Adropin as A Fat-Burning Hormone with Multiple Functions—Review of a Decade of Research

 ,
,  , and
, and 
{kind=link}
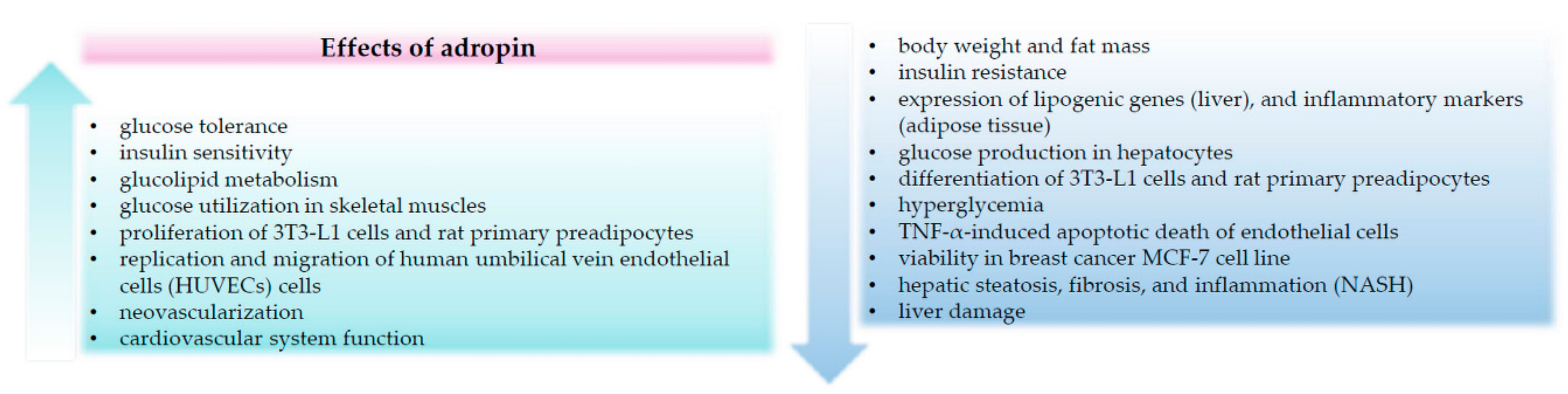
Abstract
1. Discovery of Adropin and the Adropin Receptor
2. Regulation of Enho mRNA Expression
3. Modulation of Adropin by Body Mass Index (BMI), Diet, and Diabetes
4. The Role of Adropin in Controlling Adiposity and Lipid and Glucose Metabolism
5. Adropin in the Cardiovascular System
6. Adropin in the Reproductive System
7. Adropin and Cancer
8. Adropin in Liver Disease
9. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Kumar, K.G.; Trevaskis, J.L.; Lam, D.D.; Sutton, G.M.; Koza, R.A.; Chouljenko, V.N.; Kousoulas, K.G.; Rogers, P.M.; Kesterson, R.A.; Thearle, M.; et al. Identification of adropin as a secreted factor linking dietary macronutrient intake with energy homeostasis and lipid metabolism. Cell Metab. 2008, 8, 468–481. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.A.; Zhang, J.; Price, C.A.; Stevens, J.R.; Graham, J.L.; Stanhope, K.L.; King, S.; Krauss, R.M.; Bremer, A.A.; Havel, P.J. Low plasma adropin concentrations increase risks of weight gain and metabolic dysregulation in response to a high-sugar diet in male nonhuman primates. J. Biol. Chem. 2019, 294, 9706–9719. [Google Scholar] [CrossRef] [PubMed]
- Yolbas, S.; Kara, M.; Kalayci, M.; Yildirim, A.; Gundogdu, B.; Aydin, S.; Koca, S.S. ENHO gene expression and serum adropin level in rheumatoid arthritis and systemic lupus erythematosus. Adv. Clin. Exp. Med. 2018, 27, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.A.; Tam, C.S.; Stanhope, K.L.; Wolfe, B.M.; Ali, M.R.; O’Keeffe, M.; St-Onge, M.P.; Ravussin, E.; Havel, P.J. Low circulating adropin concentrations with obesity and aging correlate with risk factors for metabolic disease and increase after gastric bypass surgery in humans. J. Clin. Endocrinol. Metab. 2012, 97, 3783–3791. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.M.; Yosten, G.L.; Samson, W.K. Adropin acts in brain to inhibit water drinking: potential interaction with the orphan G protein-coupled receptor, GPR19. Am. J. Physiol. 2016, 310, R476–R480. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Herr, D.R. G protein-coupled receptor GPR19 regulates E-cadherin expression and invasion of breast cancer cells. Biochim. Biophys. Acta 2017, 1864, 1318–1327. [Google Scholar] [CrossRef]
- Thapa, D.; Stoner, M.W.; Zhang, M.; Xie, B.; Manning, J.R.; Guimaraes, D.; Shiva, S.; Jurczak, M.J.; Scott, I. Adropin regulates pyruvate dehydrogenase in cardiac cells via a novel GPCR-MAPK-PDK4 signaling pathway. Redox Biol. 2018, 18, 25–32. [Google Scholar] [CrossRef]
- Foster, S.R.; Hauser, A.S.; Vedel, L.; Strachan, R.T.; Huang, X.P.; Gavin, A.C.; Shah, S.D.; Nayak, A.P.; Haugaard-Kedstrom, L.M.; Penn, R.B.; et al. Discovery of Human Signaling Systems: Pairing Peptides to G Protein-Coupled Receptors. Cell 2019, 179, 895.e21–908.e21. [Google Scholar] [CrossRef]
- Wong, C.M.; Wang, Y.; Lee, J.T.; Huang, Z.; Wu, D.; Xu, A.; Lam, K.S. Adropin is a brain membrane-bound protein regulating physical activity via the NB-3/Notch signaling pathway in mice. J. Biol. Chem. 2014, 289, 25976–25986. [Google Scholar] [CrossRef]
- Ghoshal, S.; Stevens, J.R.; Billon, C.; Girardet, C.; Sitaula, S.; Leon, A.S.; Rao, D.C.; Skinner, J.S.; Rankinen, T.; Bouchard, C.; et al. Adropin: An endocrine link between the biological clock and cholesterol homeostasis. Mol. Metab. 2018, 8, 51–64. [Google Scholar] [CrossRef]
- Ganesh Kumar, K.; Zhang, J.; Gao, S.; Rossi, J.; McGuinness, O.P.; Halem, H.H.; Culler, M.D.; Mynatt, R.L.; Butler, A.A. Adropin deficiency is associated with increased adiposity and insulin resistance. Obesity Silver Spring 2012, 20, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Sayin, O.; Tokgoz, Y.; Arslan, N. Investigation of adropin and leptin levels in pediatric obesity-related nonalcoholic fatty liver disease. J. Pediatr. Endocrinol. Metab. 2014, 27, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Y.; Zhao, P.; Wu, M.C.; Liu, J.; Yin, W. Serum adropin levels are decreased in patients with acute myocardial infarction. Regul. Pept. 2014, 190-191, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Yosaee, S.; Khodadost, M.; Esteghamati, A.; Speakman, J.R.; Shidfar, F.; Nazari, M.N.; Bitarafan, V.; Djafarian, K. Metabolic Syndrome Patients Have Lower Levels of Adropin When Compared With Healthy Overweight/Obese and Lean Subjects. Am. J. Men’s Health 2017, 11, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Zang, H.; Jiang, F.; Cheng, X.; Xu, H.; Hu, X. Serum adropin levels are decreased in Chinese type 2 diabetic patients and negatively correlated with body mass index. Endocrine J. 2018, 65, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.N.; Yim, J.E. Plasma Adropin as a Potential Marker Predicting Obesity and Obesity-associated Cancer in Korean Patients With Type 2 Diabetes Mellitus. J. Cancer Prevent. 2018, 23, 191–196. [Google Scholar] [CrossRef]
- Chen, R.M.; Yuan, X.; Ouyang, Q.; Lin, X.Q.; Ai, Z.Z.; Zhang, Y.; Yang, X.H. Adropin and glucagon-like peptide-2 are associated with glucose metabolism in obese children. World J. Pediatr. 2019, 15, 565–571. [Google Scholar] [CrossRef]
- Butler, A.A.; St-Onge, M.P.; Siebert, E.A.; Medici, V.; Stanhope, K.L.; Havel, P.J. Differential Responses of Plasma Adropin Concentrations To Dietary Glucose or Fructose Consumption In Humans. Sci. Rep. 2015, 5, 14691. [Google Scholar] [CrossRef]
- Ebara, T.; Ramakrishnan, R.; Steiner, G.; Shachter, N.S. Chylomicronemia due to apolipoprotein CIII overexpression in apolipoprotein E-null mice. Apolipoprotein CIII-induced hypertriglyceridemia is not mediated by effects on apolipoprotein E. J. Clin. Investig. 1997, 99, 2672–2681. [Google Scholar] [CrossRef]
- St-Onge, M.P.; Shechter, A.; Shlisky, J.; Tam, C.S.; Gao, S.; Ravussin, E.; Butler, A.A. Fasting plasma adropin concentrations correlate with fat consumption in human females. Obesity 2014, 22, 1056–1063. [Google Scholar] [CrossRef]
- Stevens, J.R.; Kearney, M.L.; St-Onge, M.P.; Stanhope, K.L.; Havel, P.J.; Kanaley, J.A.; Thyfault, J.P.; Weiss, E.P.; Butler, A.A. Inverse association between carbohydrate consumption and plasma adropin concentrations in humans. Obesity 2016, 24, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Fang, J.; Chen, L.; Zhao, Z.; Luo, Y.; Lin, C.; Fan, L. Low serum adropin is associated with coronary atherosclerosis in type 2 diabetic and non-diabetic patients. Clin. Chem. Lab. Med. 2014, 52, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Topuz, M.; Celik, A.; Aslantas, T.; Demir, A.K.; Aydin, S.; Aydin, S. Plasma adropin levels predict endothelial dysfunction like flow-mediated dilatation in patients with type 2 diabetes mellitus. J. Investig. Med. 2013, 61, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zeng, K.; Liu, Q.C.; Guo, Z.; Zhang, S.; Chen, X.R.; Lin, J.H.; Wen, J.P.; Zhao, C.F.; Lin, X.H.; et al. Adropin deficiency worsens HFD-induced metabolic defects. Cell Death Dis. 2017, 8, e3008. [Google Scholar] [CrossRef] [PubMed]
- Beigi, A.; Shirzad, N.; Nikpour, F.; Nasli Esfahani, E.; Emamgholipour, S.; Bandarian, F. Association between serum adropin levels and gestational diabetes mellitus; a case-control study. Gynecol. Endocrinol. 2015, 31, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Celik, E.; Yilmaz, E.; Celik, O.; Ulas, M.; Turkcuoglu, I.; Karaer, A.; Simsek, Y.; Minareci, Y.; Aydin, S. Maternal and fetal adropin levels in gestational diabetes mellitus. J. Perinat. Med. 2013, 41, 375–380. [Google Scholar] [CrossRef]
- Hosseini, A.; Shanaki, M.; Emamgholipour, S.; Nakhjavani, M.; Razi, F.; Golmohammadi, T. Elevated serum levels of adropin in patients with type 2 diabetes mellitus and its association with insulin resistance. J. Biol. Today’s World 2016, 5, 44–49. [Google Scholar] [CrossRef]
- Aydin, S.; Kuloglu, T.; Aydin, S.; Eren, M.N.; Yilmaz, M.; Kalayci, M.; Sahin, I.; Kocaman, N.; Citil, C.; Kendir, Y. Expression of adropin in rat brain, cerebellum, kidneys, heart, liver, and pancreas in streptozotocin-induced diabetes. Mol. Cell. Biochem. 2013, 380, 73–81. [Google Scholar] [CrossRef]
- Kuloglu, T.; Aydin, S. Immunohistochemical expressions of adropin and inducible nitric oxide synthase in renal tissues of rats with streptozotocin-induced experimental diabetes. Biotech. Histochem. 2014, 89, 104–110. [Google Scholar] [CrossRef]
- Polkowska, A.; Pasierowska, I.E.; Paslawska, M.; Pawluczuk, E.; Bossowski, A. Assessment of Serum Concentrations of Adropin, Afamin, and Neudesin in Children with Type 1 Diabetes. BioMed Res. Internat. 2019, 2019, 6128410. [Google Scholar] [CrossRef]
- Akcilar, R.; Kocak, F.E.; Simsek, H.; Akcilar, A.; Bayat, Z.; Ece, E.; Kokdasgil, H. Antidiabetic and hypolipidemic effects of adropinin streoptozotocin-induced type 2 diabetic rats. Bratislavske Lekarske Listy 2016, 117, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Thapa, D.; Xie, B.; Manning, J.R.; Zhang, M.; Stoner, M.W.; Huckestein, B.R.; Edmunds, L.R.; Zhang, X.; Dedousis, N.L.; O’Doherty, R.M.; et al. Adropin reduces blood glucose levels in mice by limiting hepatic glucose production. Physiol. Rep. 2019, 7, e14043. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Ghoshal, S.; Zhang, L.; Stevens, J.R.; McCommis, K.S.; Finck, B.N.; Lopaschuk, G.D.; Butler, A.A. The peptide hormone adropin regulates signal transduction pathways controlling hepatic glucose metabolism in a mouse model of diet-induced obesity. J. Biol. Chem. 2019, 294, 13366–13377. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; McMillan, R.P.; Jacas, J.; Zhu, Q.; Li, X.; Kumar, G.K.; Casals, N.; Hegardt, F.G.; Robbins, P.D.; Lopaschuk, G.D.; et al. Regulation of substrate oxidation preferences in muscle by the peptide hormone adropin. Diabetes 2014, 63, 3242–3252. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; McMillan, R.P.; Zhu, Q.; Lopaschuk, G.D.; Hulver, M.W.; Butler, A.A. Therapeutic effects of adropin on glucose tolerance and substrate utilization in diet-induced obese mice with insulin resistance. Mol. Metab. 2015, 4, 310–324. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Ruperez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, E1040. [Google Scholar] [CrossRef]
- Jasaszwili, M.; Wojciechowicz, T.; Billert, M.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Effects of adropin on proliferation and differentiation of 3T3-L1 cells and rat primary preadipocytes. Mol. Cell. Endocrinol. 2019, 496, 110532. [Google Scholar] [CrossRef]
- Celik, A.; Balin, M.; Kobat, M.A.; Erdem, K.; Baydas, A.; Bulut, M.; Altas, Y.; Aydin, S.; Aydin, S. Deficiency of a new protein associated with cardiac syndrome X; called adropin. Cardiovasc. Ther. 2013, 31, 174–178. [Google Scholar] [CrossRef]
- Han, W.; Zhang, C.; Wang, H.; Yang, M.; Guo, Y.; Li, G.; Zhang, H.; Wang, C.; Chen, D.; Geng, C.; et al. Alterations of irisin, adropin, preptin and BDNF concentrations in coronary heart disease patients comorbid with depression. Ann. Transl. Med. 2019, 7, 298. [Google Scholar] [CrossRef]
- Demircelik, B.; Cakmak, M.; Nazli, Y.; Gurel, O.M.; Akkaya, N.; Cetin, M.; Cetin, Z.; Selcoki, Y.; Kurtul, A.; Eryonucu, B. Adropin: A new marker for predicting late saphenous vein graft disease after coronary artery bypass grafting. Clin. Investig. Med. 2014, 37, E338–E344. [Google Scholar] [CrossRef]
- Altincik, A.; Sayin, O. Evaluation of the relationship between serum adropin levels and blood pressure in obese children. J. Pediatr. Endocrinol. Metab. 2015, 28, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Li, H.; Zhu, X.; Gu, H.; Chen, J.; Wang, L.; Harding, P.; Xu, W. Inverse Correlation Between Plasma Adropin and ET-1 Levels in Essential Hypertension: A Cross-Sectional Study. Medicine 2015, 94, e1712. [Google Scholar] [CrossRef] [PubMed]
- Gulen, B.; Eken, C.; Kucukdagli, O.T.; Serinken, M.; Kocyigit, A.; Kilic, E.; Uyarel, H. Adropin levels and target organ damage secondary to high blood pressure in the ED. Am. J. Emerg. Med. 2016, 34, 2061–2064. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Yong, J.; Ni, S.; Ou, W.; Tan, X. Negative association between serum adropin and hypertensive disorders complicating pregnancy. Hypertens. Pregnancy 2019, 38, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Celik, H.T.; Akkaya, N.; Erdamar, H.; Gok, S.; Kazanci, F.; Demircelik, B.; Cakmak, M.; Yigitoglu, R. The Effects of Valsartan and Amlodipine on the Levels of Irisin, Adropin, and Perilipin. Clin. Lab. 2015, 61, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Lovren, F.; Pan, Y.; Quan, A.; Singh, K.K.; Shukla, P.C.; Gupta, M.; Al-Omran, M.; Teoh, H.; Verma, S. Adropin is a novel regulator of endothelial function. Circulation 2010, 122, S185–S192. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yamashita, T.; Shirai, R.; Shibata, K.; Okano, T.; Yamaguchi, M.; Mori, Y.; Hirano, T.; Watanabe, T. Adropin Contributes to Anti-Atherosclerosis by Suppressing Monocyte-Endothelial Cell Adhesion and Smooth Muscle Cell Proliferation. Int. J. Mol. Sci. 2018, 19, 1293. [Google Scholar] [CrossRef]
- Lo Sasso, G.; Schlage, W.K.; Boue, S.; Veljkovic, E.; Peitsch, M.C.; Hoeng, J. The Apoe(-/-) mouse model: A suitable model to study cardiovascular and respiratory diseases in the context of cigarette smoke exposure and harm reduction. J. Transl. Med. 2016, 14, 146. [Google Scholar] [CrossRef]
- Yang, C.; DeMars, K.M.; Hawkins, K.E.; Candelario-Jalil, E. Adropin reduces paracellular permeability of rat brain endothelial cells exposed to ischemia-like conditions. Peptides 2016, 81, 29–37. [Google Scholar] [CrossRef]
- Wu, L.; Fang, J.; Yuan, X.; Xiong, C.; Chen, L. Adropin reduces hypoxia/reoxygenation-induced myocardial injury via the reperfusion injury salvage kinase pathway. Exp. Ther. Med. 2019, 18, 3307–3314. [Google Scholar] [CrossRef]
- Thapa, D.; Xie, B.; Zhang, M.; Stoner, M.W.; Manning, J.R.; Huckestein, B.R.; Edmunds, L.R.; Mullett, S.J.; McTiernan, C.F.; Wendell, S.G.; et al. Adropin treatment restores cardiac glucose oxidation in pre-diabetic obese mice. J. Mol. Cell. Cardiol. 2019, 129, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Altamimi, T.R.; Gao, S.; Karwi, Q.G.; Fukushima, A.; Rawat, S.; Wagg, C.S.; Zhang, L.; Lopaschuk, G.D. Adropin regulates cardiac energy metabolism and improves cardiac function and efficiency. Metab. Clin. Exp. 2019, 98, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Fujie, S.; Hasegawa, N.; Sato, K.; Fujita, S.; Sanada, K.; Hamaoka, T.; Iemitsu, M. Aerobic exercise training-induced changes in serum adropin level are associated with reduced arterial stiffness in middle-aged and older adults. Am. J. Physiol. 2015, 309, H1642–H1647. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, L.; Yang, Y.J.; Ge, R.K.; Zhou, M.; Hu, H.; Liu, H.; Cui, J.; Li, L.L.; Dong, Y.F.; et al. Aerobic exercise improves endothelial function and serum adropin levels in obese adolescents independent of body weight loss. Sci. Rep. 2017, 7, 17717. [Google Scholar] [CrossRef]
- Yildirim, B.; Celik, O.; Aydin, S. Adropin: A key component and potential gatekeeper of metabolic disturbances in policystic ovarian syndrome. Clin. Exp. Obstet. Gynecol. 2014, 41, 310–312. [Google Scholar] [PubMed]
- Kume, T.; Calan, M.; Yilmaz, O.; Kocabas, G.U.; Yesil, P.; Temur, M.; Bicer, M.; Calan, O.G. A possible connection between tumor necrosis factor alpha and adropin levels in polycystic ovary syndrome. J. Endocrinol. Investig. 2016, 39, 747–754. [Google Scholar] [CrossRef]
- Bousmpoula, A.; Kouskouni, E.; Benidis, E.; Demeridou, S.; Kapeta-Kourkouli, R.; Chasiakou, A.; Baka, S. Adropin levels in women with polycystic ovaries undergoing ovarian stimulation: Correlation with lipoprotein lipid profiles. Gynecol. Endocrinol. 2018, 34, 153–156. [Google Scholar] [CrossRef]
- Kuliczkowska-Plaksej, J.; Mierzwicka, A.; Jonczyk, M.; Stachowska, B.; Urbanovych, A.; Bolanowski, M. Adropin in women with polycystic ovary syndrome. Endokrynologia Polska 2019, 70, 151–156. [Google Scholar] [CrossRef]
- Chazenbalk, G.; Trivax, B.S.; Yildiz, B.O.; Bertolotto, C.; Mathur, R.; Heneidi, S.; Azziz, R. Regulation of adiponectin secretion by adipocytes in the polycystic ovary syndrome: role of tumor necrosis factor-{alpha}. J. Clin. Endocrinol. Metab. 2010, 95, 935–942. [Google Scholar] [CrossRef]
- Qiu, X.; He, J.R.; Zhao, M.G.; Kuang, Y.S.; Xu, S.Q.; Zhang, H.Z.; Hu, S.P.; Chen, J.; Xia, H.M. Relationship between human cord blood adropin levels and fetal growth. Peptides 2014, 52, 19–22. [Google Scholar] [CrossRef]
- Aydin, S.; Kuloglu, T.; Aydin, S. Copeptin, adropin and irisin concentrations in breast milk and plasma of healthy women and those with gestational diabetes mellitus. Peptides 2013, 47, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S. Presence of adropin, nesfatin-1, apelin-12, ghrelins and salusins peptides in the milk, cheese whey and plasma of dairy cows. Peptides 2013, 43, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.J. Development of the newborn GI tract and its relation to colostrum/milk intake: A review. Reprod. Fertil. Dev. 1996, 8, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Tuna, B.G.; Atalay, P.B.; Altunbek, M.; Kalkan, B.M.; Dogan, S. Effects of Chronic and Intermittent Calorie Restriction on Adropin Levels in Breast Cancer. Nutr. Cancer 2017, 69, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Nergiz, S.; Altinkaya, S.O.; Kurt Omurlu, I.; Yuksel, H.; Kucuk, M.; Demircan Sezer, S. Circulating adropin levels in patients with endometrium cancer. Gynecol. Endocrinol. 2015, 31, 730–735. [Google Scholar] [CrossRef]
- Prystupa, A.; Kicinski, P.; Luchowska-Kocot, D.; Sak, J.; Prystupa, T.; Chen, K.H.; Chen, Y.C.; Panasiuk, L.; Zaluska, W. Afamin and adropin in patients with alcohol-induced liver cirrhosis. Ann. Agric. Environ. Med. 2018, 25, 527–531. [Google Scholar] [CrossRef]
- Chen, X.; Xue, H.; Fang, W.; Chen, K.; Chen, S.; Yang, W.; Shen, T.; Chen, X.; Zhang, P.; Ling, W. Adropin protects against liver injury in nonalcoholic steatohepatitis via the Nrf2 mediated antioxidant capacity. Redox Biol. 2019, 21, 101068. [Google Scholar] [CrossRef]
- Kutlu, O.; Altun, O.; Dikker, O.; Aktas, S.; Ozsoy, N.; Arman, Y.; Ozgun Cil, E.; Ozcan, M.; Aydin Yoldemir, S.; Akarsu, M.; et al. Serum Adropin Levels Are Reduced in Adult Patients with Nonalcoholic Fatty Liver Disease. Med. Princ. Pract. 2019, 28, 463–469. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasaszwili, M.; Billert, M.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Adropin as A Fat-Burning Hormone with Multiple Functions—Review of a Decade of Research. Molecules 2020, 25, 549. https://doi.org/10.3390/molecules25030549
Jasaszwili M, Billert M, Strowski MZ, Nowak KW, Skrzypski M. Adropin as A Fat-Burning Hormone with Multiple Functions—Review of a Decade of Research. Molecules. 2020; 25(3):549. https://doi.org/10.3390/molecules25030549
Chicago/Turabian StyleJasaszwili, Mariami, Maria Billert, Mathias Z. Strowski, Krzysztof W. Nowak, and Marek Skrzypski. 2020. "Adropin as A Fat-Burning Hormone with Multiple Functions—Review of a Decade of Research" Molecules 25, no. 3: 549. https://doi.org/10.3390/molecules25030549
APA StyleJasaszwili, M., Billert, M., Strowski, M. Z., Nowak, K. W., & Skrzypski, M. (2020). Adropin as A Fat-Burning Hormone with Multiple Functions—Review of a Decade of Research. Molecules, 25(3), 549. https://doi.org/10.3390/molecules25030549