
Novel Chaperones RrGroEL and RrGroES for Activity and Stability Enhancement of Nitrilase in Escherichia coli and Rhodococcus ruber

Abstract
1. Introduction
2. Results
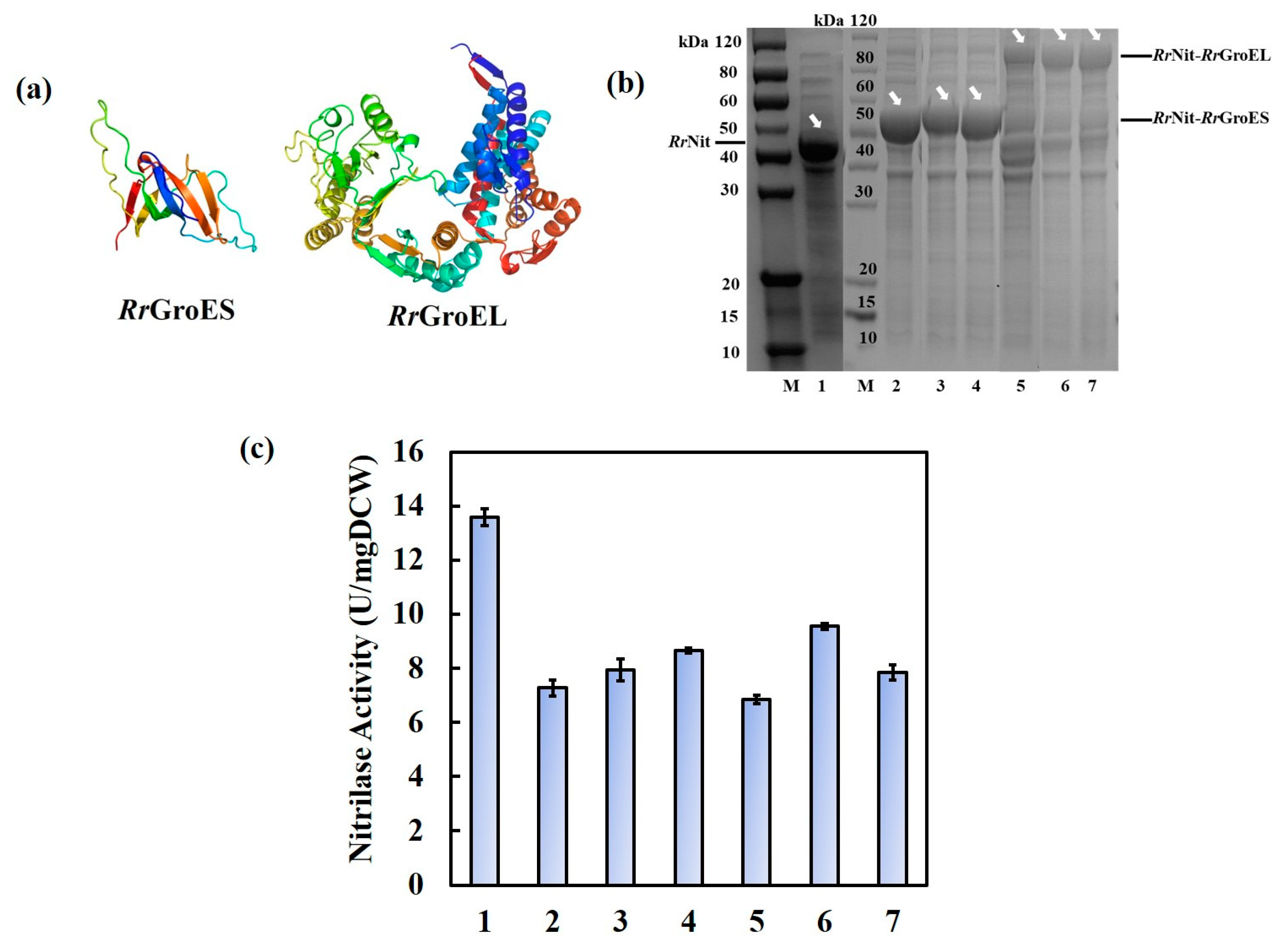
2.1. Fusion Expression of Either RrGroES or RrGroEL with RrNit Can Enhance the Enzyme Thermal Stability in E. coli
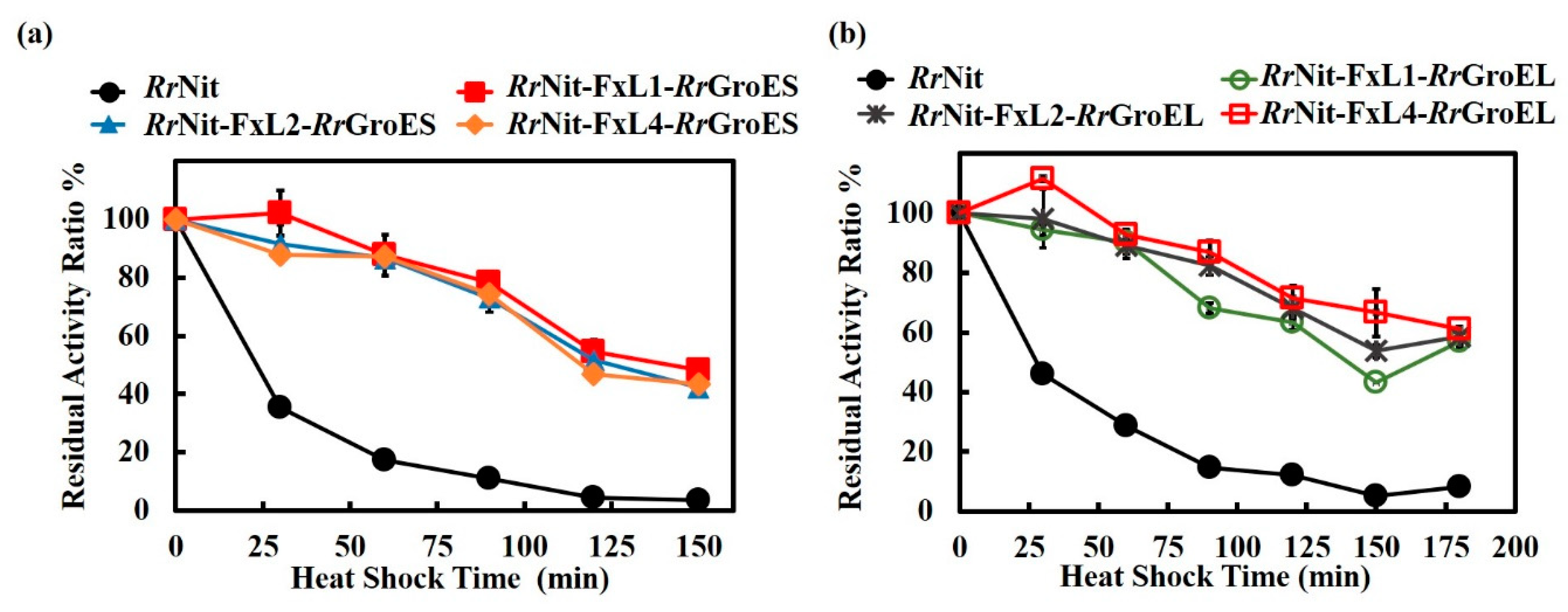
2.2. Effect of Linker Length on the Thermal Stability of RrNit-RrGroES/RrGroEL Fusion Chimeras in E. coli
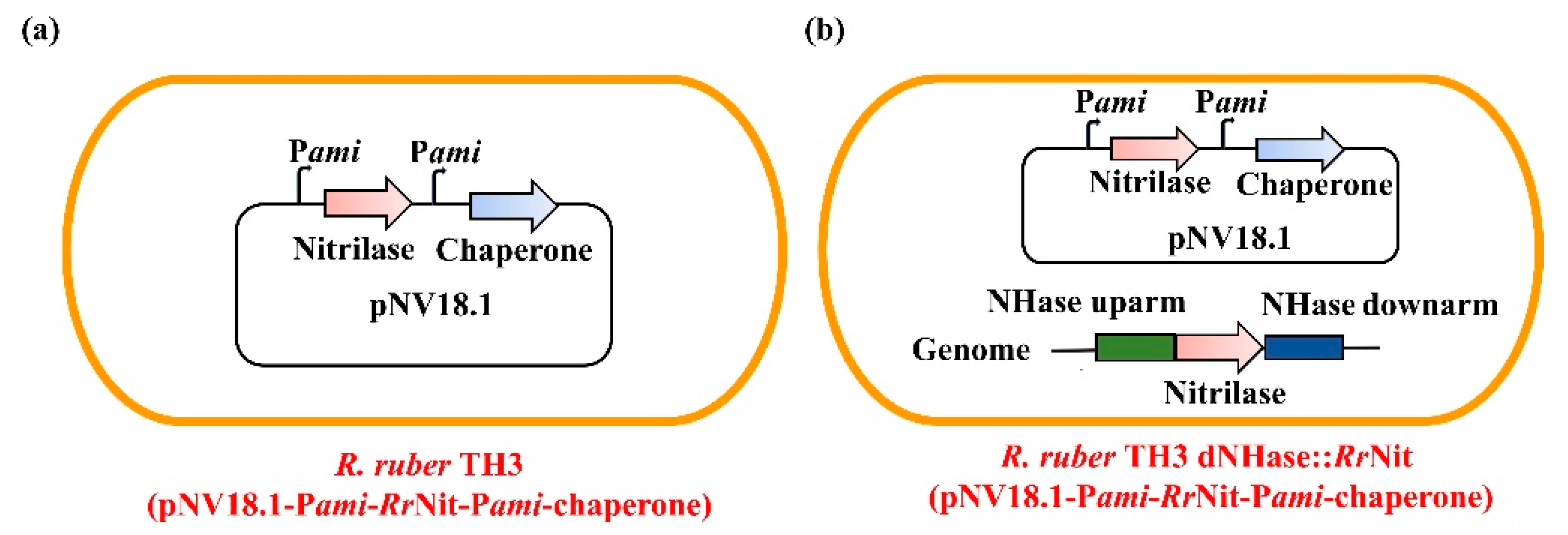
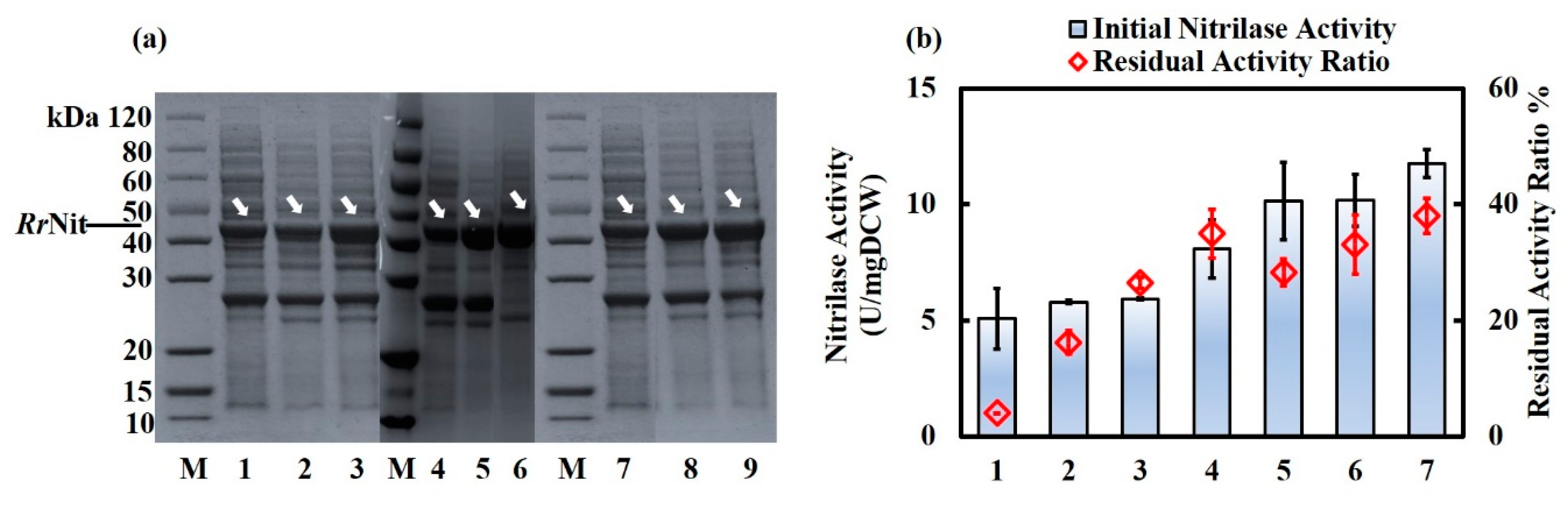
2.3. Fusion- and Co-expression of RrNit and RrGroES/RrGroEL in Parental R. ruber TH3
3. Discussion
4. Materials and Methods
4.1. Plasmids, Strains and Chemicals
4.2. Construction of the Recombinant Strains
4.3. Culture Conditions for the Recombinant Strains
4.4. Assay of Nitrilase Activity
4.5. In-cell Nitrilase Stability Evaluation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pace, H.C.; Brenner, C. The nitrilase superfamily: Classification, structure and function. Genome Biol. 2001, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yin, B.; Wang, C.; Jiang, S.; Wang, H.; Yuan, Y.A.; Wei, D. Structural insights into enzymatic activity and substrate specificity determination by a single amino acid in nitrilase from Syechocystis sp. PCC6803. J. Struct. Biol. 2014, 188, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Ma, J.; Chang, Y.; Yu, H.; Shen, Z. Directed evolution and mutant characterization of nitrilase from Rhodococcus rhodochrous tg1-A6. Appl. Biochem. Biotechnol. 2016, 178, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, J.; Jiang, S.Q.; Wei, D.Z. Recent research advancements on regioselective nitrilase: Fundamental and applicative aspects. Appl. Microbiol. Biotechnol. 2019, 103, 6393–6405. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Sharma, R.; Tewari, N.; Rawat, D.S. Nitrilase and its application as a ‘green’ catalyst. Chem. Biodivers. 2006, 3, 1279–1287. [Google Scholar] [CrossRef]
- Luo, H.; Fan, L.; Chang, Y.; Ma, J.; Yu, H.; Shen, Z. Gene cloning, overexpression, and characterization of the nitrilase from Rhodococcus rhodochrous tg1-A6 in E. coli. Appl. Biochem. Biotechnol. 2010, 160, 393–400. [Google Scholar] [CrossRef]
- Luo, H.; Wang, T.; HuiMin, Y.; Yang, H.; Shen, Z. Expression and catalyzing process of the nirilase in Rhodococcus rhodochrous tg1-A6. Modern. Chem. Ind. 2006, 26, 109–113. [Google Scholar]
- Cowan, D.; Cramp, R.; Pereira, R.; Graham, D.; Almatawah, Q. Biochemistry and biotechnology of mesophilic and thermophilic nitrile metabolizing enzymes. Extremophiles 1998, 2, 207–216. [Google Scholar] [CrossRef]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Xu, Z.; Cai, T.; Xiong, N.; Zou, S.P.; Xue, Y.P.; Zheng, Y.G. Engineering the residues on “A” surface and C-terminal region to improve thermostability of nitrilase. Enzyme Microb. Technol. 2018, 113, 52–58. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.; Xue, Y.; Zheng, Y. Activity improvement of a regioselective nitrilase from Acidovorax facilis and its application in the production of 1-(cyanocyclohexyl) acetic acid. Process. Biochem. 2014, 49, 2141–2148. [Google Scholar] [CrossRef]
- DeSantis, G.; Wong, K.; Farwell, B.; Chatman, K.; Zhu, Z.L.; Tomlinson, G.; Huang, H.J.; Tan, X.Q.; Bibbs, L.; Chen, P.; et al. Creation of a productive, highly enantioselective nitrilase through gene site saturation mutagenesis (GSSM). J. Am. Chem. Soc. 2003, 125, 11476–11477. [Google Scholar] [CrossRef] [PubMed]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiao, S.; Wang, M.; Chen, J.; Yu, H. A novel molecular chaperone GroEL2 from Rhodococcus ruber and its fusion chimera with nitrile hydratase for co-enhanced activity and stability. Chem. Eng. Sci. 2018, 192, 235–243. [Google Scholar] [CrossRef]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef]
- Nishihara, K.; Kanemori, M.; Kitagawa, M.; Yanagi, H.; Yura, T. Chaperone coexpression plasmids: Differential and synergistic roles of DnaK-DnaJ-GrpE and GroEL-GroES in assisting folding of an allergen of Japanese cedar pollen, Cryj2 in Escherichia coli. Appl. Environ. Microbiol. 1998, 64, 1694–1699. [Google Scholar] [CrossRef]
- Tian, Y.; Chen, J.; Yu, H.; Shen, Z. Overproduction of the Escherichia coli chaperones GroEL-GroES in Rhodococcus ruber improves the activity and stability of cell catalysts harboring a nitrile hydratase. J. Microbiol. Biotechnol. 2016, 26, 337–346. [Google Scholar] [CrossRef]
- Petrickova, A.; Vesela, A.B.; Kaplan, O.; Kubac, D.; Uhnakova, B.; Malandra, A.; Felsberg, J.; Rinagelova, A.; Weyrauch, P.; Kren, V.; et al. Purification and characterization of heterologously expressed nitrilases from filamentous fungi. Appl. Microbiol. Biotechnol. 2012, 93, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Hu, S.; Guan, Y.X.; Yao, S.J. Coexpression of chaperonin GroEL/GroES markedly enhanced soluble and functional expression of recombinant human interferon-gamma in Escherichia coli. Appl. Microbiol. Biotechnol. 2012, 93, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Hartl, F.U.; Martin, J. Molecular chaperones in cellular protein folding. Curr. Opin. Struct. Biol. 1996, 381, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Langer, T.; Pfeifer, G.; Martin, J.; Baumeister, W.; Hartl, F.U. Chaperonin-mediated protein folding: GroES binds to one end of the GroEL cylinder, which accommodates the protein substrate within its central cavity. EMBO J. 1992, 11, 4757–4765. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.S.; Philp, J.C.; Aw, D.W.J.; Christofi, N. A review - The genus Rhodococcus. J. Appl. Microbiol. 1998, 85, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Jiao, S.; Wang, M.; Yu, H.; Shen, Z. A CRISPR/Cas9-based genome editing system for Rhodococcus ruber TH. Metab. Eng. 2020, 57, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, J.; Yu, H.; Shen, Z. Improving stress tolerance and cell integrity of Rhodococcus ruber by overexpressing small-shock-protein Hsp16 of Rhodococcus. J. Ind. Microbiol. Biotechnol. 2018, 45, 929–938. [Google Scholar] [CrossRef]
- Qamra, R.; Srinivas, V.; Mande, S. Mycobacterium tuberculosis GroEL homologues unusually exist as lower oligomers and retain the ability to suppress aggregation of substrate proteins. J. Mol. Biol. 2004, 342, 605–617. [Google Scholar] [CrossRef]
- Sun, J.; Yu, H.; Chen, J.; Luo, H.; Shen, Z. Ammonium acrylate biomanufacturing by an engineered Rhodococcus ruber with nitrilase overexpression and double-knockout of nitrile hydratase and amidase. J. Ind. Microbiol. Biotechnol. 2016, 43, 1631–1639. [Google Scholar] [CrossRef]
- Bergeron, L.M.; Lee, C.; Tokatlian, T.; Hoellrigl, V.; Clark, D.S. Chaperone function in organic co-solvents: Experimental characterization and modeling of a hyperthermophilic chaperone subunit from Methanocaldococcus jannaschii. BBA-Proteins Proteomics 2008, 1784, 368–378. [Google Scholar] [CrossRef]
- Zhao, H.L.; Yao, X.Q.; Xue, C.; Wang, Y.; Xiong, X.H.; Liu, Z.M. Increasing the homogeneity, stability and activity of human serum albumin and interferon-α2b fusion protein by linker engineering. Protein Expr. Purif. 2008, 61, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Yu, H.; Shen, Z. Core element characterization of Rhodococcus promoters and development of a promoter-RBS mini-pool with different activity levels for efficient gene expression. New Biotechnol. 2018, 44, 41–49. [Google Scholar] [CrossRef]
- Volker, U.; Mach, H.; Schmid, R.; Hecker, H. Stress proteins and cross-protection by heat shock and salt stress in Bacillus subtilis. J. Gen. Microbiol. 1992, 138, 2125. [Google Scholar] [CrossRef]
- Taguchi, H.; Konishi, J.; Ishii, N.; Yoshida, M. A chaperonin from a thermophilic bacterium, Thermus thermophilus, that controls refoldings of several thermophilic enzymes. J. Biol. Chem. 1991, 266, 22411–22418. [Google Scholar]
- Parsell, D.A.; Kowal, A.S.; Singer, M.A.; Lindquist, S. Protein disaggregation mediated by heat-shock protein Hsp104. Nature 1994, 372, 475–478. [Google Scholar] [CrossRef]
- Martin, J.; Horwich, A.L.; Hartl, F.U. Prevention of protein denaturation under heat-stress by the chaperonin Hsp60. Science 1992, 258, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Sigler, P.B.; Xu, Z.; Rye, H.S.; Burston, S.G.; Fenton, W.A.; Horwich, A.L. Structure and function in GroEL-mediated protein folding. Annu. Rev. Biochem. 1998, 67, 581–608. [Google Scholar] [CrossRef] [PubMed]
- Almatawah, Q.A.; Cramp, R.; Cowan, D.A. Characterization of an inducible nitrilase from a thermophilic Bacillus. Extremophiles 1999, 3, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- Georis, J.; Esteves, F.D.; Lamotte-Brasseur, J.; Bougnet, V.; Devreese, B.; Giannotta, F.; Granier, B.; Frere, J.M. An additional aromatic interaction improves the thermostability and thermophilicity of a mesophilic family 11 xylanase: Structural basis and molecular study. Protein Sci. 2000, 9, 466–475. [Google Scholar] [CrossRef]
- Chen, C.C.; Adolphson, R.; Dean, J.F.D.; Eriksson, K.E.L.; Adams, M.W.W.; Westpheling, J. Release of lignin from kraft pulp by a hyperthermophilic xylanase from Thermatoga maritima. Enzyme Microb. Technol. 1997, 20, 39–45. [Google Scholar] [CrossRef]
- Kulkarni, N.; Gadre, R.V. A novel alkaline, thermostable, protease-free lipase from Pseudomonas sp. Biotechnol. Lett. 1999, 21, 897–899. [Google Scholar] [CrossRef]
- Rehm, F.B.H.; Chen, S.; Rehm, B.H.A. Enzyme engineering for in situ immobilization. Molecules 2016, 21, 1370. [Google Scholar] [CrossRef]
- Thuku, R.N.; Brady, D.; Benedik, M.J.; Sewell, B.T. Microbial nitrilases: versatile, spiral forming, industrial enzymes. J. Appl. Microbiol. 2009, 106, 703–727. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zaro, J.L.; Shen, W.-C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, L.; Gomez, L.; Whitehead, T.; Clark, D. Self-Renaturing Enzymes: Design of an Enzyme-Chaperone Chimera as a New Approach to Enzyme Stabilization. Biotechnol. Bioeng. 2010, 102, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W.; Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; 10 Skyline Drive; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 2001; pp. 11803–12500. [Google Scholar]
Sample Availability: Gene sequences of RrNit, RrGroES, RrGroEL and RrGRoEL2 are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Catalysts. | kd (min−1) | t1/2 (min) |
|---|---|---|
| BL21(DE3) (pET28a-RrNit) | 0.02453 0.00057 | 28.27 0.66 |
| BL21(DE3) (pET28a-RrNit-FxL1-RrGroES) | 0.00445 0.00039 | 156.81 13.90 |
| BL21(DE3) (pET28a-RrNit-FxL2-RrGroES) | 0.00505 0.00051 | 138.67 14.00 |
| BL21(DE3) (pET28a-RrNit-FxL4-RrGroES) | 0.00514 0.00019 | 135.04 4.94 |
| BL21(DE3) (pET28a-RrNit-FxL1-RrGroEL) | 0.00368 0.00053 | 92.34 27.70 |
| BL21(DE3) (pET28a-RrNit-FxL2-RrGroEL) | 0.00286 0.00036 | 246.26 31.00 |
| BL21(DE3) (pET28a-RrNit-FxL4-RrGroEL) | 0.00230 0.00005 | 301.57 7.87 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Tang, L.; Liang, Y.; Jiao, S.; Yu, H.; Luo, H. Novel Chaperones RrGroEL and RrGroES for Activity and Stability Enhancement of Nitrilase in Escherichia coli and Rhodococcus ruber. Molecules 2020, 25, 1002. https://doi.org/10.3390/molecules25041002
Xu C, Tang L, Liang Y, Jiao S, Yu H, Luo H. Novel Chaperones RrGroEL and RrGroES for Activity and Stability Enhancement of Nitrilase in Escherichia coli and Rhodococcus ruber. Molecules. 2020; 25(4):1002. https://doi.org/10.3390/molecules25041002
Chicago/Turabian StyleXu, Chunmeng, Lingjun Tang, Youxiang Liang, Song Jiao, Huimin Yu, and Hui Luo. 2020. "Novel Chaperones RrGroEL and RrGroES for Activity and Stability Enhancement of Nitrilase in Escherichia coli and Rhodococcus ruber" Molecules 25, no. 4: 1002. https://doi.org/10.3390/molecules25041002
APA StyleXu, C., Tang, L., Liang, Y., Jiao, S., Yu, H., & Luo, H. (2020). Novel Chaperones RrGroEL and RrGroES for Activity and Stability Enhancement of Nitrilase in Escherichia coli and Rhodococcus ruber. Molecules, 25(4), 1002. https://doi.org/10.3390/molecules25041002