Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources


Abstract
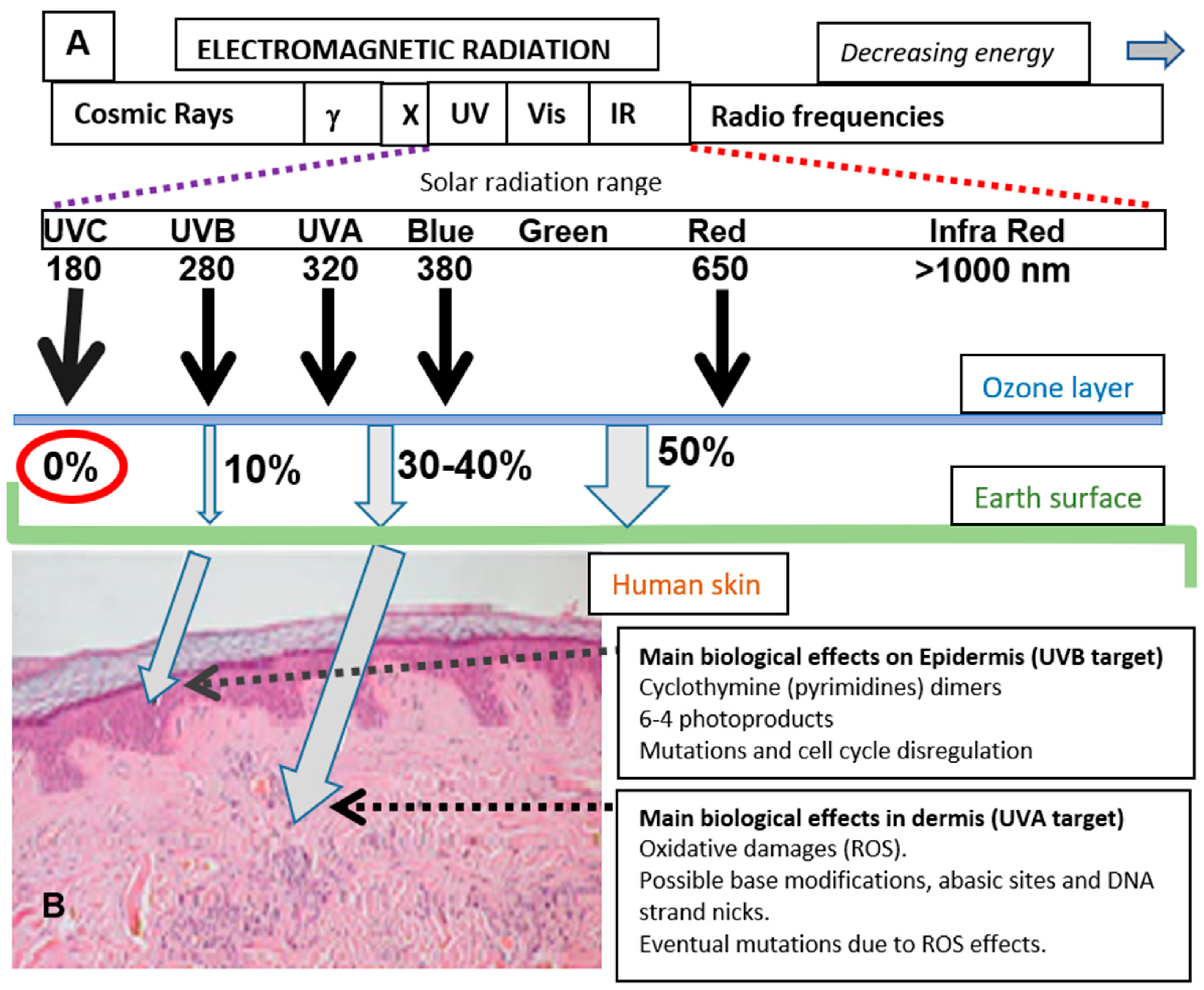
1. Solar Radiation and Skin Photodamage: General Concepts
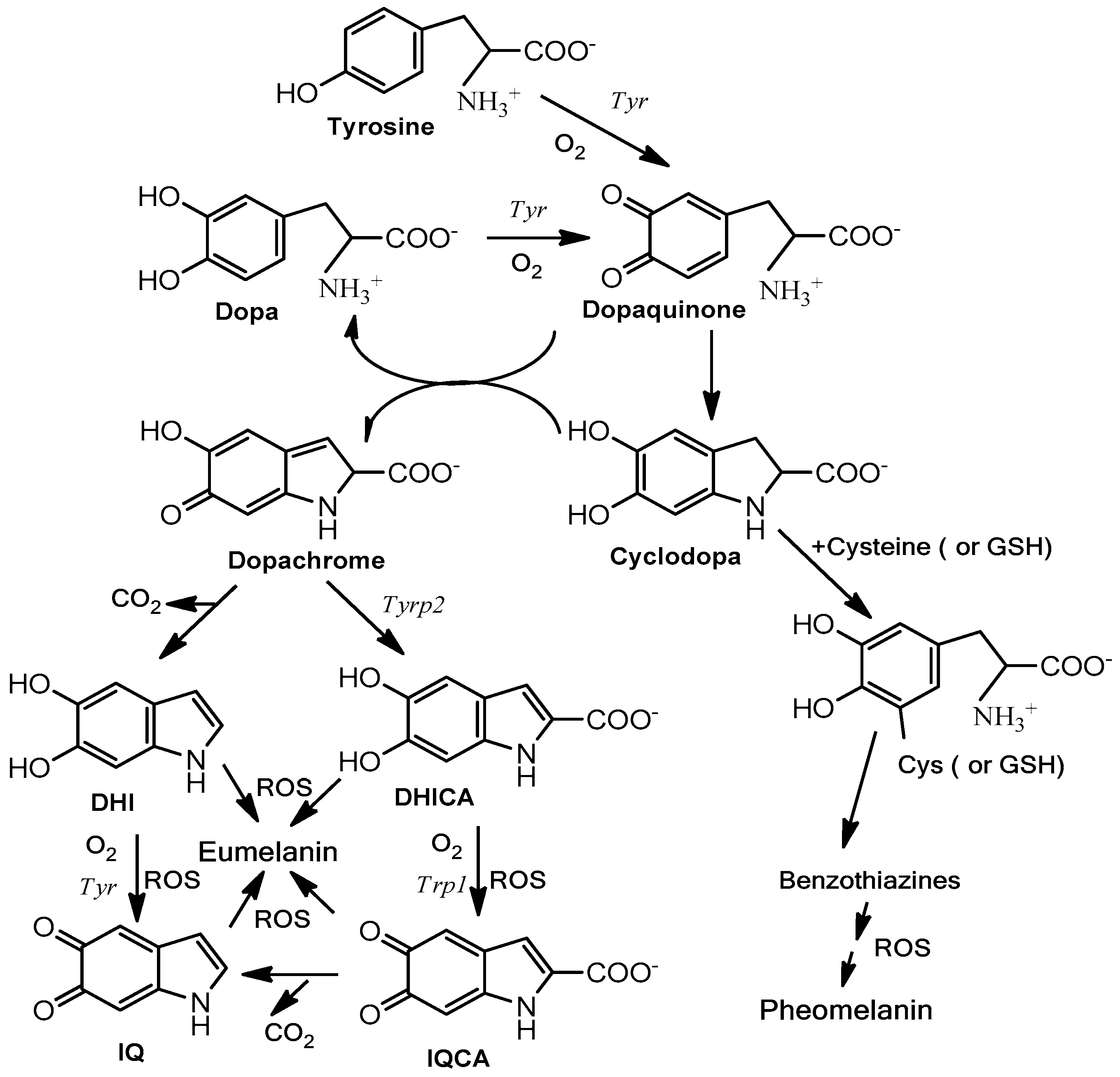
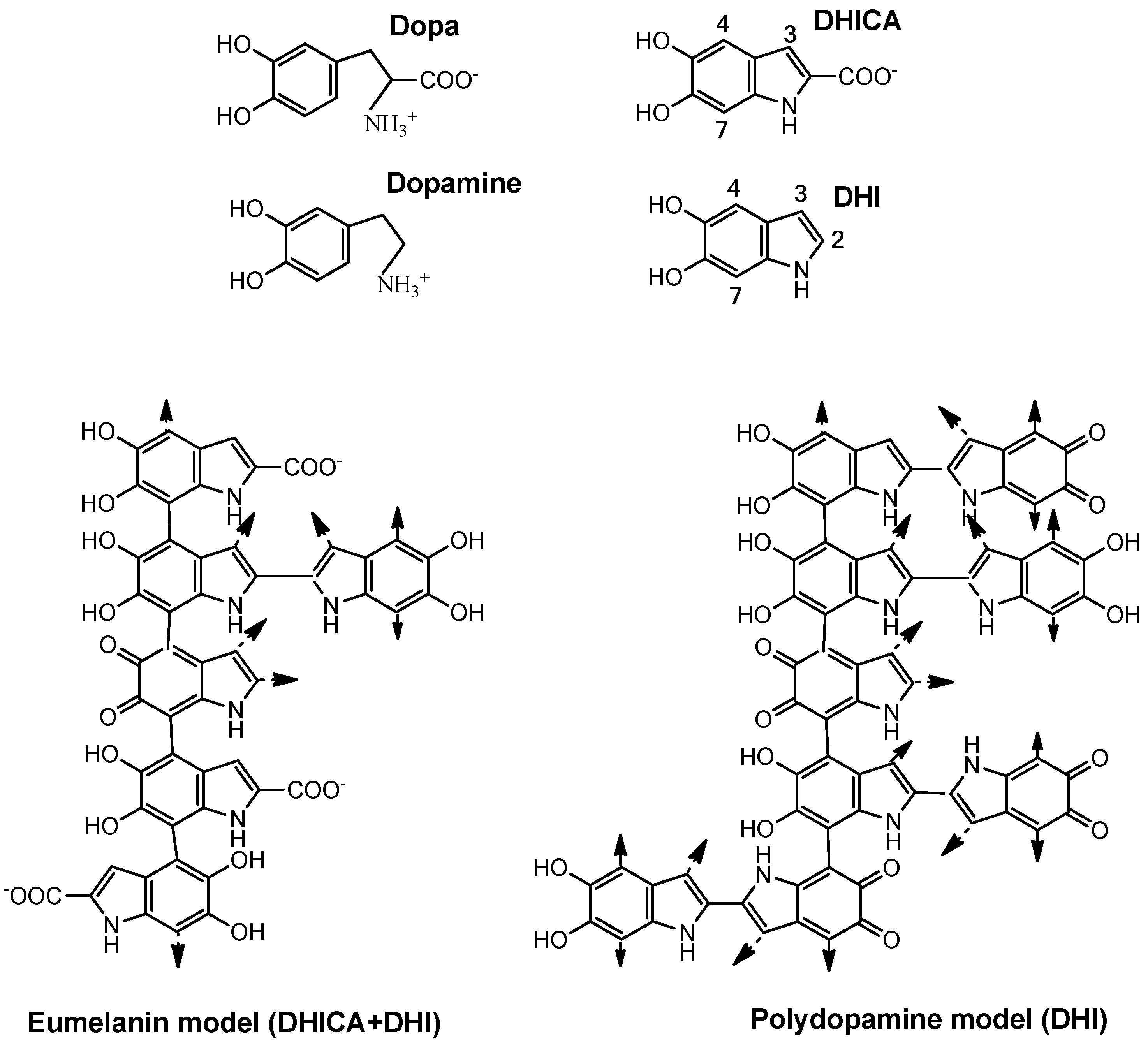
2. Natural Skin Pigments for Photoprotection: Melanin and Melanogenesis
3. Sunscreens: Parameters for Evaluating Photoprotection
4. Photoprotective and Antiaging Components in Sunscreens: Nature, Sources and Action
- (i)
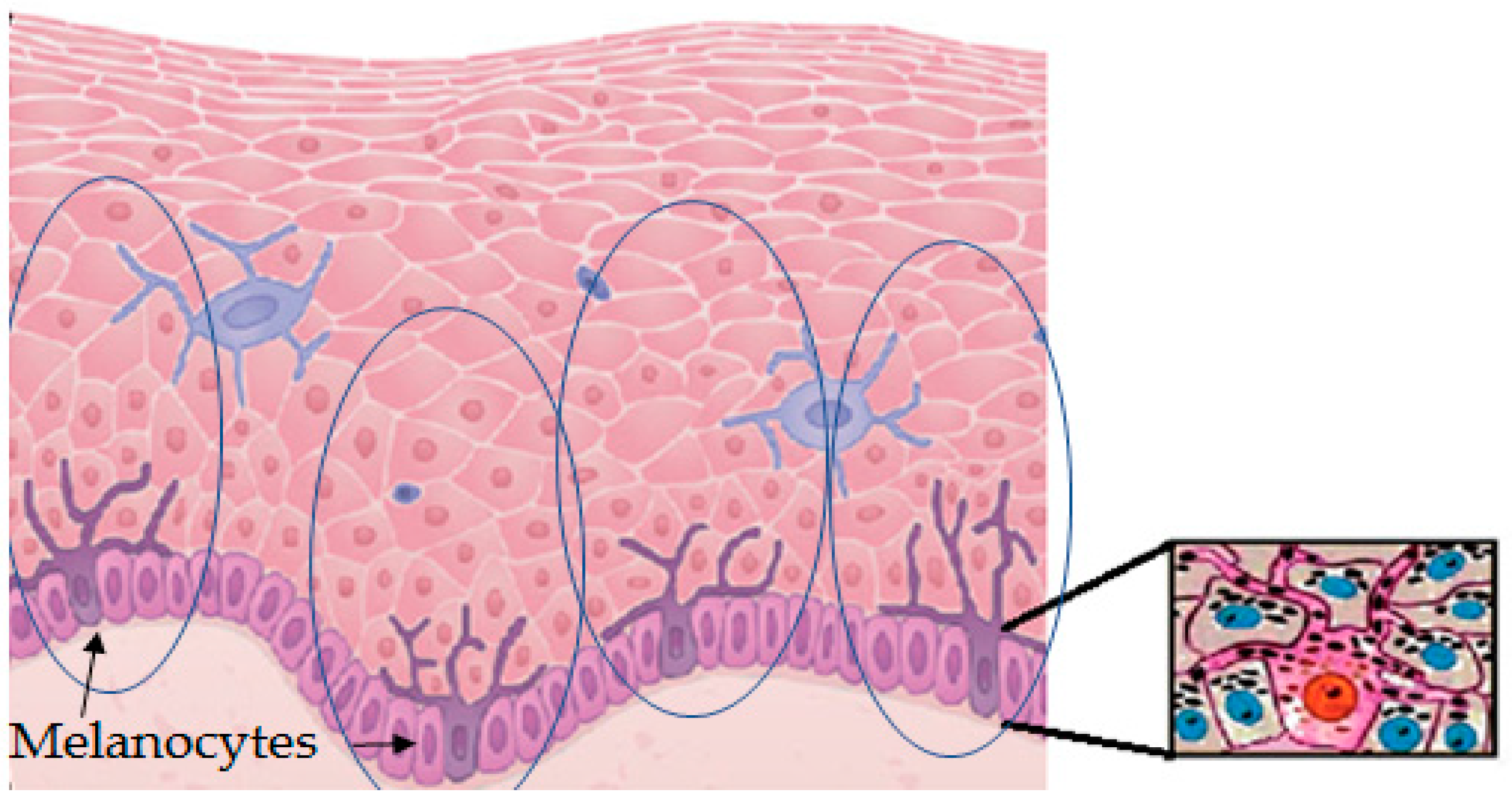
- Stimulation of natural protection through action on melanocytes and surrounding keratinocytes to stimulate the synthesis of endogenous melanin and its subsequent distribution through the melano-epidermal unit. Basically, they act by stimulating the tyrosinase activity and melanogenic proteins (Figure 3) and/or the subsequent transference of melanosomes to keratinocytes (Figure 2).
- (ii)
- Use of biopolymers related to natural melanin or unpolymerized molecules to cover the skin as an external shield against solar damage. Nevertheless, some of these biopolymers can partially penetrate inside epidermal keratinocytes.
- (iii)
- Stimulation of other mechanisms of the skin not directly related to pigmentation, but involved in retarding the aging, oxidizing, and inflammatory effects of sun exposure. Maintenance of hydration or collagen is somehow a way of photoprotection, minimizing accelerated wrinkling, sagging, and solar elastosis.
4.1. Natural Compounds Related to Animal Melanin
4.2. Natural Products from Plants, Herbs, and Marine Organisms
4.2.1. Plants and Herbs
4.2.2. Marine Organisms and Microalgae
5. Conclusions
Funding
Conflicts of Interest
References
- Madronich, S.; McKenzie, R.L.; Björn, L.O.; Caldwell, M.M. Changes in biologically active ultraviolet radiation reaching the Earth’s surface. J. Photochem. Photobiol. B 1998, 46, 5–19. [Google Scholar] [CrossRef]
- Juzeniene, A.; Moan, J. Beneficial effects of UV radiation other than via vitamin D production. Dermatoendocrinol 2012, 4, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Wacker, M.; Holick, M.F. Sunlight and Vitamin D: A global perspective for health. Dermatoendocrinol 2013, 5, 51–108. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Premi, S.; Wallisch, S.; Mano, C.M.; Weiner, A.B.; Bacchiocchi, A.; Wakamatsu, K.; Bechara, E.J.H.; Halaban, R.; Douki, T.; Brash, D.E. Photochemistry. Chemiexcitation of melanin derivatives induces DNA photoproducts long after UV exposure. Science 2015, 347, 842–847. [Google Scholar] [CrossRef]
- Solano, F. Photoprotection versus photodamage: Updating an old but still unsolved controversy about melanin. Polymer Intern. 2016, 65, 1276–1287. [Google Scholar] [CrossRef]
- Fisher, J.M.; Fisher, D.E. From suntan to skin cancers: Molecular pathways and prevention strategies. Targ. Oncol. 2008, 3, 41–44. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin Against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J. Invest. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef]
- Nordlund, J.J. The melanocyte and the epidermal melanin unit: An expanded concept. Dermatol. Clin. 2007, 25, 271–281. [Google Scholar] [CrossRef]
- Huang, Y.; Li, Y.; Hu, Z.; Yue, X.; Proetto, M.T.; Jones, Y.; Gianneschi, N.C. Mimicking melanosomes: Polydopamine nanoparticles as artificial microparasols. ACS Cent. Sci. 2017, 3, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Solano, F. Melanins: Skin pigments and much more-types, structural models, biological functions, and formation routes. New J. Sci. 2014, 498276. [Google Scholar] [CrossRef]
- Solano, F.; García-Borrón, J.C. Enzymology of melanin formation. In The Pigmentary System: Physiology and Pathophysiology; Blackwell Pub. Ltd.: Oxford, UK, 2006; pp. 261–281. [Google Scholar]
- Solano, F. On the metal cofactor in the tyrosinase family. Int. J. Mol. Sci. 2018, 19, 633. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Phenylthiourea Binding to Human Tyrosinase-Related Protein 1. Int. J. Mol. Sci. 2020, 21, 915. [Google Scholar] [CrossRef] [PubMed]
- Panzella, L.; Ebato, A.; Napolitano, A.; Koike, K. The Late Stages of Melanogenesis: Exploring the chemical facets and the application opportunities. Int. J. Mol. Sci. 2018, 19, 1753. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Edwards, C.; Gaskell, S.; Pearse, A.; Marks, R. Melanin content and distribution in the surface corneocyte with skin phototypes. Br. J. Derm. 1996, 135, 263–267. [Google Scholar] [CrossRef]
- Wei, M.X.; Weiyao, C.; Zhao, L.J.; Hong, Y.-L.; Nishiyama, Y.; Miyoshi, T.; Shawkey, M.D.; Dhinojwala, A. Elucidation of the hierarchical structure of natural eumelanins. J. R. Soc. Interface 2018, 15. [Google Scholar] [CrossRef]
- Simon, J.D.; Peles, D.N. The red and the black. Acc. Chem. Res. 2010, 43, 1452–1460. [Google Scholar] [CrossRef]
- Napolitano, A.; Panzella, L.; Monfrecola, G.; d’Ischia, M. Pheomelanin-induced oxidative stress: Bright and dark chemistry bridging red hair phenotype and melanoma. Pigment. Cell Melanoma Res. 2014, 27, 721–733. [Google Scholar] [CrossRef]
- Pathak, M.A. Sunscreens: Progress and perspectives on photoprotection of human skin against UVB and UVA radiation. J. Dermatol. 1996, 23, 783–800. [Google Scholar] [CrossRef]
- Sambandan, D.R.; Ratner, D. Sunscreens: An overview and update. J. Am. Acad. Dermatol. 2011, 64, 748–758. [Google Scholar] [CrossRef]
- Ngoc, L.T.N.; Tran, V.V.; Moon, J.-Y.; Chae, M.; Park, D.; Lee, Y.-C. Recent Trends of Sunscreen Cosmetic: An Update Review. Cosmetics 2019, 6, 64. [Google Scholar] [CrossRef]
- Geoffrey, K.; Mwangi, A.N.; Maru, S.M. Sunscreen products: Rationale for use, formulation development and regulatory considerations. Saudi Pharm. J. 2019, 27, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Dennis, L.K.; Beane Freeman, L.E.; Van Beek, M.J. Sunscreen use and the risk for melanoma: A quantitative review. Ann. Intern. Med. 2003, 139, 966–978. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). FDA Rules Regulations for Sunscreen. 2012. Available online: https://smartshield.com/news/reviews/54-resources/127-new-fda-rules-regulations-for-sunscreen (accessed on 1 March 2020).
- Osterwalder, U.; Herzog, B. Sun protection factors: Worldwide confusion. Br. J. Dermatol. 2009, 161, 13–24. [Google Scholar] [CrossRef]
- Moyal, D. UVA protection labeling and in vitro testing methods. Photochem. Photobiol. Sci. 2010, 9, 516–523. [Google Scholar] [CrossRef]
- Wang, S.Q.; Stanfield, J.W.; Osterwalder, U. In vitro assessments of UVA protection by popular sunscreens available in the United States. J. Am. Acad. Dermatol. 2008, 59, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Donglikar, M.; Deore, L.S. Sunscreens: A review. Pharmacogn. J. 2016, 8, 171–179. [Google Scholar] [CrossRef]
- De la Coba, F.; Aguilera, J.; Korbee, N.; de Gálvez, M.V.; Herrera-Ceballos, E.; Álvarez-Gómez, F.; Figueroa, F.L. UVA and UVB Photoprotective Capabilities of Topical Formulations Containing Mycosporine-like Amino Acids (MAAs) through Different Biological Effective Protection Factors (BEPFs). Marine Drugs. 2019, 14, 17. [Google Scholar] [CrossRef]
- Diffey, B.L.; Tanner, P.R.; Matts, P.J.; Nash, J.F. In vitro assessment of the broad-spectrum ultraviolet protection of sunscreen products. J. Am. Acad. Dermatol. 2000, 43, 1024–1035. [Google Scholar] [CrossRef]
- Kligman, A.M. Cosmeceuticals: A broad-spectrum category between cosmetics and drugs. In Cosmeceuticals and Active Cosmetics Drug Versus Cosmetics, 2nd ed.; Elsner, P., Maibach, H.I., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2005; pp. 1–9. [Google Scholar]
- Micillo, R.; Panzella, L.; Iacomino, M.; Prampolini, G.; Cacelli, I.; Ferretti, A.; Crescenzi, O.; Koike, K.; Napolitano, A.; d’Ischia, M. Eumelanin broadband absorption develops from aggregation-modulated chromophore interactions under structural and redox control. Sci. Rep. 2017, 7, 41532. [Google Scholar] [CrossRef] [PubMed]
- Panzella, L.; Gentile, G.; D’Errico, G.; Della Vecchia, N.F.; Errico, M.E.; Napolitano, A.; Carfagna, C.; d’Ischia, M. Atypical structural and π-electron features of a melanin polymer that lead to superior free-radical-scavenging properties. Angew. Chem. 2013, 52, 12684–12687. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Takekoshi, Y.; Arai, Y. Cosmetics Based on Naturally Derived Melanin-Coated Pigments. US Patent 5,380,359, 1 October 1995. [Google Scholar]
- Nacht, S. Melanin, Nature’s Own Sunscreen Polymer. In Cosmetic and Pharmaceutical Applications of Polymers; Gebelein, C.G., Cheng, T.C., Yang, V.C., Eds.; Springer: Boston, MA, USA, 1991; pp. 83–94. [Google Scholar] [CrossRef]
- Zahner, P. All natural sunscreen lotion. US Patent App. 10/643,270, 24 February 2005. [Google Scholar]
- Geng, J.; Tang, W.; Wan, X.; Zhou, Q.; Wang, X.-J.; Shen, P.; Lei, T.-C.; Chen, X.-D. Photoprotection of bacterial-derived melanin against ultraviolet A–induced cell death and its potential application as an active sunscreen. J. Eur. Acad. Dermatol. Venereol. JEADV 2008, 22, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Leong, H.; Katz, M.O.; Delk, A.; Nacht, S.; Berliner, D. Synthetic Melanin Aggregates. US Patent 4,855,144, 8 August 1989. [Google Scholar]
- Pawelek, J.; Osber, M.P.; Orlow, S.J. Synthetic Melanin As A Sunscreen And Tanning Agent. US Patent 5,384,116, 24 January 1995. [Google Scholar]
- Menon, I.A.; Haberman, H.F. Lipomelanin Sunscreen Composition. US Patent 5,750,093, 12 May 1998. [Google Scholar]
- Hyun, S.R.; Jung, C.K.; Kim, W.; Zhai, L.; Zhu, Q.Y.; Kim, J. Incorporation of melanin nanoparticles improves UV-shielding, mechanical and antioxidant properties of cellulose nanofiber based nanocomposite films. Mater. Commun. 2020, 24, 100984. [Google Scholar] [CrossRef]
- Micillo, R.; Iacomino, M.; Perfetti, M.; Panzella, L.; Koike, K.; D’Errico, G.; d’Ischia, M.; Napolitano, A. Unexpected impact of esterification on the antioxidant activity and (photo)stability of a eumelanin from 5,6-dihydroxyindole-2-carboxylic acid. Pigment Cell Melanoma Res. 2018, 31, 475–483. [Google Scholar] [CrossRef]
- d’Ischia, M.; Napolitano, A.; Ball, V.; Chen, C.-T.; Buehler, M.J. Polydopamine and Eumelanin: From Structure–Property Relationships to a Unified Tailoring Strategy. Acc. Chem. Res. 2014, 47, 3541–3550. [Google Scholar] [CrossRef]
- Dreyer, D.R.; Miller, D.J.; Freeman, B.D.; Paul, D.R.; Bielawski, C.W. Elucidating the structure of poly (dopamine). Langmuir 2012, 28, 6428–6435. [Google Scholar] [CrossRef]
- Solano, F. Melanin and Melanin-Related Polymers as Materials with Biomedical and Biotechnological Applications—Cuttlefish Ink and Mussel Foot Proteins as Inspired Biomolecules. Int. J. Mol. Sci. 2017, 18, 1561. [Google Scholar] [CrossRef]
- Fajuyigbe, D.; Lwin, S.M.; Diffey, B.L.; Baker, R.; Tobin, D.J.; Sarkany, R.P.E.; Young, A.R. Melanin distribution in human epidermis affords localized protection against DNA photodamage and concurs with skin cancer incidence difference in extreme phototypes. FASEB J. 2018, 32, 3700–3706. [Google Scholar] [CrossRef]
- Grieco, C.; Empey, J.M.; Kohl, F.R.; Kohler, B. Probing eumelanin photoprotection using a catechol: Quinone heterodimer model system. Faraday Discuss. 2019, 216, 520–537. [Google Scholar] [CrossRef]
- Hollinger, J.C.; Angra, K.; Halder, R.M. Are Natural Ingredients Effective in the Management of Hyperpigmentation? A Systematic Review. J. Clin. Aesthetic Dermatol. 2018, 11, 28–37. [Google Scholar]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Niu, C.; Liao, L.-X.; Dou, J.; Habasi, M.; Aisa, H.A. An Isoxazole Chalcone Derivative Enhances Melanogenesis in B16 Melanoma Cells via the Akt/GSK3 Catenin Signaling Pathways. Molecules 2017, 22, 2077. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Kim, M.-Y.; Kang, E.Y.; Kim, Y.B.; Kim, B.S.; Park, S.-M.; Hyun, C.-G. Anti-Melanogenic Effects of Hydroxyectoine via MITF Inhibition by JNK, p38, and AKT Pathways in B16F10 Melanoma Cells. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Dermatol. 2012, 67, 1013–1024. [Google Scholar] [CrossRef]
- Jenkins, G.J.; Stephens, L.A.; Masnavi, N.; Parry, J.M. Molecular analysis of the chemoprotective effects of topical sunscreen and vitamin C in preventing UV-induced and reactive oxygen species-induced DNA damage, respectively, using the PCR inhibition methodology. Anticancer Res. 2002, 22, 3873–3877. [Google Scholar] [PubMed]
- McVean, M.; Liebler, D.C. Prevention of DNA photodamage by vitamin E compounds and sunscreens: Roles of ultraviolet absorbance and cellular uptake. Mol. Carcinog. 1999, 24, 169–176. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Beta-carotene and other carotenoids in protection from sunlight. Am. J. Clin. Nutr. 2012, 96, 1179S–1184S. [Google Scholar] [CrossRef]
- Dzialo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Szopa, J.; Kulma, A. The potential of plant phenolics in prevention and therapy of skin disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef]
- de Alencar Filho, J.M.T.; Sampaio, P.A.; Valença, E.C.; da Silva Almeida, G.; Rolim, L.A.; Nunes, X.P.; da Cruz Araújo, E.C. Flavonoids as photoprotective agents: A systematic review. J. Med. Plants Res. 2016, 10, 848–864. [Google Scholar]
- Sandmann, G. Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures. Antioxidants 2019, 8, 219. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, D.; Williams, J.A.; Kučerka, N.; Atkinson, J.; Wassall, S.R.; Katsaras, J.; Harroun, T.A. Tocopherol activity correlates with its location in a membrane: A new perspective on the antioxidant vitamin E. J. Amer. Chem. Soc. 2013, 135, 7523–7533. [Google Scholar] [CrossRef] [PubMed]
- Anitha, D.; Reddy, K.Y.; Venkatesh, P.; Raani, M.J. A review-herbal sunscreen agents on skin protection. Eur. J. Pharm. Med. Res. 2016, 3, 308–313. [Google Scholar]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Choi, S.W.; Chung, M.-H. A review on the relationship between aloe vera components and their biologic effects. Semin. Integr. Med. 2003, 1, 53–62. [Google Scholar] [CrossRef]
- Richardson, J.; Smith, J.E.; McIntyre, M.; Thomas, R.; Pilkington, K. Aloe Vera for Preventing Radiation-induced Skin Reactions: A Systematic Literature Review. Clin. Oncol. 2005, 17, 478–484. [Google Scholar] [CrossRef]
- Park, K.; Choi, H.S.; Hong, Y.H.; Jung, E.Y.; Suh, H.J. Cactus cladodes (Opuntia humifusa) extract minimizes the effects of UV irradiation on keratinocytes and hairless mice. Pharm. Biol. 2017, 55, 1032–1040. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Elmets, C.A. Green tea polyphenolic antioxidants and skin photoprotection. Int. J. Oncol. 2001, 18, 1307–1313. [Google Scholar] [CrossRef]
- Afaq, F.; Zaid, M.A.; Khan, N.; Dreher, M.; Mukhtar, H. Protective effect of pomegranate-derived products on UVB-mediated damage in human reconstituted skin. Exp. Dermatol. 2009, 18, 553–561. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Kim, J.-H.; Youn Cho, J.-Y. Photo-aging protective effects of Ranunculus bulumei methanol extract. Evid.-Based Complementary Altern. Med. 2020, in press. [Google Scholar]
- Sopyan, I.; Gozali, D.; Tiassetiana, S. Formulation of tomato extracts (Solanum lycopersicum L.) as a sunscreen lotion. Natl. J. Physiol. Pharm. Pharmacol. 2017, 8, 453–458. [Google Scholar] [CrossRef]
- Maheshwar, G.H.; Patil, B.S.; Prashant, D. Comparative sun protection factor determination fo fresh fruits extract of cucumber vs marketed cosmetic formulation. Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 55–59. [Google Scholar]
- Shenoy, P.; Khot, S.; Chavan, M.; Takawale, J.; Singh, S. Study of sunscreen activity of aqueous, methanol and acetone extracts of leaves of Pongamia pinnata (L.) pierre, fabaceae. Int. J. Green Pharm. 2010, 4, 270. [Google Scholar] [CrossRef]
- Ortel, B.; Gange, R.W. An Action Spectrum for the Elicitation of Erythema in Skin persistently Sensitized by Photobound 8-Methoxypsoralen. J. Invest. Dermatol. 1990, 94, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Solano, F.; Briganti, S.; Picardo, M.; Ghanem, G. Hypopigmenting agents: An updated review on biological, chemical and clinical aspects. Pigment Cell Res. 2003, 19, 550–571. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.-H. Inhibitors of Melanogenesis: An Updated Review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef] [PubMed]
- Na, J.-I.; Shin, J.-W.; Choi, H.-R.; Kwon, S.-H.; Park, K.C. Resveratrol as a Multifunctional Topical Hypopigmenting Agent. Int. J. Mol. Sci. 2019, 20, 956. [Google Scholar] [CrossRef]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef]
- Yun, C.-Y.; Ko, S.M.; Choi, Y.P.; Kim, Y.I. α-Viniferin Improves Facial Hyperpigmentation via Accelerating Feedback Termination of cAMP/PKA-Signaled Phosphorylation Circuit in Facultative Melanogenesis. Theranostics 2018, 8, 2031–2043. [Google Scholar] [CrossRef]
- Takekoshi, S.; Nagata, H.; Kitatani, K. Flavonoids enhance melanogenesis in human melanoma cells. Tokai J. Exp. Clin. Med. 2014, 39, 116–121. [Google Scholar]
- Takekoshi, S.; Matsuzaki, K.; Kitatani, K. Quercetin stimulates melanogenesis in hair follicle melanocyte of the mouse. Tokai J. Exp. Clin. Med. 2013, 38, 129–134. [Google Scholar]
- Taguchi, N.; Hata, T.; Kamiya, E.; Kobayashi, A.; Aoki, H.; Kunisada, T. Reduction in human hair graying by sterubin, an active flavonoid of Eriodictyon angustifolium. J. Dermatol. Sci. 2018, 92, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.J.; Zou, J.; Zou, T.; Shang, H.; Sun, L.Y. A Newly Authenticated Compound from Traditional Chinese Medicine Decoction Induces Melanogenesis in B16-F10 Cells by Increasing Tyrosinase Activity. Evid.-Based Complementary Altern. Med. 2018, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, J.Y.; Cho, Y.; Ahn, E.; Jeong, W.; Shin, H.S.; Kim, M.; Oh, J.S. Anti-melanogenic and Anti-oxidant Activities of an Ethanolic Extract of Kummerowia striata and its active compounds, p-coumaric acid and quercetin. Preprints 2018, 2018060341. [Google Scholar] [CrossRef]
- Choi, M.-C.; Shin, H.-J. Anti-Melanogenesis Effect of Quercetin. Cosmetics 2016, 3, 18. [Google Scholar] [CrossRef]
- Pallela, R. Antioxidants from Marine Organisms and Skin Care. In Systems Biology of Free Radicals and Antioxidants; Springer-Verlag: Berlin, Germany, 2012; pp. 3771–3783. [Google Scholar] [CrossRef]
- Hwang, J.; Kim, D.; Park, J.S.; Park, H.J.; Shin, J.; Lee, S.K. Photoprotective Activity of Topsentin, A Bis(Indole) Alkaloid from the Marine Sponge Spongosorites genitrix, by Regulation of COX-2 and Mir-4485 Expression in UVB-Irradiated Human Keratinocyte Cells. Marine Drugs 2020, 18, 87. [Google Scholar] [CrossRef]
- Sripada, L.; Singh, K.; Lipatova, A.V.; Singh, A.; Prajapati, P.; Tomar, D.; Bhatelia, K.; Roy, M.; Singh, R.; Godbole, M.M.; et al. Hsa-miR-4485 regulates mitochondrial functions and inhibits the tumorigenicity of breast cancer cells. J. Mol. Med 2017, 95, 641–651. [Google Scholar] [CrossRef]
- Zanella, L.; Alam, M.A. Extracts and Bioactives from Microalgae (Sensu Stricto): Opportunities and Challenges for a New Generation of Cosmetics. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M., Xu, J.L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 295–349. [Google Scholar]
- Safafar, H.; Van Wagenen, J.; Moller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Marine Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; De Paepe, D.; Baart, G.J.E.; De Cooman, L. Detection of flavonoids in microalgae from different evolutionary lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.-S. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Marine Drugs 2018, 16, 26. [Google Scholar] [CrossRef]
- Lee, A.; Kim, J.Y.; Heo, J.; Cho, D.-H.; Kim, H.-S.; An, I.-S.; An, S.; Bae, S. The inhibition of melanogenesis via the PKA and ERK signaling pathways by Chlamydomonas reinhardtii extract in B16F10 melanoma cells and artificial human skin equivalents. J. Microbiol. Biotechnol. 2018, 28, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Letsiou, S.; Kalliampakou, K.; Gardikis, K.; Mantecon, L.; Infante, C.; Chatzikonstantinou, M.; Labrou, N.E.; Flemetakis, E. Skin protective effects of Nannochloropsis gaditana extract on H2O2-stressed human dermal fibroblasts. Front. Mar. Sci. 2017, 4, 221. [Google Scholar] [CrossRef]
- Ando, H.; Funasaka, Y.; Oka, M.; Ohashi, A.; Furumura, M.; Matsunaga, J.; Matsunaga, N.; Hearing, V.J.; Ichihashi, M. Possible involvement of proteolytic degradation of tyrosinase in the regulatory effect of fatty acids on melanogenesis. J. Lipid Res. 1999, 40, 1312–1316. [Google Scholar] [PubMed]
- Chiang, H.-M.; Pan, Y.-Y.; Chen, C.-W.; Wen, K.-C. Fatty acids and their related products modulate melanogenesis. Focus on skin care: Ethnic, whitening & tanning—Supplement to household and personal care today. Skin Care 2011, 6, 15–19. [Google Scholar]
- Hartmann, K.B.; Karsten, U.; Remias, B.; Ganzera, M. Analysis of mycosporine-like amino acids in selected algae and cyanobacteria by hydrophilic interaction liquid chromatography and a novel MAA from the red alga Catenella repens. Mar. Drugs 2015, 13, 6291–6305. [Google Scholar] [CrossRef]
- Lawrence, K.P.; Gacesa, R.; Long, P.F.; Young, A.R. Molecular photoprotection of human keratinocytes in vitro by the naturally occurring mycosporine-like amino acid palythine. Br. J. Derm. 2018, 178, 1353–1363. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
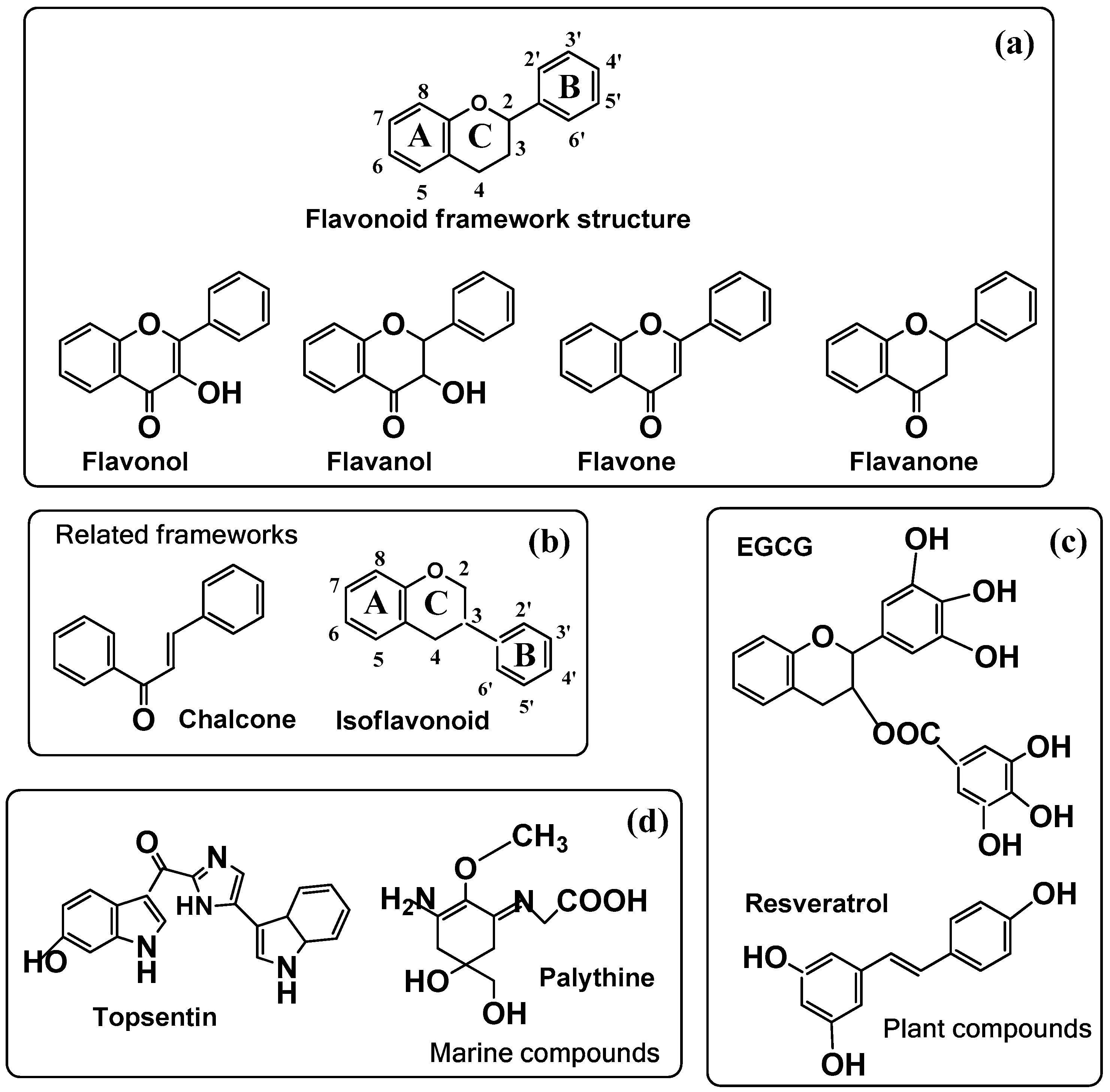{kind=link}
| Agent | Melanin action | Chemical name |
|---|---|---|
| Flavonols | ||
| Galangin | − | 3,5,7-trihydroxyflavone |
| Kaempferol | + | 3,4′,5,7-tetrahydroxyflavone |
| Quercetin | + | 3,3′,4′,5,7-pentahydroxyflavone |
| Rutin | − | 3,3′(O-glycoside),4′,5,7-pentahydroxyflavone |
| Myricetin | − | 3,3′,4′,5′,5,7- hexahydroxyflavone |
| Fisetin | + | 3,3′,4′,7-tetrahydroxyflavone |
| Robinetin | − | 3,3′,4′,5′,7-pentahydroxyflavone |
| Flavones | ||
| Chrysin | + | 5,7-dihydroxyflavone |
| Apigenin | + | 4′,5,7-trihydroxyflavone |
| Apigenin-7-butylene Glycoside | + | 4′,5,7-(O-butyl-glycoside)-trihydroxyflavone |
| Luteoin | + | 3′,4′,5,7-tetrahydroxyflavone |
| Flavanones | ||
| Narigin | − | 4′,5,7(O-glycoside)-dihydroxyflavanone |
| Naringenin | − | 4′,5,7-trihydroxyflavanone |
| Sterubin | + | 3′,4′,5,7(methoxy)- tetrahydroxyflavanone |
| Flavonoid-related molecules | ||
| EGCG | − | EpiGalloCatechin Gallate (actually a Flavanol) |
| Genestein (isoflavonoid) | + | 4′,5,7- trihydroxyisoflavone |
| Chalcone | + | Benzyliden-acetophenone |
| Resveratrol | − | 3,5,4′-trihydroxy-trans-stilbene |
| Topsentin | + | 6-hydroxy-1H-indol-3-yl)-[5-(1H-indol-3-yl)-1H-imidazol-2-yl]-methanone |
| Palythine | + | 2-[[3-amino-5-hydroxy-5-(hydroxymethyl)-2-methoxycyclohex-2-en-1-ylidene]-amino]-acetic acid |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solano, F. Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources. Molecules 2020, 25, 1537. https://doi.org/10.3390/molecules25071537
Solano F. Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources. Molecules. 2020; 25(7):1537. https://doi.org/10.3390/molecules25071537
Chicago/Turabian StyleSolano, Francisco. 2020. "Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources" Molecules 25, no. 7: 1537. https://doi.org/10.3390/molecules25071537
APA StyleSolano, F. (2020). Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources. Molecules, 25(7), 1537. https://doi.org/10.3390/molecules25071537