---|---|---|---|---|---|---|---|
| 1 | 3.51 | Glucoiberin | 3-Methylsulfinylpropyl | GIB | Aliphatic | y = 30.468x + 0.236 | 0.9999 |
| 2 | 4.46 | Glucolepidiin | Ethyl | GLP | Aliphatic | y = 38.502x − 0.447 | 0.9999 |
| 3 | 4.95 | Progoitrin | 2-Hydroxy-3-butenyl | PRO | Aliphatic | y = 35.060x − 0.647 | 0.9999 |
| 4 | 6.11 | Epiprogoitrin | (2R)-2-Hydroxy-3-butenyl | EPI | Aliphatic | y = 18.618x − 0.537 | 0.9999 |
| 5 | 6.81 | Sinigrin | 2-Propenyl | SIN | Aliphatic | y = 43.343x − 0.236 | 0.9999 |
| 6 | 7.15 | Glucoraphanin | 4-Methylsulfinylbutyl | GRA | Aliphatic | y = 31.056x + 2.932 | 0.9992 |
| 7 | 8.06 | Glucoraphenin | 4-(Methylthio)butyl | GRE | Aliphatic | y = 7.5889x − 2.068 | 0.9989 |
| 8 | 10.62 | Sinalbin | 4-hydroxybenzyl | SNB | Aromatic | y = 132.13x − 6.977 | 0.9999 |
| 9 | 11.78 | Gluconapin | 3-Butenyl | GNA | Aliphatic | y = 30.566x − 0.487 | 0.9999 |
| 10 | 12.34 | 4-Hydroxyglucobrassicin | 4-Hydroxy-3-indolylmethyl | 4HGBS | Indolyl | y = 53.070x − 1.559 | 0.9996 |
| 11 | 12.67 | Glucomoringin | 4-(α-l-rhamnopyranosyloxy)benzyl | GMR | Aromatic | y = 9.494x − 0.769 | 0.9990 |
| 12 | 15.01 | Glucobarbarin | 2-Hydroxy-2-phenylethyl | GBA | Aliphatic | y = 35.957x + 0.660 | 0.9997 |
| 13 | 15.89 | Glucobrassicanapin | 4-Pentenyl | GBN | Aliphatic | y = 34.109x − 0.258 | 0.9999 |
| 14 | 16.11 | Glucotropaeolin | Benzyl | GTR | Aromatic | y = 34.081x + 0.067 | 0.9999 |
| 15 | 16.22 | Glucoerucin | 4-(Methylthio)butyl | GER | Aliphatic | y = 29.598x − 5.741 | 0.9994 |
| 16 | 16.67 | Glucobrassicin | 3-Indolylmethyl | GBS | Indolyl | y = 67.591x + 0.541 | 0.9997 |
| 17 | 17.24 | 4-Methoxyglucobrassicin | 4-Methoxy-3-indolylmethyl | 4MGBS | Indolyl | y = 81.427x +1.562 | 0.9996 |
| 18 | 17.42 | Gluconasturtiin | 2-Penylethyl | GNS | Aromatic | y = 20.730x − 0.538 | 0.9999 |
| 19 | 18.56 | Neoglucobrassicin | N-Methoxy-3-indolylmethyl | GNBS | Indolyl | y = 90.766x − 1.228 | 0.9997 |
| GSLs a | Average (μmol g−1) | Range (μmol g−1) | CV (%) b |
|---|---|---|---|
| GIB | 1.50 | 0.01–5.01 | 69.93 |
| PRO | 0.77 | 0.00–3.86 | 106.57 |
| EPI | 0.11 | 0.00–0.26 | 36.13 |
| SIN | 3.45 | 0.00–12.87 | 79.15 |
| GRA | 0.62 | 0.00–3.85 | 129.88 |
| GNA | 0.47 | 0.00–2.81 | 126.77 |
| GER | 0.03 | 0.00–0.61 | 280.47 |
| Total Aliphatic GSLs | 6.95 | 1.31–17.88 | 46.39 |
| 4HGBS | 0.06 | 0.00–0.29 | 85.47 |
| GBS | 3.91 | 0.79–1.14 | 53.53 |
| 4MGBS | 0.29 | 0.00–2.57 | 94.99 |
| NGBS | 0.40 | 0.05–2.00 | 84.33 |
| Total Indolyl GSLs | 4.46 | 1.01–15.57 | 51.16 |
| GNS | 0.09 | 0.00–0.67 | 137.77 |
| Total Aromatic GSLs | 0.09 | 0.00–0.67 | 137.77 |
| Total | 11.71 | 3.99–23.75 | 36.44 |
| IT No. | Temporary No. | Name | aGlucosinolate Content (μmol g−1 DW) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GIB | PRO | EPI | SIN | GRA | GNA | 4HGBS | GER | GBS | 4MGBS | GNS | NGBS | Total GSLs | |||
| bNA | 908151 | BOL-AWS-1999-156 | 3.86 | 0.20 | 0.11 | 3.46 | 0.20 | 0.16 | 0.03 | cND | 13.14 | 0.21 | 0.67 | 1.73 | 23.76 |
| NA | 908149 | BOL-AWS-1999-153 | 2.92 | 1.17 | 0.12 | 12.87 | 0.20 | 0.61 | 0.11 | ND | 4.09 | 0.26 | 0.29 | 0.74 | 23.37 |
| NA | K045064 | Valcatiecskaya | 1.22 | 2.06 | 0.21 | 10.74 | 0.30 | 1.93 | 0.04 | 0.15 | 5.88 | 0.20 | ND | 0.58 | 23.31 |
| 180791 | NA | Late Flat Dutch | 4.87 | 0.49 | 0.09 | 5.77 | 0.31 | 0.23 | 0.11 | ND | 9.08 | 0.27 | 0.12 | 1.26 | 22.59 |
| NA | K045062 | Kashirka 202 | 3.16 | 0.90 | 0.12 | 6.88 | 0.54 | 0.46 | 0.28 | ND | 7.97 | 0.29 | 0.12 | 0.90 | 21.62 |
| NA | K246894 | Succession Green Leaved | 2.83 | 0.25 | 0.14 | 9.53 | 0.09 | 0.05 | 0.06 | ND | 7.48 | 0.40 | ND | 0.49 | 21.33 |
| NA | K139130 | Sagyahwak | 3.39 | 0.89 | 0.15 | 10.05 | 0.26 | 0.49 | 0.09 | ND | 4.90 | 0.17 | 0.24 | 0.10 | 20.72 |
| NA | 907279 | Yujanka 31 | 2.79 | 1.03 | 0.09 | 11.29 | 0.20 | 0.53 | 0.06 | ND | 3.25 | 0.10 | 0.24 | 0.28 | 19.86 |
| NA | K004525 | Podarok | 5.01 | 0.53 | 0.11 | 5.67 | 0.35 | 0.30 | 0.29 | ND | 5.87 | 0.37 | 0.16 | 0.81 | 19.49 |
| NA | 803369 | Zuun kharaa No 10 | 2.35 | 0.52 | 0.10 | 10.29 | 0.13 | 0.56 | 0.04 | ND | 4.38 | 0.17 | 0.28 | 0.56 | 19.39 |
| NA | 803374 | Zuun kharaa No 15 | 4.55 | 0.50 | 0.10 | 8.28 | 0.21 | 0.32 | 0.04 | ND | 4.51 | 0.39 | ND | 0.29 | 19.19 |
| NA | K145187 | Kirmizi | 0.57 | 3.41 | 0.15 | 2.00 | 1.53 | 2.79 | ND | 0.16 | 6.40 | 0.22 | ND | 0.95 | 18.18 |
| NA | 906764 | Natsuzoka | 1.08 | 3.28 | 0.22 | 2.17 | 1.47 | 1.60 | 0.08 | 0.12 | 7.46 | 0.13 | ND | 0.22 | 17.82 |
| NA | 906777 | UR Gogetsu | 2.11 | 0.44 | 0.11 | 2.81 | 0.29 | 0.23 | 0.02 | ND | 10.60 | 0.47 | 0.26 | 0.33 | 17.66 |
| NA | K004353 | Rubin | 1.00 | 3.28 | 0.17 | 2.56 | 2.04 | ND | 0.15 | ND | 7.76 | 0.19 | ND | 0.41 | 17.55 |
| 160677 | NA | Gyeongphong 1 ho | 3.15 | 0.86 | 0.10 | 6.33 | 0.31 | 0.25 | 0.17 | ND | 4.50 | 0.74 | 0.62 | 0.15 | 17.16 |
| 189963 | NA | Skvirskaya N32 | 2.02 | 0.45 | 0.14 | 2.98 | 0.22 | 0.22 | 0.10 | ND | 10.10 | 0.39 | ND | 0.19 | 16.81 |
| NA | K166205 | 153 | 1.84 | 0.08 | 0.09 | 4.11 | 0.06 | 0.08 | 0.02 | ND | 6.34 | 0.22 | ND | 1.96 | 14.78 |
| NA | K155038 | Tashkent 110 | 0.05 | 3.86 | 0.26 | 0.09 | 2.65 | 0.92 | 0.04 | 0.56 | 5.47 | 0.46 | ND | 0.36 | 14.71 |
| NA | K166240 | Red Drumhead 2 | 0.65 | 3.06 | 0.19 | 1.49 | 1.78 | 2.36 | 0.06 | 0.22 | 4.42 | ND | ND | 0.16 | 14.41 |
| NA | 803360 | Zuun kharaa No 1 | 4.06 | 0.20 | 0.08 | 4.48 | 0.11 | 0.13 | 0.03 | ND | 4.03 | 0.17 | ND | 0.25 | 13.54 |
| NA | K100197 | Pourovo cervene | 0.31 | 2.49 | 0.18 | 0.77 | 1.47 | 2.81 | 0.07 | 0.10 | 3.26 | 0.16 | ND | 0.42 | 12.03 |
| 204203 | 707561 | Golden Acre | 2.10 | 0.32 | 0.09 | 3.28 | 0.05 | 0.07 | 0.01 | ND | 3.48 | 0.11 | 0.10 | 1.70 | 11.32 |
| NA | K142931 | Sudya | 0.01 | 0.01 | 0.04 | 0.01 | 3.36 | 0.03 | 0.06 | 0.26 | 4.03 | 2.55 | ND | 0.39 | 10.75 |
| NA | K004526 | Sudiya-146 | 0.05 | 1.62 | 0.11 | 0.03 | 3.85 | 0.52 | 0.07 | 0.18 | 2.51 | 1.09 | 0.22 | 0.15 | 10.40 |
| NA | K247741 | TJK-PHJ-2014-6-8 | 0.78 | 0.04 | 0.09 | 0.02 | 3.10 | 0.04 | 0.04 | ND | 3.72 | 0.69 | 0.27 | 0.11 | 8.90 |
| GSLs | PRO | EPI | SIN | GRA | GNA | 4HGBS | GER | GBS | 4MGBS | GNS | NGBS | Total GSL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GIB | −0.285** | −0.131 | 0.598** | −0.485** | −0.273** | 0.192* | −0.318** | 0.239** | −0.112 | 0.272** | 0.254** | 0.576** |
| PRO | 0.686** | −0.106 | 0.360** | 0.805** | 0.073 | 0.254** | 0.099 | −0.141 | −0.031 | −0.061 | 0.280** | |
| EPI | −0.065 | 0.260** | 0.592** | 0.032 | 0.192* | 0.231** | −0.195* | −0.001 | −0.097 | 0.296** | ||
| SIN | −0.555** | −0.050 | 0.110 | −0.271** | 0.006 | −0.180* | 0.263** | 0.113 | 0.658** | |||
| GRA | 0.199* | −0.008 | 0.555** | 0.193* | 0.336** | −0.069 | −0.010 | −0.061 | ||||
| GNA | 0.016 | 0.202* | −0.015 | −0.183* | -0.052 | −0.088 | 0.214** | |||||
| 4HGBS | −0.063 | 0.143 | 0.130 | 0.085 | 0.062 | 0.229** | ||||||
| GER | 0.045 | 0.251** | −0.164* | −0.059 | −0.019 | |||||||
| GBS | 0.089 | 0.132 | 0.411** | 0.654** | ||||||||
| 4MGBS | −0.028 | 0.028 | −0.017 | |||||||||
| GNS | 0.110 | 0.308** | ||||||||||
| NGBS | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | 0.393** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhandari, S.R.; Rhee, J.; Choi, C.S.; Jo, J.S.; Shin, Y.K.; Lee, J.G. Profiling of Individual Desulfo-Glucosinolate Content in Cabbage Head (Brassica oleracea var. capitata) Germplasm. Molecules 2020, 25, 1860. https://doi.org/10.3390/molecules25081860
Bhandari SR, Rhee J, Choi CS, Jo JS, Shin YK, Lee JG. Profiling of Individual Desulfo-Glucosinolate Content in Cabbage Head (Brassica oleracea var. capitata) Germplasm. Molecules. 2020; 25(8):1860. https://doi.org/10.3390/molecules25081860
Chicago/Turabian StyleBhandari, Shiva Ram, Juhee Rhee, Chang Sun Choi, Jung Su Jo, Yu Kyeong Shin, and Jun Gu Lee. 2020. "Profiling of Individual Desulfo-Glucosinolate Content in Cabbage Head (Brassica oleracea var. capitata) Germplasm" Molecules 25, no. 8: 1860. https://doi.org/10.3390/molecules25081860
APA StyleBhandari, S. R., Rhee, J., Choi, C. S., Jo, J. S., Shin, Y. K., & Lee, J. G. (2020). Profiling of Individual Desulfo-Glucosinolate Content in Cabbage Head (Brassica oleracea var. capitata) Germplasm. Molecules, 25(8), 1860. https://doi.org/10.3390/molecules25081860