
Inhibition of Phytosterol Biosynthesis by Azasterols

 , ,
, ,  ,
, 
Abstract

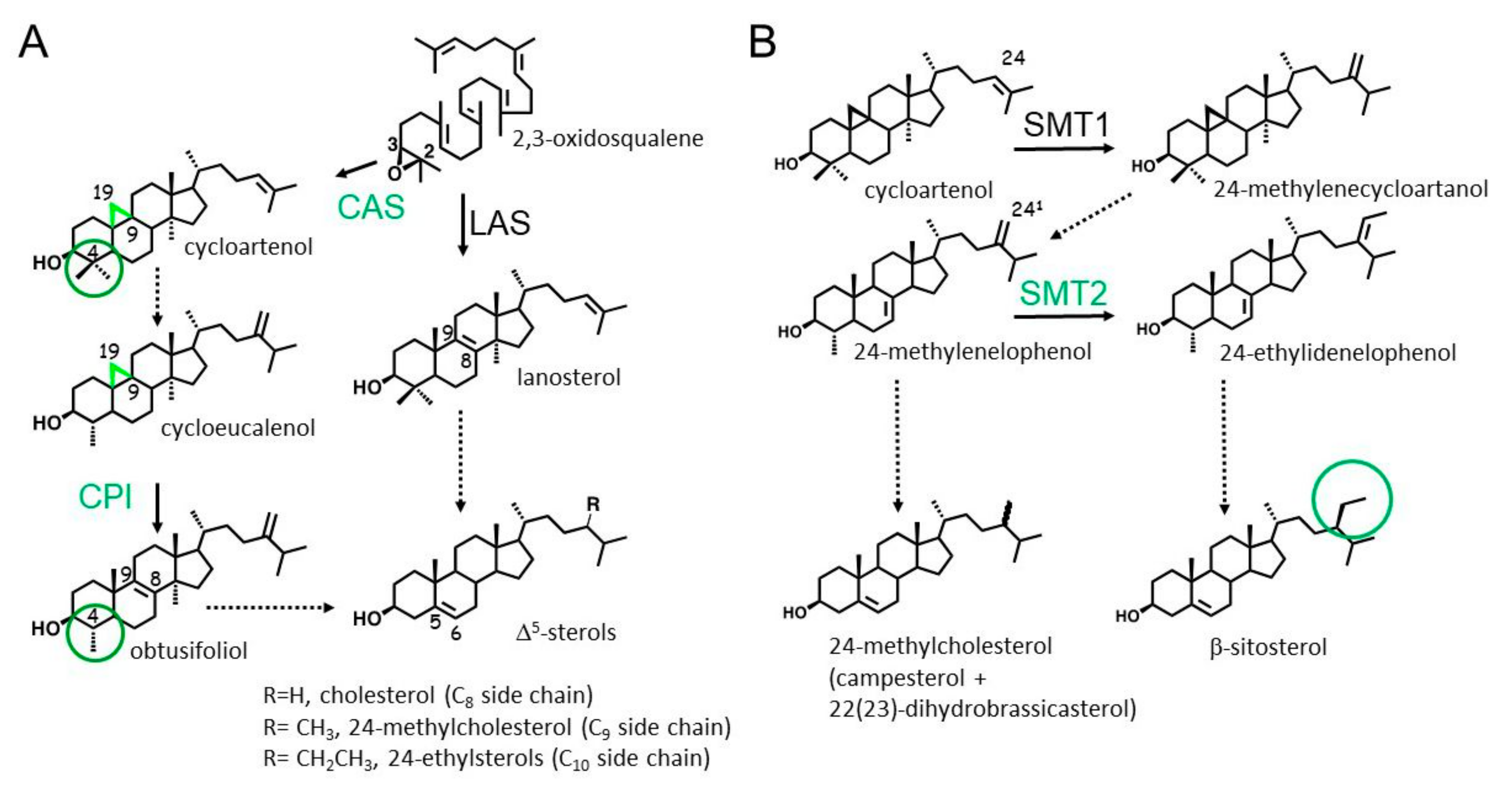
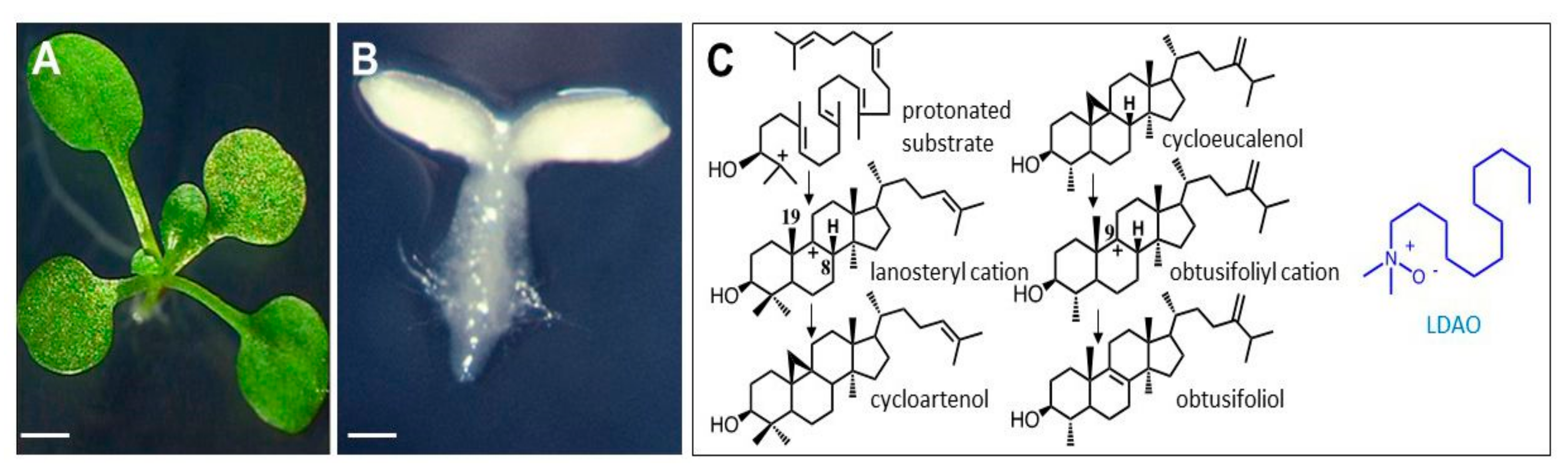
1. Introduction
2. Results
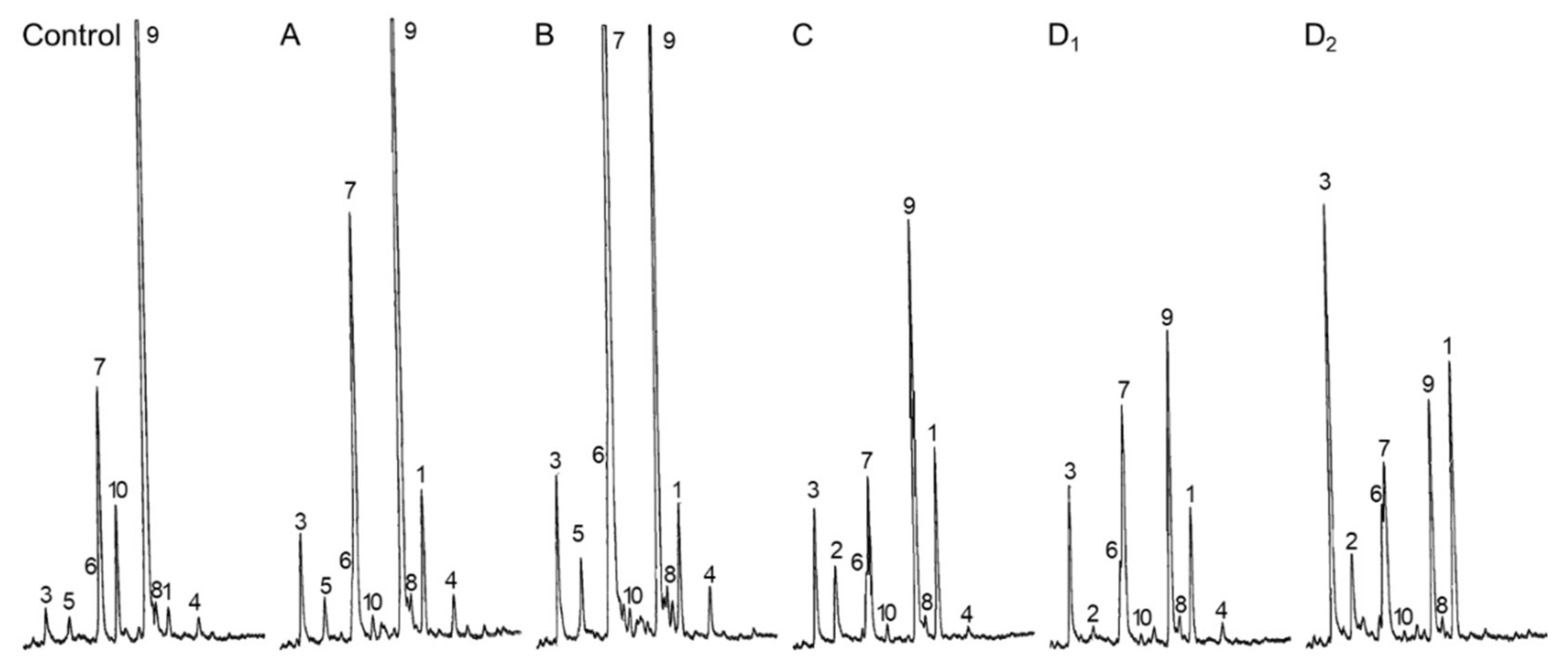
2.1. Multiple Target Sites of LDAO
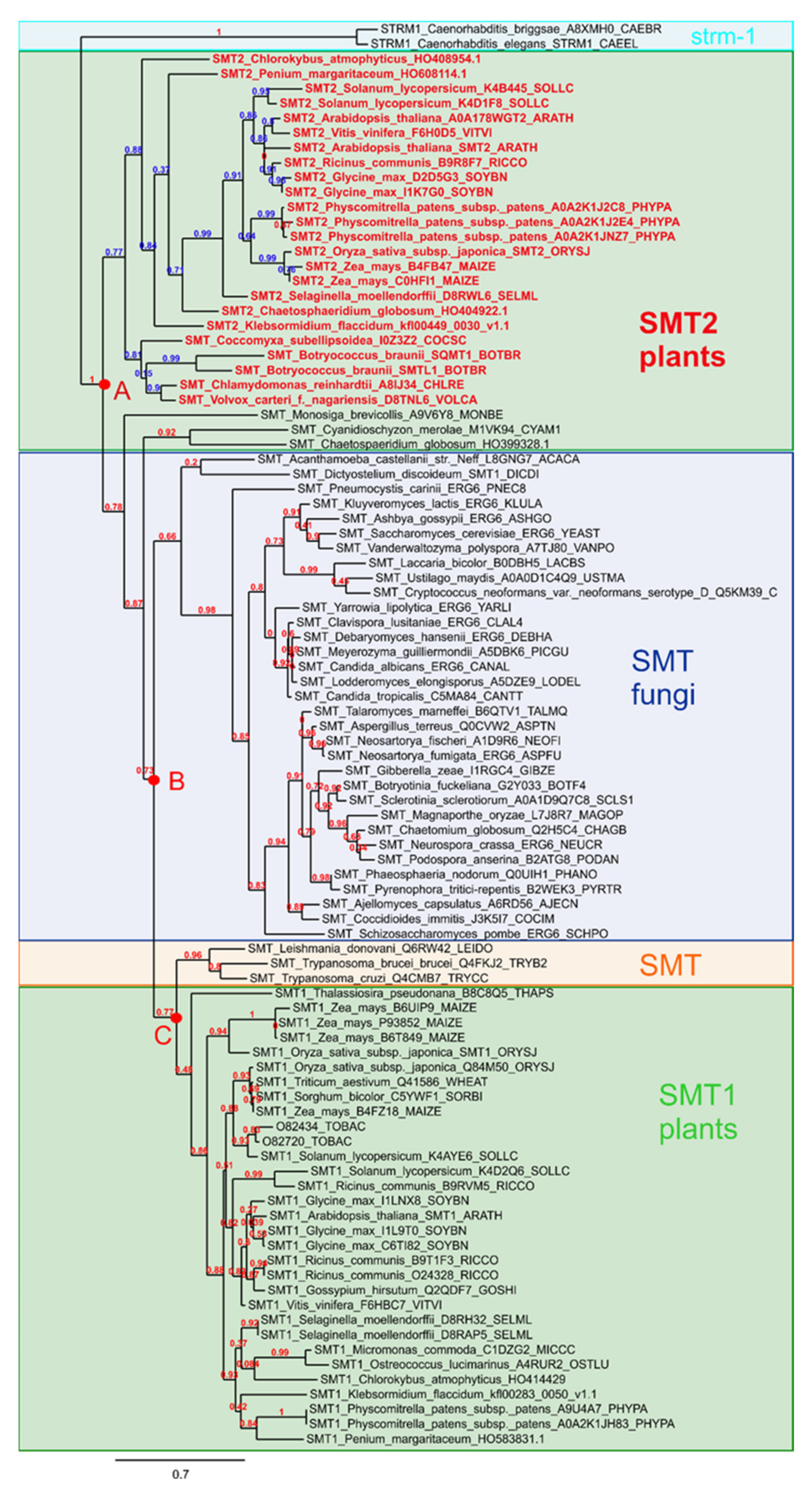
2.2. Selective Inhibition of SMTs
3. Discussion
4. Material and Methods
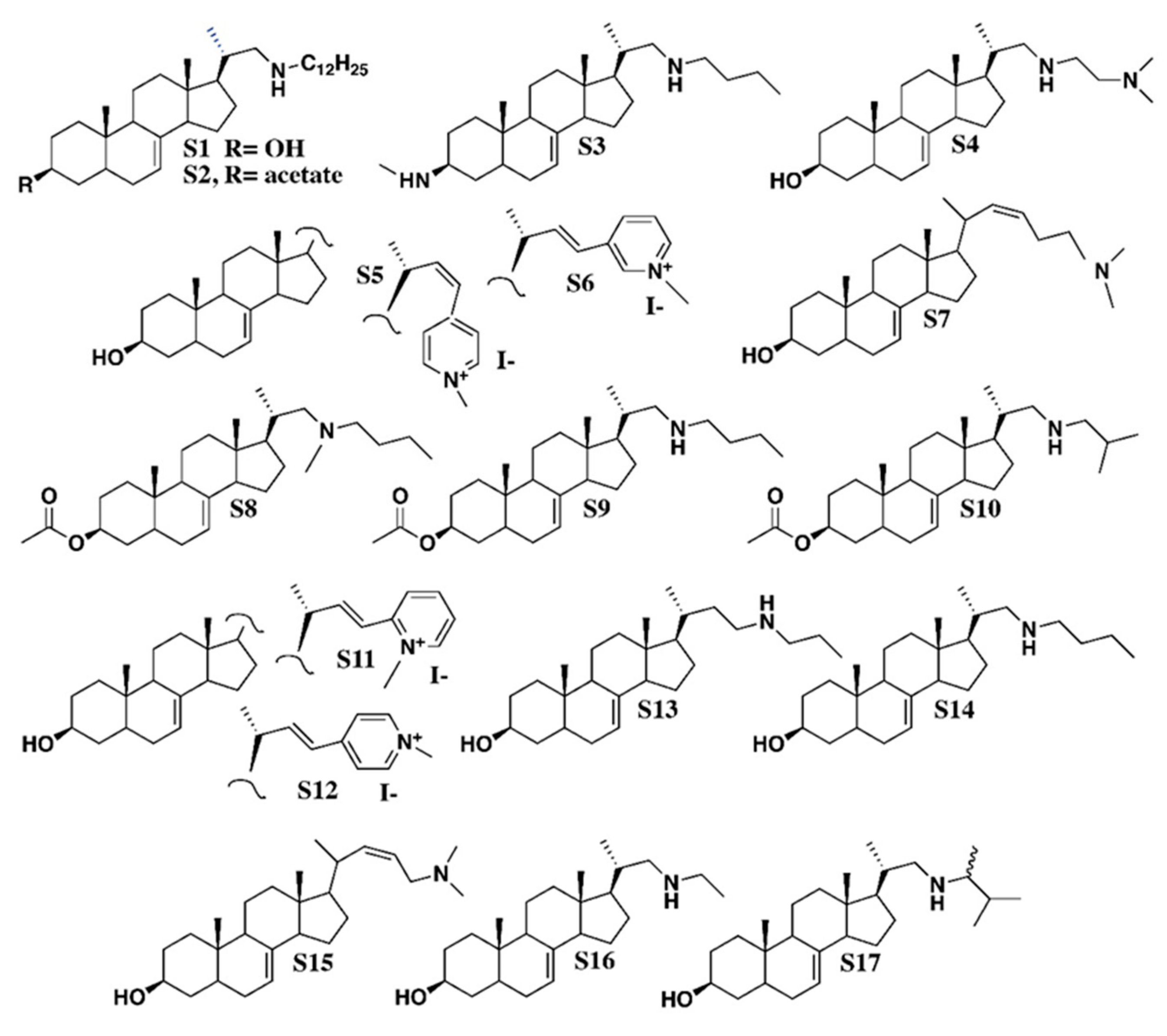
4.1. Chemicals
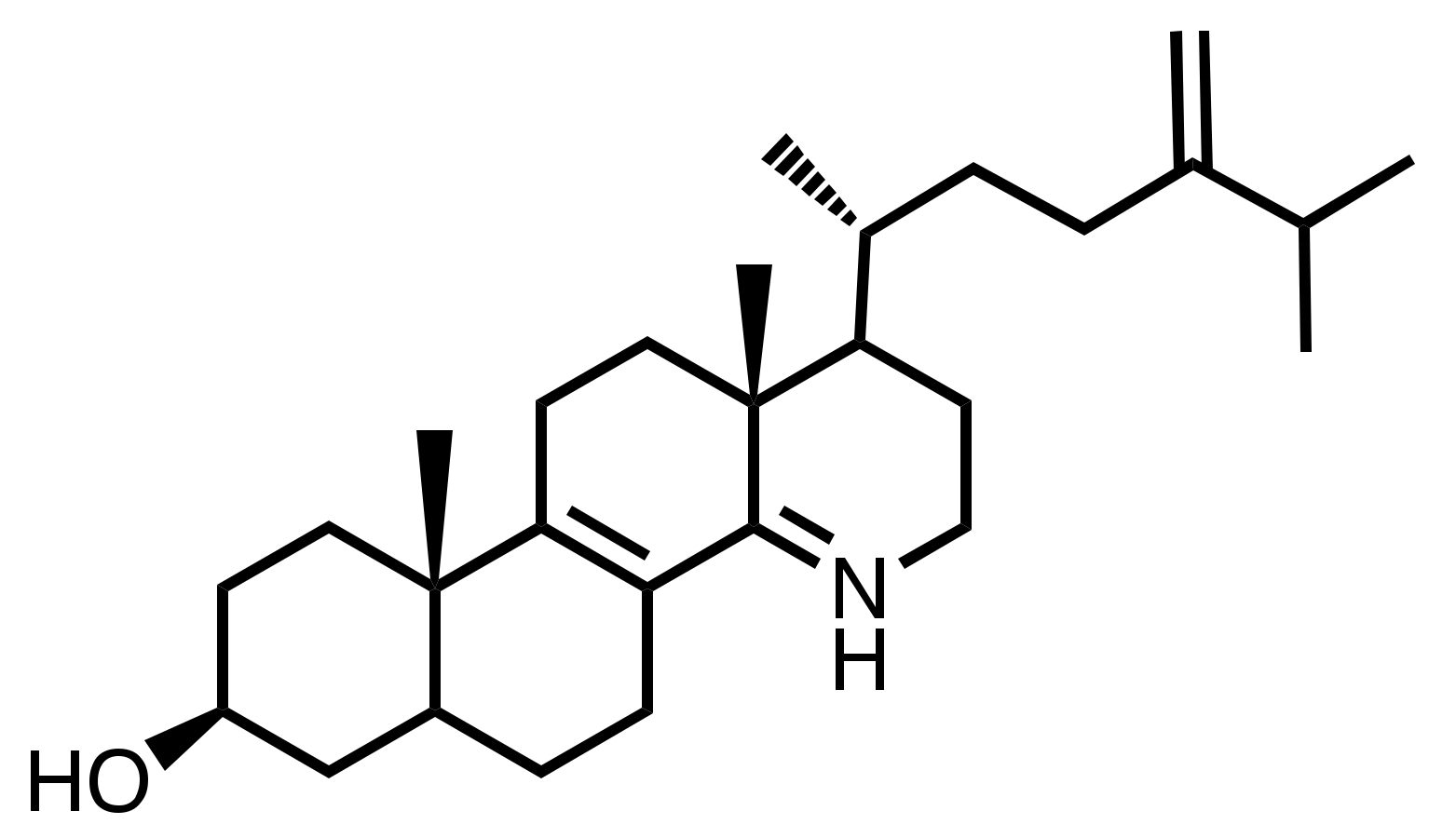
- S1, C34H61NO, CAS 1202777-29-4: Pregn-7-en-3-ol, 21-(dodecylamino)-20-methyl, (3β, 5α, 20S)-;
- S2, C36H63NO2, CAS 1202777-41-0: Pregn-7-en-3-ol, 21-(dodecylamino)-20-methyl, 3-acetate, (3β, 5α, 20S)-;
- S3, C27H48N2, Pregn-7-en-3-aminomethyl-21-(butylamino)-20-methyl-, (3β, 5α, 20S)-;
- S4, C26H46N2O, CAS 1202777-19-2: Pregn-7-en-3-ol, 21-[[2-(dimethylamino) ethyl] amino]-20-methyl, (3β, 5α, 20S)-;
- S5, C29H42NOI, Pyridinium, 4-[(3β, 5α, 22Z)-3-hydroxy-24-norchola-7,22-dien-23-yl]-1-methyl-iodide (1:1);
- S6, C29H42NOI, Pyridinium, 3-[(3β, 5α, 22E)-3-hydroxy-24-norchola-7,22-dien-23-yl]-1-methyl iodide (1:1);
- S7, C26H43NO, 26, 27-dinorcholesta-7,22-dien-3-ol, 25-(dimethylamino), (3β, 22Z)-;
- S8, C29H49NO2, CAS 1202777-10-3: Pregn-7-en-3-ol, 21-(butylmethylamino)-20-methyl-3-acetate, (3β, 5α, 20S)-;
- S9, C28H47NO2, CAS 1202777-37-4: Pregn-7-en-3-ol, 21-(butylamino)-20-methyl-3-acetate, (3β, 5α, 20S)-;
- S10, C28H47NO2, CAS 1202777-39-6: Pregn-7-en-3-ol, 20-methyl-21-[(2-methylpropylamino]-3acetate, (3β, 5α, 20S)-;
- S11, C29H42INO, CAS 1202777-30-7: Pyridinium, 2-[(3β, 5α, 22E)-3-hydroxy-24-norchola-7,22-dien-23-yl]-1-methyl-iodide (1:1);
- S12, C29H42NOI, CAS 1001324-61-3: Pyridinium, 4-[(3β, 5α, 22E)-3-hydroxy-24-norchola-7,22-dien-23-yl]-1-methyl-iodide (1:1);
- S13, C26H45NO, CAS 1202777-24-9: Pregn-7-en-3-ol, 20-methyl-21-(propylamino) (3β, 5α, 20S)-;
- S14, C26H45NO, CAS 1202777-25-0: Pregn-7-en-3-ol, 21-(butylamino)-20-methyl-(3β, 5α, 20S)-;
- S15, C25H41NO, chola-7,22-dienol-24-(dimethylamino) (3β, 5α, 22Z);
- S16, C24H41NO, CAS 1202777-23-8: Pregn-7-en-3-ol, 21-(ethylamino)-20-methyl-(3β, 5α, 20S)-;
- S17, C27H47NO, CAS 1202777-28-3: Pregn-7-en-3-ol, 21-[(1,2-dimethylpropyl)amino]-20-methyl-(3β, 5α, 20S)-.
4.2. Plant Treatment with Chemicals
4.3. Sterol Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Schaller, H. Sterol and Steroid Biosynthesis and Metabolism in Plants and Microorganisms. In Comprehensive Natural Products II: Chemistry and Biology, Volumes 1-10; Mander, L.N., Hung-Wen, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 132, pp. 755–787. [Google Scholar]
- Darnet, S.; Schaller, H. Metabolism and Biological Activities of 4-Methyl-Sterols. Molecules 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.J.; Matsuda, S.P.; Bartel, B. Molecular cloning, characterization, and overexpression of ERG7, the Saccharomyces cerevisiae gene encoding lanosterol synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.J.; Matsuda, S.P.; Bartel, B. Isolation of an Arabidopsis thaliana gene encoding cycloartenol synthase by functional expression in a yeast mutant lacking lanosterol synthase by the use of a chromatographic screen. Proc. Natl. Acad. Sci. USA 1993, 90, 11628–11632. [Google Scholar] [CrossRef]
- Baker, C.H.; Matsuda, S.P.; Liu, D.R.; Corey, E.J. Molecular cloning of the human gene encoding lanosterol synthase from a liver cDNA library. Biochem. Biophys. Res. Commun. 1995, 213, 154–160. [Google Scholar] [CrossRef]
- Ohyama, K.; Suzuki, M.; Kikuchi, J.; Saito, K.; Muranaka, T. Dual biosynthetic pathways to phytosterol via cycloartenol and lanosterol in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 725–730. [Google Scholar] [CrossRef]
- Forestier, E.; Romero-Segura, C.; Pateraki, I.; Centeno, E.; Compagnon, V.; Preiss, M.; Berna, A.; Boronat, A.; Bach, T.J.; Darnet, S.; et al. Distinct triterpene synthases in the laticifers of Euphorbia lathyris. Sci. Rep. 2019, 9, 4840. [Google Scholar] [CrossRef]
- Lovato, M.A.; Hart, E.A.; Segura, M.J.; Giner, J.L.; Matsuda, S.P. Functional cloning of an Arabidopsis thaliana cDNA encoding cycloeucalenol cycloisomerase. J. Biol. Chem. 2000, 275, 13394–13397. [Google Scholar] [CrossRef]
- Waterman, M.R.; Lepesheva, G.I. Sterol 14 alpha-demethylase, an abundant and essential mixed-function oxidase. Biochem. Biophys. Res. Commun. 2005, 338, 418–422. [Google Scholar] [CrossRef]
- Villette, C.; Berna, A.; Compagnon, V.; Schaller, H. Plant Sterol Diversity in Pollen from Angiosperms. Lipids 2015, 50, 749–760. [Google Scholar] [CrossRef]
- Darnet, S.; Rahier, A. Plant sterol biosynthesis: Identification of two distinct families of sterol 4α-methyl oxidases. Biochem. J. 2004, 378, 889–898. [Google Scholar] [CrossRef]
- Darnet, S.; Rahier, A. Enzymological properties of sterol-C4-methyl-oxidase of yeast sterol biosynthesis. Biochim. Biophys. Acta 2003, 1633, 106–117. [Google Scholar] [CrossRef]
- Nes, W.D. Sterol methyl transferase: Enzymology and inhibition. Biochim. Biophys. Acta 2000, 1529, 63–88. [Google Scholar] [CrossRef]
- Nakamoto, M.; Schmit, A.C.; Heintz, D.; Schaller, H.; Ohta, D. Diversification of sterol methyltransferase enzymes in plants and a role for beta-sitosterol in oriented cell plate formation and polarized growth. Plant. J. 2015, 84, 860–874. [Google Scholar] [CrossRef]
- Bouvier-Nave, P.; Husselstein, T.; Benveniste, P. Two families of sterol methyltransferases are involved in the first and the second methylation steps of plant sterol biosynthesis. Eur. J. Biochem. 1998, 256, 88–96. [Google Scholar] [CrossRef]
- Schaeffer, A.; Bronner, R.; Benveniste, P.; Schaller, H. The ratio of campesterol to sitosterol with modulates growth in Arabidopsis is controlled by STEROL METHYLTRANSFERASE 2-1. Plant. J. 2001, 25, 605–615. [Google Scholar] [CrossRef]
- Darnet, S.; Schaller, H. Unleashed sterol production in thale cress. Nat. Plants 2019, 5, 1112–1113. [Google Scholar] [CrossRef]
- Rubinstein, I.; Goad, L.J.; Clague, A.D.H.; Mulheirn, L.J. The 220 MHz NMR spectra of phytosterols. Phytochemistry 1976, 15, 195–200. [Google Scholar] [CrossRef]
- Rendeli, N.; Misso, N.L.; Goad, L.J. Biosynthesis of 24-methylcholest-5-en-3beta-ol and 24-ethylcholest-5-en-3beta-ol in Zea mays. Lipids 1986, 21, 63–68. [Google Scholar] [CrossRef]
- Moss, G.P. Nomenclature of steroids (Recommendations 1989). Pure Appl. Chem. 1989, 61, 1783–1822. [Google Scholar] [CrossRef]
- Specialist Technical Group of CropLife International Fungicide Resistance Management by Fungicide Common Name. Available online: https://www.frac.info/fungicide-resistance-management/by-fungicide-common-name (accessed on 3 February 2019).
- Muller, C.; Junker, J.; Bracher, F.; Giera, M. A gas chromatography-mass spectrometry-based whole-cell screening assay for target identification in distal cholesterol biosynthesis. Nat. Protoc. 2019, 14, 2546–2570. [Google Scholar] [CrossRef]
- Price, C.L.; Parker, J.E.; Warrilow, A.G.; Kelly, D.E.; Kelly, S.L. Azole fungicides-understanding resistance mechanisms in agricultural fungal pathogens. Pest. Manag. Sci. 2015, 71, 1054–1058. [Google Scholar] [CrossRef]
- Leaver, D.J.; Patkar, P.; Singha, U.K.; Miller, M.B.; Haubrich, B.A.; Chaudhuri, M.; Nes, W.D. Fluorinated Sterols Are Suicide Inhibitors of Ergosterol Biosynthesis and Growth in Trypanosoma brucei. Chem. Biol. 2015, 22, 1374–1383. [Google Scholar] [CrossRef]
- Franz, J.; Schwinn, F.J. Ergosterol Biosynthesis Inhibitors. An Overview of Their History and Contribution to Medicine and Agriculture. Pestic. Sci. 1983, 15, 4W7. [Google Scholar]
- Lepesheva, G.I.; Friggeri, L.; Waterman, M.R. CYP51 as drug targets for fungi and protozoan parasites: Past, present and future. Parasitology 2018, 145, 1820–1836. [Google Scholar] [CrossRef]
- Muller, C.; Binder, U.; Maurer, E.; Grimm, C.; Giera, M.; Bracher, F. Fungal sterol C22-desaturase is not an antimycotic target as shown by selective inhibitors and testing on clinical isolates. Steroids 2015, 101, 1–6. [Google Scholar] [CrossRef]
- Helliwell, S.B.; Karkare, S.; Bergdoll, M.; Rahier, A.; Leighton-Davis, J.R.; Fioretto, C.; Aust, T.; Filipuzzi, I.; Frederiksen, M.; Gounarides, J.; et al. FR171456 is a specific inhibitor of mammalian NSDHL and yeast Erg26p. Nat. Commun. 2015, 6, 8613. [Google Scholar] [CrossRef]
- Kuchta, T.; Léka, C.; Kubinec, R.; Russell, N.J. Inhibition of ergosterol biosynthesis is not accompanied by a change in fatty acid composition in Saccharomyces cerevisiae treated with the antifungal agent 6-amino-2-n-pentylthiobenzothiazole. FEMS Microbiol. Lett. 2006, 150, 43–47. [Google Scholar] [CrossRef]
- Benveniste, P. Sterol biosynthesis. Ann. Rev. Plant. Physiology. 1986, 37, 275–307. [Google Scholar] [CrossRef]
- Rahier, A.; Bouvier, P.; Cattel, L.; Narula, A.; Benveniste, P. Inhibition of 2,3-oxidosqualene: Beta-amyrin-cyclase, S-adenosyl-L-methionine: Cycloartenol C-24-methyltransferase and cycloeucalenol: Obtusifoliol isomerase by rationally designed molecules containing a tertiary amine function. Biochem. Soc. Trans. 1983, 11, 537–543. [Google Scholar] [CrossRef]
- Rahier, A.; Taton, M. Fungicides as tools in studying postsqualene sterol synthesis in plants. Pestic. Biochem. Physiol. 1997, 57, 1–27. [Google Scholar] [CrossRef]
- Rahier, A.; Narula, A.S.; Benveniste, P.; Schmitt, P. 25-Azacycloartanol, a potent inhibitor of S-adenosyl-L-methionine-sterol-C-24 and C-28 methyltransferases in higher plant cells. Biochem. Biophys. Res. Commun. 1980, 92, 20–25. [Google Scholar] [CrossRef]
- Cerutti, M.; Delprino, L.; Cattel, L.; Bouviernave, P.; Duriatti, A.; Schuber, F.; Benveniste, P. N-Oxide as a Potential Function in the Design of Enzyme-Inhibitors-Application to 2,3-Epoxysqualene-Sterol Cyclases. J. Chem. Soc. Chem. Commun. 1985, 1054–1055. [Google Scholar] [CrossRef]
- Kushiro, T.; Ebizuka, Y. Triterpenes. In Comprehensive Natural Products II; Liu, H.-W., Mander, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 1, pp. 673–708. [Google Scholar]
- Rahier, A.; Taton, M.; Benveniste, P. Cycloeucalenol-obtusifoliol isomerase. Structural requirements for transformation or binding of substrates and inhibitors. Eur. J. Biochem. 1989, 181, 615–626. [Google Scholar] [CrossRef]
- Jachak, G.R.; Ramesh, R.; Sant, D.G.; Jorwekar, S.U.; Jadhav, M.R.; Tupe, S.G.; Deshpande, M.V.; Reddy, D.S. Silicon Incorporated Morpholine Antifungals: Design, Synthesis, and Biological Evaluation. ACS Med. Chem. Lett. 2015, 6, 1111–1116. [Google Scholar] [CrossRef]
- Asami, T.; Mizutani, M.; Shimada, Y.; Goda, H.; Kitahata, N.; Sekimata, K.; Han, S.Y.; Fujioka, S.; Takatsuto, S.; Sakata, K.; et al. Triadimefon, a fungicidal triazole-type P450 inhibitor, induces brassinosteroid deficiency-like phenotypes in plants and binds to DWF4 protein in the brassinosteroid biosynthesis pathway. Biochem. J. 2003, 369, 71–76. [Google Scholar] [CrossRef]
- Renard, D.; Perruchon, J.; Giera, M.; Muller, J.; Bracher, F. Side chain azasteroids and thiasteroids as sterol methyltransferase inhibitors in ergosterol biosynthesis. Bioorg. Med. Chem. 2009, 17, 8123–8137. [Google Scholar] [CrossRef]
- Diener, A.C.; Li, H.; Zhou, W.; Whoriskey, W.J.; Nes, W.D.; Fink, G.R. Sterol methyltransferase 1 controls the level of cholesterol in plants. Plant. Cell 2000, 12, 853–870. [Google Scholar] [CrossRef]
- Nes, W.D.; Zhou, W.; Ganapathy, K.; Liu, J.; Vatsyayan, R.; Chamala, S.; Hernandez, K.; Miranda, M. Sterol 24-C-methyltransferase: An enzymatic target for the disruption of ergosterol biosynthesis and homeostasis in Cryptococcus neoformans. Arch. Biochem. Biophys. 2009, 481, 210–218. [Google Scholar] [CrossRef]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef]
- Schaller, H.; Bouvier-Navé, P.; Benveniste, P. Overexpression of an Arabidopsis cDNA Encoding a Sterol-C241-Methyltransferase in Tobacco Modifies the Ratio of 24-Methyl Cholesterol to Sitosterol and Is Associated with Growth Reduction. Plant. Physiol. 1998, 118, 461–469. [Google Scholar] [CrossRef]
- Haubrich, B.A.; Collins, E.K.; Howard, A.L.; Wang, Q.; Snell, W.J.; Miller, M.B.; Thomas, C.D.; Pleasant, S.K.; Nes, W.D. Characterization, mutagenesis and mechanistic analysis of an ancient algal sterol C24-methyltransferase: Implications for understanding sterol evolution in the green lineage. Phytochemistry 2015, 113, 64–72. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; Parks, L.W. Growth and antifungal homoazasterol production in Geotrichum flavo-brunneum. Antimicrob. Agents Chemother. 1980, 18, 822–828. [Google Scholar] [CrossRef]
- Taton, M.; Benveniste, P.; Rahier, A. Microsomal Delta8,14-sterol Delta14-reductase in higher plants. Characterization and inhibition by analogues of a presumptive carbocationic intermediate of the reduction reaction. Eur. J. Biochem. 1989, 185, 605–614. [Google Scholar] [CrossRef]
- Kidane, M.E.; Vanderloop, B.H.; Zhou, W.; Thomas, C.D.; Ramos, E.; Singha, U.; Chaudhuri, M.; Nes, W.D. Sterol methyltransferase a target for anti-amoeba therapy: Towards transition state analog and suicide substrate drug design. J. Lipid Res. 2017, 58, 2310–2323. [Google Scholar] [CrossRef]
- Zhou, W.; Ramos, E.; Zhu, X.; Fisher, P.M.; Kidane, M.E.; Vanderloop, B.H.; Thomas, C.D.; Yan, J.; Singha, U.; Chaudhuri, M.; et al. Steroidal antibiotics are antimetabolites of Acanthamoeba steroidogenesis with phylogenetic implications. J. Lipid Res. 2019, 60, 981–994. [Google Scholar] [CrossRef]
- Miller, M.B.; Patkar, P.; Singha, U.K.; Chaudhuri, M.; David Nes, W. 24-Methylenecyclopropane steroidal inhibitors: A Trojan horse in ergosterol biosynthesis that prevents growth of Trypanosoma brucei. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2017, 1862, 305–313. [Google Scholar] [CrossRef]
- Gas-Pascual, E.; Berna, A.; Bach, T.J.; Schaller, H. Plant oxidosqualene metabolism: Cycloartenol synthase-dependent sterol biosynthesis in Nicotiana benthamiana. PLoS ONE 2014, 9, e109156. [Google Scholar] [CrossRef]
- Babiychuk, E.; Bouvier-Nave, P.; Compagnon, V.; Suzuki, M.; Muranaka, T.; Van Montagu, M.; Kushnir, S.; Schaller, H. Allelic mutant series reveal distinct functions for Arabidopsis cycloartenol synthase 1 in cell viability and plastid biogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 3163–3168. [Google Scholar] [CrossRef]
- Rahier, A.; Benveniste, P. Mass spectral identification of phytosterols. In Analysis of sterols and other significant steroids; Nes, W.D., Parish, E., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 223–250. [Google Scholar]
Sample Availability: Samples of the compounds named S1 to S17 are available upon reasonable request to F.B. Department of Pharmacy, Center for Drug Research, Ludwig-Maximilians University Munich. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LDAO mg·L−1 | 0 | 10 | 20 |
|---|---|---|---|
| 2,3-Oxidosqualene in mg·g−1 dry weight | tr | 0.2 | 1.4 |
| Sterols% | |||
| Cycloartenol | 1.0 | Nd | Nd |
| 24-Methylenecycloartanol | 2.0 | 2.0 | 5.0 |
| Cycloeucalenol | 0.3 | 48.0 | 51.0 |
| Cyclofontumienol | nd | 3.0 | 2.0 |
| 24-Methylenepollinastanol | nd | 27.0 | 25.2 |
| 24-Methylpollinastanol | nd | 1.0 | Nd |
| Total 9β,19-cyclopropylsterols | 3.3 | 81.0 | 83.2 |
| Cholesterol | 4.5 | 0.3 | Nd |
| 24-Methylenecholesterol | 1.2 | Nd | Nd |
| Campesterol | 13.5 | 1.0 | 0.8 |
| Isofucosterol | 2.3 | 1.0 | 1.2 |
| Sitosterol | 69.5 | 9.5 | 9.5 |
| Stigmasterol | 3.0 | 1.0 | 1.3 |
| Total 24-alkyl-5-sterols | 94.0 | 12.8 | 12.8 |
| Other minor sterols | 2.7 | 6.2 | 4.0 |
| Total sterols in mg·g−1 dry weight | 2.2 | 1.2 | 1.7 |
| Group A | Group B | Group C | Group D | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sterols % | control | S6 | S8 | S16 | S17 | S11 | S13 | S14 | |
| Cycloartenol (1) | 1.0 | 14.0 | 5.1 | 4.5 | 3.0 | 16.3 | 20.3 | 15.0 | |
| Desmosterol (2) | nd | 1.8 | Nd | Nd | nd | 7.5 | 5.3 | 1.0 | |
| Cholesterol (3) | 2.0 | 8.5 | 4.0 | 5.3 | 1.8 | 10.5 | 28.7 | 14.5 | |
| Total C8 side chain sterols | 3.0 | 24.3 | 9.1 | 9.8 | 4.8 | 34.3 | 54.3 | 30.5 | |
| 24-Methylenecycloartanol (4) | 0.6 | 0.5 | 1.4 | 1.3 | 1.2 | nd | Nd | 1.0 | |
| Brassicasterol (5) | 1.1 | nd | 2.3 | 2.3 | 2.4 | nd | 0.9 | Nd | |
| 24-Methylenecholesterol (6) | nd | 3.7 | * | * | * | 7.0 | 8.7 | 7.5 | |
| 24-Methylcholesterol (7) | 14.5 | 21.2 | 27.1 | 59.0 | 42.4 | 15.0 | 16.0 | 21.5 | |
| Total C9 side chain sterols | 16.2 | 25.4 | 30.8 | 62.6 | 46.0 | 22 | 25.6 | 30.0 | |
| Isofucosterol (8) | 0.9 | 1.4 | 0.8 | 0.6 | 0.5 | 1.5 | 0.9 | 0.5 | |
| Sitosterol (9) | 73.9 | 48.0 | 58.1 | 26.5 | 47.8 | 40.8 | 18.2 | 38.5 | |
| Stigmasterol (10) | 6.0 | 0.9 | 1.2 | 0.5 | 0.9 | 1.4 | 1.0 | 0.5 | |
| Total C10 side chain sterols | 80.8 | 50.3 | 60.1 | 27.6 | 49.2 | 43.7 | 20.1 | 39.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darnet, S.; B B Martin, L.; Mercier, P.; Bracher, F.; Geoffroy, P.; Schaller, H. Inhibition of Phytosterol Biosynthesis by Azasterols. Molecules 2020, 25, 1111. https://doi.org/10.3390/molecules25051111
Darnet S, B B Martin L, Mercier P, Bracher F, Geoffroy P, Schaller H. Inhibition of Phytosterol Biosynthesis by Azasterols. Molecules. 2020; 25(5):1111. https://doi.org/10.3390/molecules25051111
Chicago/Turabian StyleDarnet, Sylvain, Laetitia B B Martin, Pierre Mercier, Franz Bracher, Philippe Geoffroy, and Hubert Schaller. 2020. "Inhibition of Phytosterol Biosynthesis by Azasterols" Molecules 25, no. 5: 1111. https://doi.org/10.3390/molecules25051111
APA StyleDarnet, S., B B Martin, L., Mercier, P., Bracher, F., Geoffroy, P., & Schaller, H. (2020). Inhibition of Phytosterol Biosynthesis by Azasterols. Molecules, 25(5), 1111. https://doi.org/10.3390/molecules25051111