Salvinorin A Does Not Affect Seizure Threshold in Mice


Abstract
1. Introduction
2. Results
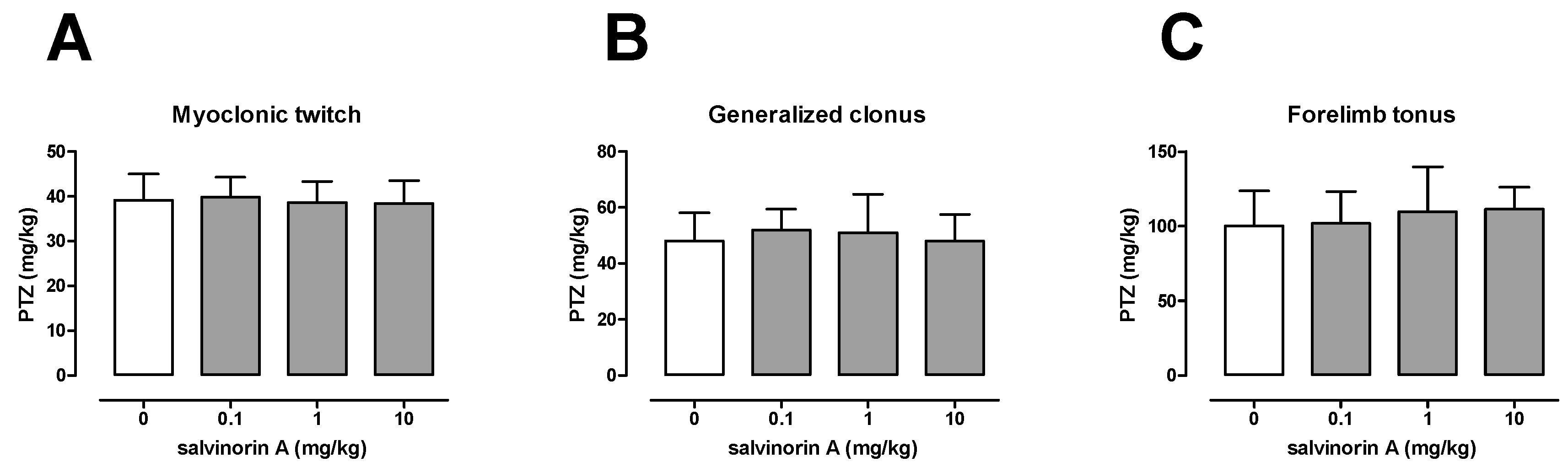
2.1. Effect of Salvinorin A in the i.v. PTZ Seizure Threshold Test
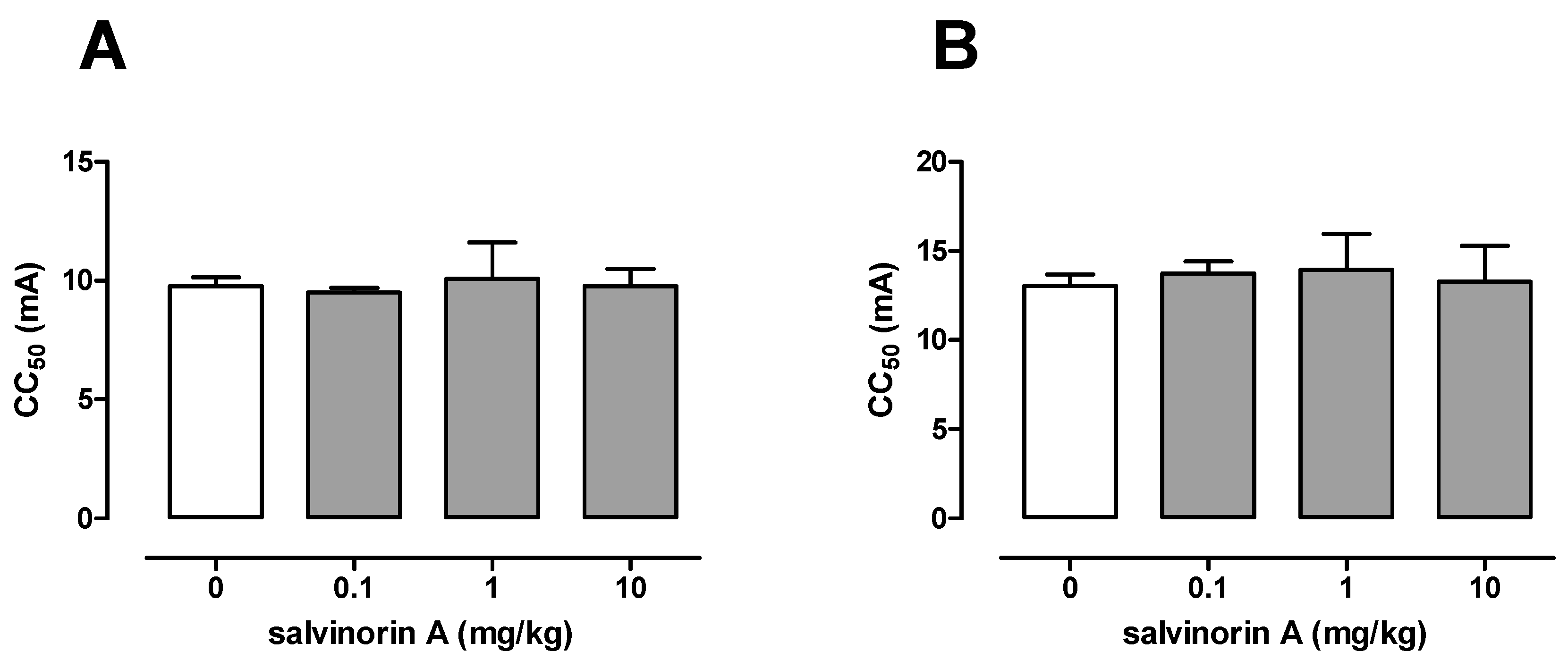
2.2. Effect of Salvinorin A in the Maximal Electroshock Seizure Threshold Test
2.3. Effect of Salvinorin A in the 6 Hz-Induced Seizure Threshold Test
2.4. Effect of Salvinorin A on Muscular Strength and Motor Coordination
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Treatment
4.3. Intravenous PTZ Seizure Threshold Test
4.4. Maximal Electroshock Seizure Threshold Test
4.5. 6 Hz-Induced Psychomotor Seizure Threshold Test
4.6. Grip Strength Test
4.7. Chimney Test
4.8. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Casselman, I.; Nock, C.J.; Wohlmuth, H.; Weatherby, R.P.; Heinrich, M. From local to global-fifty years of research on Salvia divinorum. J. Ethnopharmacol. 2014, 151, 768–783. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.; Domingos, S.; Gallardo, E.; Martinho, A. A unique natural selective kappa-opioid receptor agonist, salvinorin A, and its roles in human therapeutics. Phytochemistry 2017, 137, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Butelman, E.R.; Kreek, M.J. Salvinorin A, a kappa-opioid receptor agonist hallucinogen: pharmacology and potential template for novel pharmacotherapeutic agents in neuropsychiatric disorders. Front. Pharmacol. 2015, 6, 190. [Google Scholar] [PubMed]
- Cunningham, C.W.; Rothman, R.B.; Prisinzano, T.E. Neuropharmacology of the naturally occurring kappa-opioid hallucinogen salvinorin A. Pharmacol. Rev. 2011, 63, 316–347. [Google Scholar] [CrossRef]
- Roth, B.L.; Baner, K.; Westkaemper, R.; Siebert, D.; Rice, K.C.; Steinberg, S.; Ernsberger, P.; Rothman, R.B. Salvinorin A: A potent naturally occurring nonnitrogenous kappa opioid selective agonist. Proc. Natl. Acad. Sci. USA 2002, 99, 11934–11939. [Google Scholar] [CrossRef]
- Yan, F.; Mosier, P.D.; Westkaemper, R.B.; Stewart, J.; Zjawiony, J.K.; Vortherms, T.A.; Sheffler, D.J.; Roth, B.L. Identification of the molecular mechanisms by which the diterpenoid salvinorin A binds to kappa-opioid receptors. Biochemistry 2005, 44, 8643–8651. [Google Scholar] [CrossRef]
- Willmore-Fordham, C.B.; Krall, D.M.; McCurdy, C.R.; Kinder, D.H. The hallucinogen derived from Salvia divinorum, salvinorin A, has kappa-opioid agonist discriminative stimulus effects in rats. Neuropharmacology 2007, 53, 481–486. [Google Scholar] [CrossRef]
- Crowley, N.A.; Kash, T.L. Kappa opioid receptor signaling in the brain: Circuitry and implications for treatment. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 62, 51–60. [Google Scholar] [CrossRef]
- Dunn, A.D.; Reed, B.; Erazo, J.; Ben Ezra, A.; Kreek, M.J. Signaling properties of structurally diverse kappa opioid receptor ligands: Toward in vitro models of in vivo responses. ACS Chem. Neurosci. 2019, 10, 3590–3600. [Google Scholar] [CrossRef]
- Loacker, S.; Sayyah, M.; Wittmann, W.; Herzog, H.; Schwarzer, C. Endogenous dynorphin in epileptogenesis and epilepsy: Anticonvulsant net effect via kappa opioid receptors. Brain 2007, 130, 1017–1028. [Google Scholar] [CrossRef]
- Carlezon, W.A., Jr.; Beguin, C.; DiNieri, J.A.; Baumann, M.H.; Richards, M.R.; Todtenkopf, M.S.; Rothman, R.B.; Ma, Z.; Lee, D.Y.; Cohen, B.M. Depressive-like effects of the kappa-opioid receptor agonist salvinorin A on behavior and neurochemistry in rats. J. Pharmacol. Exp. Ther. 2006, 316, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Butelman, E.R.; Schlussman, S.D.; Ho, A.; Kreek, M.J. Effects of the plant-derived hallucinogen salvinorin A on basal dopamine levels in the caudate putamen and in a conditioned place aversion assay in mice: Agonist actions at kappa opioid receptors. Psychopharmacology 2005, 179, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Braida, D.; Capurro, V.; Zani, A.; Rubino, T.; Vigano, D.; Parolaro, D.; Sala, M. Potential anxiolytic- and antidepressant-like effects of salvinorin A, the main active ingredient of Salvia divinorum, in rodents. Br. J. Pharmacol. 2009, 157, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Schwarzer, C. The opioid system in temporal lobe epilepsy: Functional role and therapeutic potential. Front. Mol. Neurosci. 2017, 10, 245. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.L. Electroshock. In Neuropharmacology Methods in Epilepsy Research; Peterson, S.L., Albertson, T.E., Eds.; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Barton, M.E.; Klein, B.D.; Wolf, H.H.; White, H.S. Pharmacological characterization of the 6 Hz psychomotor seizure model of partial epilepsy. Epilepsy Res. 2001, 47, 217–227. [Google Scholar] [CrossRef]
- Listos, J.; Merska, A.; Fidecka, S. Pharmacological activity of salvinorin A, the major component of Salvia divinorum. Pharmacol. Rep. 2011, 63, 1305–1309. [Google Scholar] [CrossRef]
- Frey, H.H. Effect of mu- and kappa-opioid agonists on the electroconvulsive seizure threshold in mice and antagonism by naloxone and MR 2266. Pharmacol. Toxicol. 1988, 62, 150–154. [Google Scholar] [CrossRef]
- Manocha, A.; Mediratta, P.K.; Sharma, K.K. Studies on the anticonvulsant effect of U50488H on maximal electroshock seizure in mice. Pharmacol. Biochem. Behav. 2003, 76, 111–117. [Google Scholar] [CrossRef]
- Tortella, F.C.; Robles, L.; Echevarria, E.; Hunter, J.C.; Hughes, J. PD117302, a selective non-peptide opioid kappa agonist, protects against NMDA and maximal electroshock convulsions in rats. Life Sci. 1990, 46, L1–L7. [Google Scholar] [CrossRef]
- Tortella, F.C.; Robles, L.; Holaday, J.W. U50,488, a highly selective kappa opioid: Anticonvulsant profile in rats. J. Pharmacol. Exp. Ther. 1986, 237, 49–53. [Google Scholar]
- Kaminski, R.M.; Witkin, J.M.; Shippenberg, T.S. Pharmacological and genetic manipulation of kappa opioid receptors: Effects on cocaine- and pentylenetetrazol-induced convulsions and seizure kindling. Neuropharmacology 2007, 52, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Bausch, S.B.; Esteb, T.M.; Terman, G.W.; Chavkin, C. Administered and endogenously released kappa opioids decrease pilocarpine-induced seizures and seizure-induced histopathology. J. Pharmacol. Exp. Ther. 1998, 284, 1147–1155. [Google Scholar] [PubMed]
- Przewłocka, B.; Machelska, H.; Lasoń, W. Kappa opioid receptor agonists inhibit the pilocarpine-induced seizures and toxicity in the mouse. Eur. Neuropsychopharmacol. 1994, 4, 527–533. [Google Scholar] [CrossRef]
- Yajima, Y.; Narita, M.; Takahashi-Nakano, Y.; Misawa, M.; Nagase, H.; Mizoguchi, H.; Tseng, L.F.; Suzuki, T. Effects of differential modulation of mu-, delta- and kappa-opioid systems on bicuculline-induced convulsions in the mouse. Brain Res. 2000, 862, 120–126. [Google Scholar] [CrossRef]
- Lasoń, W.; Simpson, J.N.; McGinty, J.F. The interaction between kappa-opioid agonist, U-50, 488H, and kainic acid: Behavioral and histological assessments. Brain Res. 1989, 482, 333–339. [Google Scholar] [CrossRef]
- Singh, L.; Vass, C.A.; Hunter, J.C.; Woodruff, G.N.; Hughes, J. The anticonvulsant action of CI-977, a selective kappa-opioid receptor agonist: A possible involvement of the glycine/NMDA receptor complex. Eur. J. Pharmacol. 1990, 191, 477–480. [Google Scholar] [CrossRef]
- Albertson, T.E.; Joy, R.M.; Stark, L.G. Modification of kindled amygdaloid seizures by opiate agonists and antagonists. J. Pharmacol. Exp. Ther. 1984, 228, 620–627. [Google Scholar]
- Queenan, B.N.; Dunn, R.L.; Santos, V.R.; Feng, Y.; Huizenga, M.N.; Hammack, R.J.; Vicini, S.; Forcelli, P.A.; Pak, D.T.S. Kappa opioid receptors regulate hippocampal synaptic homeostasis and epileptogenesis. Epilepsia 2018, 59, 106–122. [Google Scholar] [CrossRef]
- Becker, A.; Braun, H.; Schroder, H.; Grecksch, G.; Hollt, V. Effects of enadoline on the development of pentylenetetrazol kindling, learning performance, and hippocampal morphology. Brain Res. 1999, 823, 191–197. [Google Scholar] [CrossRef]
- Hooker, J.M.; Patel, V.; Kothari, S.; Schiffer, W.K. Metabolic changes in the rodent brain after acute administration of salvinorin A. Mol. Imaging Biol. 2009, 11, 137–143. [Google Scholar] [CrossRef]
- Placzek, M.S.; Van de Bittner, G.C.; Wey, H.Y.; Lukas, S.E.; Hooker, J.M. Immediate and persistent effects of salvinorin a on the kappa opioid receptor in rodents, monitored in vivo with PET. Neuropsychopharmacology 2015, 40, 2865–2872. [Google Scholar] [CrossRef] [PubMed]
- Potter, D.N.; Damez-Werno, D.; Carlezon, W.A., Jr.; Cohen, B.M.; Chartoff, E.H. Repeated exposure to the kappa-opioid receptor agonist salvinorin A modulates extracellular signal-regulated kinase and reward sensitivity. Biol. Psychiatry 2011, 70, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, K.; Inan, S.; Siebert, D.; Holzgrabe, U.; Lee, D.Y.; Huang, P.; Li, J.G.; Cowan, A.; Liu-Chen, L.Y. Comparison of pharmacological activities of three distinct kappa ligands (Salvinorin A, TRK-820 and 3FLB) on kappa opioid receptors in vitro and their antipruritic and antinociceptive activities in vivo. J. Pharmacol. Exp. Ther. 2005, 312, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Rothman, R.B.; Murphy, D.L.; Xu, H.; Godin, J.A.; Dersch, C.M.; Partilla, J.S.; Tidgewell, K.; Schmidt, M.; Prisinzano, T.E. Salvinorin A: Allosteric interactions at the mu-opioid receptor. J. Pharmacol. Exp. Ther. 2007, 320, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Seeman, P.; Guan, H.C.; Hirbec, H. Dopamine D2High receptors stimulated by phencyclidines, lysergic acid diethylamide, salvinorin A, and modafinil. Synapse 2009, 63, 698–704. [Google Scholar] [CrossRef]
- Xu, X.; Ma, S.; Feng, Z.; Hu, G.; Wang, L.; Xie, X.Q. Chemogenomics knowledgebase and systems pharmacology for hallucinogen target identification-Salvinorin A as a case study. J. Mol. Graph. Model. 2016, 70, 284–295. [Google Scholar] [CrossRef][Green Version]
- Fantegrossi, W.E.; Kugle, K.M.; Valdes, L.J., III; Koreeda, M.; Woods, J.H. Kappa-opioid receptor-mediated effects of the plant-derived hallucinogen, salvinorin A, on inverted screen performance in the mouse. Behav. Pharmacol. 2005, 16, 627–633. [Google Scholar] [CrossRef]
- Kivell, B.M.; Paton, K.F.; Kumar, N.; Morani, A.S.; Culverhouse, A.; Shepherd, A.; Welsh, S.A.; Biggerstaff, A.; Crowley, R.S.; Prisinzano, T.E. Kappa opioid receptor agonist Mesyl Sal B attenuates behavioral sensitization to cocaine with fewer aversive side-effects than Salvinorin A in Rodents. Molecules 2018, 23, 2602. [Google Scholar] [CrossRef]
- White, K.L.; Robinson, J.E.; Zhu, H.; DiBerto, J.F.; Polepally, P.R.; Zjawiony, J.K.; Nichols, D.E.; Malanga, C.J.; Roth, B.L. The G protein-biased kappa-opioid receptor agonist RB-64 is analgesic with a unique spectrum of activities in vivo. J. Pharmacol. Exp. Ther. 2015, 352, 98–109. [Google Scholar] [CrossRef]
- Bansinath, M.; Ramabadran, K.; Turndorf, H.; Shukla, V.K. Intracerebroventricular administration of kappa-agonists induces convulsions in mice. Brain Res. Bull. 1991, 27, 75–79. [Google Scholar] [CrossRef]
- Mahendran, R.; Lim, H.A.; Tan, J.Y.; Chua, S.M.; Winslow, M. Salvia divinorum: An overview of the usage, misuse, and addiction processes. Asia Pac. Psychiatry 2016, 8, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.E.; Panos, J.J.; Killinger, B.A.; Peet, M.M.; Bell, L.M.; Haliw, L.A.; Walker, S.L. Comparison of the discriminative stimulus effects of salvinorin A and its derivatives to U69,593 and U50,488 in rats. Psychopharmacology 2009, 203, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Pace, S.; Tedesco, F.; Pagano, E.; Guerra, G.; Troisi, F.; Werner, M.; Roviezzo, F.; Zjawiony, J.K.; Werz, O.; et al. The hallucinogenic diterpene salvinorin A inhibits leukotriene synthesis in experimental models of inflammation. Pharmacol. Res. 2016, 106, 64–71. [Google Scholar] [CrossRef]
- Kimball, A.W.; Burnett, W.T., Jr.; Doherty, D.G. Chemical protection against ionizing radiation. I. Sampling methods for screening compounds in radiation protection studies with mice. Radiat. Res. 1957, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Treatment | Neuromuscular Strength (mN/g) | Impairment of Motor Coordination (%) |
|---|---|---|
| control | 25.19 ± 3.34 | 0 |
| salvinorin A 0.1 mg/kg | 30.35 ± 6.49 | 0 |
| salvinorin A 1 mg/kg | 28.99 ± 7.27 | 0 |
| salvinorin A 10 mg/kg | 28.29 ± 7.15 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Socała, K.; Doboszewska, U.; Wlaź, P. Salvinorin A Does Not Affect Seizure Threshold in Mice. Molecules 2020, 25, 1204. https://doi.org/10.3390/molecules25051204
Socała K, Doboszewska U, Wlaź P. Salvinorin A Does Not Affect Seizure Threshold in Mice. Molecules. 2020; 25(5):1204. https://doi.org/10.3390/molecules25051204
Chicago/Turabian StyleSocała, Katarzyna, Urszula Doboszewska, and Piotr Wlaź. 2020. "Salvinorin A Does Not Affect Seizure Threshold in Mice" Molecules 25, no. 5: 1204. https://doi.org/10.3390/molecules25051204
APA StyleSocała, K., Doboszewska, U., & Wlaź, P. (2020). Salvinorin A Does Not Affect Seizure Threshold in Mice. Molecules, 25(5), 1204. https://doi.org/10.3390/molecules25051204