
Cardioprotective Activity of Selected Polyphenols Based on Epithelial and Aortic Cell Lines. A Review





Abstract
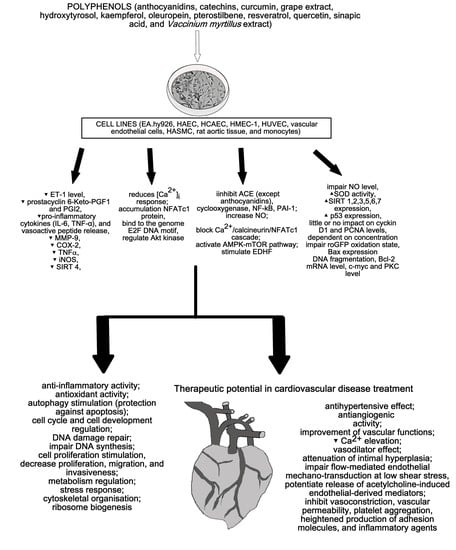
1. Introduction
2. Cardioprotective Activity of Polyphenols
2.1. Flavonoids
2.2. Nonflavonoids
2.3. Phenolic Acids
2.4. Polyphenol Containing Plant Concentrates, Extracts, and Fractions
3. Discussion
- decrease of ACE, ET-1, ROS production, endothelial tube formation, migration, endothelin 1, and tissue plasminogen activator release;
- stimulation of acetylcholine-induced endothelial-derived mediator release, autophagy, NO, DNA synthesis, and cell cycle regulation;
- increase of viability; and
- regulation of sirtuins.
Author Contributions
Funding
Conflicts of Interest
References
- Dias-Pedroso, D.; Guerra, J.; Gomes, A.; Oudot, C.; Brenner, C.; Santos, C.N.; Vieira, H.L.A. Phenolic metabolites modulate cardiomyocyte beating in response to isoproterenol. Cardiovasc. Toxicol. 2019, 19, 156–167. [Google Scholar] [CrossRef]
- Balea, Ş.S.; Pârvu, A.E.; Pop, N.; Marín, F.Z.; Pârvu, M. Polyphenolic compounds, antioxidant, and cardioprotective effects of pomace extracts from Fetească Neagră Cultivar. Oxid. Med. Cell Longev. 2018, 8194721. [Google Scholar] [CrossRef]
- Chu, A.J. Cardioprotection by bioactive polyphenols: A strategic view. Austin, J. Cardiovasc. Dis. Atherosclerosis 2018, 5, 1034. [Google Scholar]
- Thandapilly, S.J.; Wojciechowski, P.; Behbahani, J.; Louis, X.L.; Yu, L.; Juric, D.; Kopilas, M.A.; Anderson, H.D.; Netticadan, T. Resveratrol prevents the development of pathological cardiac hypertrophy and contractile dysfunction in the SHR without lowering blood pressure. Am. J. Hypertens. 2010, 23, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Grootaert, C.; Kamiloglu, S.; Capanoglu, E.; Van Camp, J. Cell systems to investigate the impact of polyphenols on cardiovascular health. Nutrients 2015, 7, 9229–9255. [Google Scholar] [CrossRef]
- Atrahimovich, D.; Samson, A.O.; Barsheshet, Y.; Vaya, J.; Khatib, S.; Reuveni, E. Genome-wide localization of the polyphenol quercetin in human monocytes. BMC Genomics 2019, 20, 606. [Google Scholar] [CrossRef]
- Goszcz, K.; Duthie, G.G.; Stewart, D.; Leslie, S.J.; Megson, I.L. Bioactive polyphenols and cardiovascular disease: Chemical antagonists, pharmacological agents or xenobiotics that drive an adaptive response? Br. J. Pharmacol. 2017, 174, 1209–1225. [Google Scholar] [CrossRef]
- Michalska, M.; Gluba, A.; Mikhailidis, D.P.; Nowak, P.; Bielecka-Dabrowa, A.; Rysz, J.; Banach, M. The role of polyphenols in cardiovascular disease. Med. Sci. Monit. 2010, 16, RA110–RA119. [Google Scholar] [PubMed]
- Andriantsitohaina, R.; Auger, C.; Chataigneau, T.; Étienne-Selloum, N.; Li, H.; Martínez, M.C.; Schini-Kerth, V.B.; Laher, I. Molecular mechanisms of the cardiovascular protective effects of polyphenols. Br. J. Nutr. 2012, 108, 1532–1549. [Google Scholar] [CrossRef]
- WHO Home Page. Available online: https://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 2 September 2020).
- Eurostat Home Page. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php/Cardiovascular_disease_statistics#Deaths_from_cardiovascular_diseases (accessed on 2 September 2020).
- European Hearth Network Home Page. Available online: http://www.ehnheart.org/cvd-statistics.html (accessed on 2 September 2020).
- American College of Cardiology Home Page. Available online: https://www.acc.org/latest-in-cardiology/ten-points-to-remember/2019/02/15/14/39/aha-2019-heart-disease-and-stroke-statistics (accessed on 7 November 2020).
- OECD iLibrary Home Page. Available online: https://www.oecd-ilibrary.org/sites/4b1b32a8-en/index.html?itemId=/content/component/4b1b32a8-en (accessed on 7 November 2020).
- Persson, I.A.; Persson, K.; Andersson, R.G. Effect of Vaccinium myrtillus and its polyphenols on angiotensin-converting enzyme activity in human endothelial cells. J. Agric. Food Chem. 2009, 57, 4626–4629. [Google Scholar] [CrossRef] [PubMed]
- Negrão, R.; Costa, R.; Duarte, D.; Gomes, T.T.; Azevedo, I.; Soares, R. Different effects of catechin on angiogenesis and inflammation depending on VEGF levels. J. Nutr. Biochem. 2013, 24, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Olszanecki, R.; Bujak-Gizycka, B.; Madej, J.; Suski, M.; Wołkow, P.P.; Jawień, J.; Korbut, R. Kaempferol, but not resveratrol inhibits angiotensin converting enzyme. J. Physiol. Pharmacol. 2008, 59, 387–392. [Google Scholar] [PubMed]
- Zhao, X.; Gu, Z.; Attele, A.S.; Yuan, C.S. Effects of quercetin on the release of endothelin, prostacyclin and tissue plasminogen activator from human endothelial cells in culture. J. Ethnopharmacol. 1999, 67, 279–285. [Google Scholar] [CrossRef]
- Scoditti, E.; Calabriso, N.; Massaro, M.; Pellegrino, M.; Storelli, C.; Martines, G.; De Caterina, R.; Carluccio, M.A. Mediterranean diet polyphenols reduce inflammatory angiogenesis through MMP-9 and COX-2 inhibition in human vascular endothelial cells: A potentially protective mechanism in atherosclerotic vascular disease and cancer. Arch. Biochem. Biophys. 2012, 527, 81–89. [Google Scholar] [CrossRef]
- Akinwumi, B.C.; Bordun, K.M.; Anderson, H.D. Biological activities of stilbenoids. Int. J. Mol. Sci. 2018, 19, 792. [Google Scholar] [CrossRef]
- Kang, D.G.; Kim, Y.C.; Sohn, E.J.; Lee, Y.M.; Lee, A.S.; Yin, M.H.; Lee, H.S. Hypotensive effect of butein via the inhibition of angiotensin converting enzyme. Biol. Pharm. Bull. 2003, 26, 1345–1347. [Google Scholar] [CrossRef]
- Liu, J.C.; Hsu, F.L.; Tsai, J.C.; Chan, P.; Liu, J.Y.; Thomas, G.N.; Tomlinson, B.; Lo, M.Y.; Lin, J.Y. Antihypertensive effects of tannins isolated from traditional Chinese herbs as non-specific inhibitors of angiontensin converting enzyme. Life Sci. 2003, 73, 1543–1555. [Google Scholar] [CrossRef]
- Hernández, M.; Wicz, S.; Santamaría, M.H.; Corral, R.S. Curcumin exerts anti-inflammatory and vasoprotective effects through amelioration of NFAT-dependent endothelin-1 production in mice with acute Chagas cardiomyopathy. Mem. Inst. Oswaldo Cruz 2018, 113, e180171. [Google Scholar] [CrossRef]
- Catalogna, G.; Moraca, F.; D’Antona, L.; Dattilo, V.; Perrotti, G.; Lupia, A.; Costa, G.; Ortuso, F.; Iuliano, R.; Trapasso, F.; et al. Review about the multi-target profile of resveratrol and its implication in the SGK1 inhibition. Eur, J. Med. Chem. 2019, 183, 111675. [Google Scholar] [CrossRef]
- Thandapilly, S.J.; Louis, X.L.; Behbahani, J.; Movahed, A.; Yu, L.; Fandrich, R.; Zhang, S.; Kardami, E.; Anderson, H.D.; Netticadan, T. Reduced hemodynamic load aids low-dose resveratrol in reversing cardiovascular defects in hypertensive rats. Hypertens. Res. 2013, 36, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Riche, D.M.; Riche, K.D.; Blackshear, C.T.; McEwen, C.L.; Sherman, J.J.; Wofford, M.R.; Griswold, M.E. Pterostilbene on metabolic parameters: A randomized.; double-blind.; and placebo-controlled trial. Evid. Based Complementary Alternat. Med. 2014, 459165. [Google Scholar] [CrossRef] [PubMed]
- Remsberg, C.M.; Martinez, S.E.; Akinwumi, B.C.; Anderson, H.D.; Takemoto, J.K.; Sayre, C.L.; Davies, N.M. Preclinical pharmacokinetics and pharmacodynamics and content analysis of gnetol in foodstuffs. Phytother. Res. 2015, 29, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. One-year consumption of a grape nutraceutical containing resveratrol improves the inflammatory and fibrinolytic status of patients in primary prevention of cardiovascular disease. Am. J. Cardiol. 2012, 110, 356–363. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Grape resveratrol increases serum adiponectin and downregulates inflammatory genes in peripheral blood mononuclear cells: A triple-blind, placebo-controlled, one-year clinical trial in patients with stable coronary artery disease. Cardiovasc. Drugs Ther. 2013, 27, 37–48. [Google Scholar] [CrossRef]
- Song, J.; Huang, Y.; Zheng, W.; Yan, J.; Cheng, M.; Zhao, R.; Chen, L.; Hu, C.; Jia, W. Resveratrol reduces intracellular reactive oxygen species levels by inducing autophagy through the AMPK-mTOR pathway. Front. Med. 2018, 12, 697–706. [Google Scholar] [CrossRef]
- Diaz, M.; Parikh, V.; Ismail, S.; Maxamed, R.; Tye, E.; Austin, C.; Dew, T.; Graf, B.A.; Vanhees, L.; Degens, H.; et al. Differential effects of resveratrol on the dilator responses of femoral arteries, ex vivo. Nitric Oxide 2019, 92, 1–10. [Google Scholar] [CrossRef]
- Posadino, A.M.; Giordo, R.; Cossu, A.; Nasrallah, G.K.; Shaito, A.; Abou-Saleh, H.; Eid, A.H.; Pintus, G. Flavin oxidase-induced ROS generation modulates PKC biphasic effect of resveratrol on endothelial cell survival. Biomolecules 2019, 9, 209. [Google Scholar] [CrossRef]
- Yu, H.; Pan, W.; Huang, H.; Chen, J.; Sun, B.; Yang, L.; Zhu, P. Screening analysis of sirtuins family expression on anti-inflammation of resveratrol in endothelial cells. Med. Sci. Monit. 2019, 25, 4137–4148. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Labinskyy, N.; Hsieh, T.C.; Ungvari, Z.; Wu, J.M. Regulation of proliferation and gene expression in cultured human aortic smooth muscle cells by resveratrol and standardized grape extracts. Biochem. Biophys. Res. Commun. 2006, 346, 367–376. [Google Scholar] [CrossRef]
- Silambarasan, T.; Manivannan, J.; Krishna Priya, M.; Suganya, N.; Chatterjee, S.; Raja, B. Sinapic acid prevents hypertension and cardiovascular remodeling in pharmacological model of nitric oxide inhibited rats. PLoS ONE 2014, 9, e115682. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Corbi, G.; Righetti, S.; Sears, B.; Olarte, H.H.; Grassi, D.; Scapagnini, G. Cardioprotection by cocoa polyphenols and ω-3 fatty acids: A disease-prevention perspective on aging-associated cardiovascular risk. J. Med. Food 2018, 21, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Peters, U.; Poole, C.; Arab, L. Does tea affect cardiovascular disease? A meta-analysis. Am. J. Epidemiol. 2001, 154, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Rzepecka-Stojko, A.; Stojko, J.; Jasik, K.; Buszman, E. Anti-Atherogenic Activity of polyphenol-rich extract from bee pollen. Nutrients 2017, 9, 1369. [Google Scholar] [CrossRef] [PubMed]
- Rzepecka-Stojko, A.; Kabała-Dzik, A.; Kubina, R.; Jasik, K.; Kajor, M.; Wrześniok, D.; Stojko, J. Protective effect of polyphenol-rich extract from bee pollen in a high-fat diet. Molecules (Basel, Switzerland) 2018, 23, 805. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Cardiovascular Diseases | ||
|---|---|---|
| Number of Death Cases | Deaths (%) | |
| World Health Organization [11] | 17.9 million (globally) | 31.0 (globally) |
| Eurostat statistics [12] | 1.8 million * | 35.7 * |
| European Heart Network [13] | 3.9 million ** and 1.8 million * | 45 ** and 37 * |
| Country | Stroke Death Cases per 100,000 Population in 2017 [15] |
|---|---|
| Canada | 35.1 |
| Australia | 42.0 |
| USA | 43.2 |
| Japan | 46.1 |
| Korea | 56.1 |
| Poland | 71.0 |
| Brazil | 81.5 |
| Turkey | 97.2 |
| South Africa | 140.7 |
| Russia | 234.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otręba, M.; Kośmider, L.; Stojko, J.; Rzepecka-Stojko, A. Cardioprotective Activity of Selected Polyphenols Based on Epithelial and Aortic Cell Lines. A Review. Molecules 2020, 25, 5343. https://doi.org/10.3390/molecules25225343
Otręba M, Kośmider L, Stojko J, Rzepecka-Stojko A. Cardioprotective Activity of Selected Polyphenols Based on Epithelial and Aortic Cell Lines. A Review. Molecules. 2020; 25(22):5343. https://doi.org/10.3390/molecules25225343
Chicago/Turabian StyleOtręba, Michał, Leon Kośmider, Jerzy Stojko, and Anna Rzepecka-Stojko. 2020. "Cardioprotective Activity of Selected Polyphenols Based on Epithelial and Aortic Cell Lines. A Review" Molecules 25, no. 22: 5343. https://doi.org/10.3390/molecules25225343
APA StyleOtręba, M., Kośmider, L., Stojko, J., & Rzepecka-Stojko, A. (2020). Cardioprotective Activity of Selected Polyphenols Based on Epithelial and Aortic Cell Lines. A Review. Molecules, 25(22), 5343. https://doi.org/10.3390/molecules25225343