Recent Progress in Amaryllidaceae Biotechnology




Abstract
1. Introduction
2. Plant In Vitro Systems Producing Amaryllidaceae Alkaloids
3. Biotechnological Production of Amaryllidaceae Alkaloids
3.1. Selection of Primary Plant Material
3.2. Decision on the Type of Plant In Vitro System
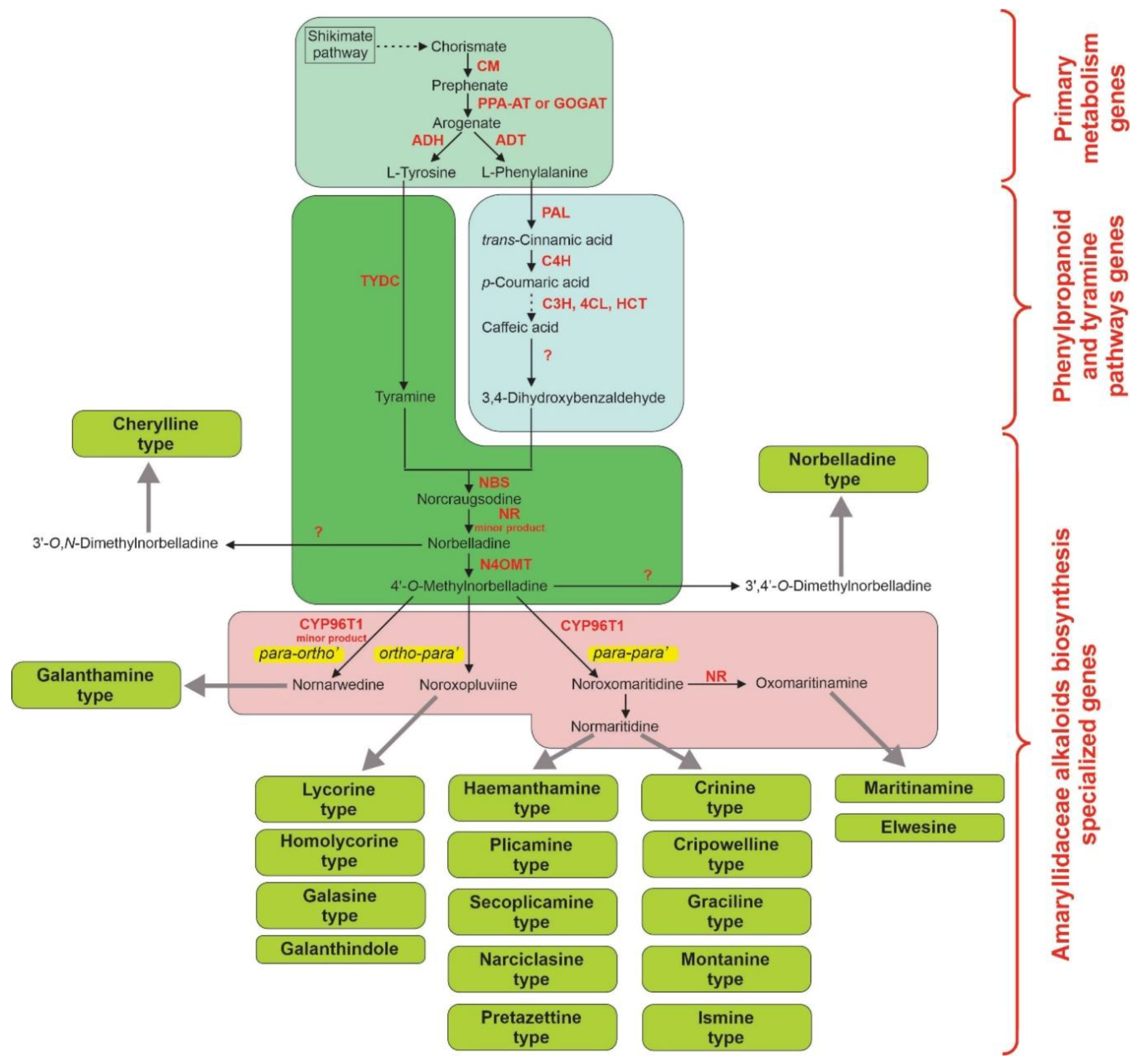
3.3. Metabolic Engineering and Synthetic Biology Approaches to Increase the Production of Amaryllidaceae Alkaloids
3.4. Optimization of the Biosynthetic Process
4. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Takos, A.M.; Rook, F. Towards a molecular understanding of the biosynthesis of amaryllidaceae alkaloids in support of their expanding medical use. Int. J. Mol. Sci. 2013, 14, 11713–11741. [Google Scholar] [CrossRef]
- Jin, Z.; Xu, X.H. Amaryllidaceae alkaloids. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin, Germany, 2013; pp. 479–522. [Google Scholar]
- Jin, Z. Amaryllidaceae and Sceletium alkaloids. Nat. Prod. Rep. 2005, 22, 111–126. [Google Scholar] [CrossRef]
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and biological aspects of Narcissus alkaloids. In Alkaloids: Chemistry and Biology; Geoffrey, A.C., Ed.; Academic Press: New York, NY, USA, 2006; Volume 63, pp. 87–179. [Google Scholar]
- Nair, J.J.; Bastida, J.; Codina, C.; Viladomat, F.; van Staden, J. Alkaloids of the South African Amaryllidaceae: A review. Nat. Prod. Comm. 2013, 8, 1335–1350. [Google Scholar] [CrossRef]
- Nair, J.J.; van Staden, J. Pharmacological and toxicological insights to the South African Amaryllidaceae. Food Chem. Toxicol. 2013, 6, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.J.; van Staden, J. Traditional usage, phytochemistry and pharmacology of the South African medicinal plant Boophone disticha (L.f.) Herb. (Amaryllidaceae). J. Ethnopharmacol. 2014, 151, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Amaryllidaceae alkaloids: Absolute configuration and biological activity. Chirality 2017, 29, 486–499. [Google Scholar] [CrossRef]
- Harvey, A.L. The pharmacology of galanthamine and its analogues. Pharmacol. Ther. 1995, 68, 113–128. [Google Scholar] [CrossRef]
- Rhee, I.K.; van de Meent, M.; Ingkaninan, K.; Verpoorte, R. Screening for acetylcholinesterase inhibitors from Amaryllidaceae using silica gel thin-layer chromatography in combination with bioactivity staining. J. Chromatogr. A 2001, 915, 217–223. [Google Scholar] [CrossRef]
- Farinon, M.; Clarimundo, V.S.; Pedrazza, G.P.; Gulko, P.S.; Zuanazzi, J.A.; Xavier, R.M.; de Oliveira, P.G. Disease modifying anti-rheumatic activity of the alkaloid montanine on experimental arthritis and fibroblast-like synoviocytes. Eur. J. Pharmacol. 2017, 799, 180–187. [Google Scholar] [CrossRef]
- da Silva, A.F.; de Andrade, J.P.; Bevilaqua, L.R.; de Souza, M.M.; Izquierdo, I.; Henriques, A.T.; Zuanazzi, J.A. Anxiolytic-, antidepressant- and anticonvulsant-like effects of the alkaloid montanine isolated from Hippeastrum vittatum. Pharmacol. Biochem. Behav. 2006, 85, 148–154. [Google Scholar] [CrossRef]
- de Andrade, J.P.; Pigni, N.B.; Torras-Claveria, L.; Guo, Y.; Berkov, S.; Reyes-Chilpa, R.; el Amrani, A.; Zuanazzi, J.A.S.; Codina, C.; Viladomat, F.; et al. Alkaloids from the Hippeastrum genus: Chemistry and biological activity. Rev. Latinoam. Quim. 2012, 40, 83–98. [Google Scholar]
- Szlávik, L.; Gyuris, A.; Minárovits, J.; Forgo, P.; Molnár, J.; Hohmann, J. Alkaloids from Leucojum vernum and antiretroviral activity of Amaryllidaceae alkaloids. Planta Med. 2004, 70, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Tahchy, A.E.; Boisbrun, M.; Ptak, A.; Dupire, F.; Chrétien, F.; Henry, M.; Chapleur, Y.; Laurain-Mattar, D. New method for the study of biosynthesis using biotransformation of deuterium-labeled precursor in tissue cultures. Acta Biochem. Pol. 2010, 57, 75–82. [Google Scholar] [CrossRef]
- Houghton, P.J.; Ren, Y.; Howes, M.J. Acetylcholinesterase inhibitors from plants and fungi. Nat. Prod. Rep. 2006, 23, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Liang, L.; Xiao, X.; Feng, P.; Ye, M.; Liu, J. Lycorine: A prospective natural lead for anticancer drug discovery. Biomed. Pharmacother. 2018, 107, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Koutová, D.; Maafi, N.; Havelek, R.; Opletal, L.; Blunden, G.; Rezácová, M.; Cahlíková, L. Chemical and Biological Aspects of Montanine-Type Alkaloids Isolated from Plants of the Amaryllidaceae Family. Molecules 2020, 25, 2337. [Google Scholar] [CrossRef]
- Kornienko, A.; Evidente, A. Chemistry, Biology, and Medicinal Potential of Narciclasine and its Congeners. Chem. Rev. 2008, 108, 1982–2014. [Google Scholar]
- Goietsenoven, G.V.; Andolfi, A.; Lallemand, B.; Cimmino, A.; Lamoral-Theys, D.; Gras, T.; Abou-Donia, A.; Dubois, J.; Lefranc, F.; Mathieu, V.; et al. Amaryllidaceae alkaloids belonging to different structural subgroups display activity against apoptosis-resistant cancer cells. J. Nat. Prod. 2010, 73, 1223–1227. [Google Scholar] [CrossRef]
- Pavlov, A.; Berkov, S.; Courot, E.; Gocheva, T.; Tuneva, D.; Pandova, B.; Georgiev, M.; Georgiev, V.; Yanev, S.; Burrus, M. Galanthamine production by Leucojum aestivum in vitro systems. Proc. Biochem. 2007, 42, 734–739. [Google Scholar] [CrossRef]
- Ferdausi, A.; Chang, X.; Hall, A.; Jones, M. Galanthamine production in tissue culture and metabolomic study on Amaryllidaceae alkaloids in Narcissus pseudonarcissus cv. Carlton. Ind. Crop. Prod. 2020, 144, 112058. [Google Scholar] [CrossRef]
- Bogdanova, Y.; Pandova, B.; Yanev, S.; Stanilova, M. Biosynthesis of lycorine by in vitro cultures of Pancratium maritimum L. (Amaryllidaceae). Biotechnol. Biotechnol. Equip. 2009, 23, 919–922. [Google Scholar] [CrossRef]
- Georgiev, V.; Ivanov, I.; Berkov, S.; Ilieva, M.; Georgiev, M.; Gocheva, T.; Pavlov, A. Galanthamine production by Leucojum aestivum L. shoot culture in a modified bubble column bioreactor with internal sections. Eng. Life Sci. 2012, 12, 534–543. [Google Scholar] [CrossRef]
- Schumann, A.; Berkov, S.; Claus, D.; Gerth, A.; Bastida, J.; Codina, C. Production of galanthamine by Leucojum aestivum shoots grown in different bioreactor systems. Appl. Biochem. Biotechnol. 2012, 167, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Stanilova, M.I.; Molle, E.D.; Yanev, S.G. Galanthamine production by Leucojum aestivum cultures in vitro. In The Alkaloids; Cordell, G.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 68, pp. 167–270. [Google Scholar]
- Berkov, S.; Ivanov, I.; Georgiev, V.; Codina, C.; Pavlov, A. Galanthamine biosynthesis in plant in vitro systems. Eng. Life Sci. 2014, 14, 643–650. [Google Scholar] [CrossRef]
- Laurain-Mattar, D.; Ptak, A. Amaryllidaceae Alkaloid accumulation by plant in vitro systems. In Bioprocessing of Plant In Vitro Systems, Reference Series in Phytochemistry; Pavlov, A., Bley, T.H., Eds.; Springer International Publishing: Geneva, Switzerland, 2018; pp. 203–224. [Google Scholar]
- Georgiev, V.; Berkov, S.; Georgiev, M.; Burrus, M.; Codina, C.; Bastida, J.; Ilieva, M.; Pavlov, A. Optimized nutrient nedium for galanthamine production in Leucojum aestivum L. in vitro shoot system. Z. Naturforsch. 2009, 64C, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.; Georgiev, V.; Georgiev, M.; Ilieva, M.; Pavlov, A. Galanthamine and related alkaloids production by Leucojum aestivum L. shoot culture using a temporary immersion technology. Appl. Biochem. Biotechnol. 2011, 163, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.; Georgiev, V.; Berkov, S.; Pavlov, A. Alkaloid patterns in Leucojum aestivum shoot culture cultivated at temporary immersion conditions. J. Plant. Physiol. 2012, 169, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.; Georgiev, V.; Pavlov, A. Elicitation of galanthamine biosynthesis by Leucojum aestivum liquid shoot cultures. J. Plant. Physiol. 2013, 170, 1122–1129. [Google Scholar] [CrossRef]
- Ivanov, I.; Berkov, S.; Pavlov, A.; Georgiev, V. In sito galanthamine extraction during the cultivation of Leucojum aestivum L. shoot culture in two—Phase bubble column cultivation system. Eng. Life Sci. 2019, 19, 1000–1005. [Google Scholar] [CrossRef]
- Fales, H.M.; Mann, J.; Mudd, S.H. In vitro alkaloid biosynthesis in the Amaryllidaceae; Norbelladine O-Methylpherase. J. Am. Chem. Soc. 1963, 85, 2025–2026. [Google Scholar]
- Sellés, M.; Bergoñón, S.; Viladomat, F.; Bastida, J.; Codina, C. Effect of sucrose on growth and galanthamine production in shoot-clump cultures of Narcissus confusus in liquid-shake medium. Plant. Cell Tissue. Org. Cult. 1997, 4, 129–136. [Google Scholar] [CrossRef]
- Berkov, S.; Pavlov, A.; Georgiev, V.; Bastida, J.; Burrus, M.; Ilieva, M.; Codina, C. Alkaloid synthesis and accumulation in Leucojum aestivum in vitro cultures. Nat. Prod. Comm. 2009, 4, 359–364. [Google Scholar] [CrossRef]
- Berkov, S.; Pavlov, A.; Georgiev, V.; Weber, J.; Bley, T.; Viladomat, F.; Bastida, J.; Codina, C. Changes in apolar metabolites during in vitro organogenesis of Pancratium maritimum. Plant. Physiol. Biochem. 2010, 48, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Tarakemeh, A.; Azizi, M.; Rowshan, V.; Salehi, H.; Spina, R.; Dupire, F.; Arouie, H.; Laurain-Mattar, D. Screening of Amaryllidaceae alkaloids in bulbs and tissue cultures of Narcissus papyraceus and four varieties of Narcissus tazetta. J. Pharm. Biomed. Anal. 2019, 172, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Tarakemeh, A.; Azizi, M.; Rowshan, V.; Salehi, H.; Spina, R.; Dupire, F.; Arouie, H.; Laurain-Mattar, D. Quantitative determination of lycorine and galanthamine in different in vitro tissues of Narcissus tazetta by GC-MS. Int. J. Hort. Sci. Technol. 2019, 6, 151–157. [Google Scholar]
- Bergonon, S.; Codina, C.; Bastida, J.; Viladomat, E.; Mele, E. Galanthamine production in “shoot-clump” cultures of Narcissus confusus in liquid-shake medium. Plant Cell Tissue Org. Cult. 1996, 45, 191–199. [Google Scholar] [CrossRef]
- Subramaniam, S.; Sundarasekar, J.; Sahgal, G.; Murugaiyah, V. Comparative analysis of lycorine in wild plant and callus culture samples of Hymenocallis littoralis by HPLC-UV method. Sci. World J. 2014, 408306. [Google Scholar]
- Reis, A.; Magne, K.; Massot, S.; Tallini, L.R.; Scopel, M.; Bastida, J.; Ratet, P.; Zuanazzi, J.A.S. Amaryllidaceae alkaloids: Identification and partial characterization of montanine production in Rhodophiala bifida plant. Sci. Rep. 2019, 9, 8471. [Google Scholar] [CrossRef]
- Georgiev, V.; Ivanov, I.; Berkov, S.; Pavlov, A. Temporary immersion systems for Amaryllidaceae alkaloids biosynthesis by Pancratium maritimum L. shoot culture. J. Plant. Biochem. Biotechnol. 2014, 23, 389–398. [Google Scholar] [CrossRef]
- Ptak, A.; El Tahchy, A.; Dupire, F.; Boisbrun, M.; Henry, M.; Chapleur, Y.; Mos, M.; Laurain-Mattar, D. LCMS and GCMS for the screening of alkaloids in natural and in vitro extracts of Leucojum aestivum. J. Nat. Prod. 2009, 72, 142–147. [Google Scholar] [CrossRef]
- Georgieva, L.; Berkov, S.; Kondakova, V.; Bastida, J.; Viladomar, F.; Atanasov, A.; odina, C. Alkaloid variability in Leucojum aestivum from wild populations. Z. Naturforsch. C 2007, 62, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Fennell, C.W.; Elgorashi, E.E.; van Staden, J. Alkaloid production in Crinum moorei cultures. J. Nat. Prod. 2003, 66, 1524–1526. [Google Scholar] [CrossRef] [PubMed]
- Zayed, R.; El-Shamy, H.; Berkov, S.; Bastida, J.; Codina, C. In vitro micropropagation and alkaloids of Hippeastrum vittatum. Vitro Cell. Dev. Biol. Plant. 2011, 47, 695–701. [Google Scholar] [CrossRef]
- Pereira da Costa, G.G.; Gasca Silva, C.A.; Dutra Gomes, J.V.; Torres, A.G.; Imaculada Santos, I.R.; Cunha de Almeida, F.T.; Fagg, C.W.; Simeoni, L.A.; Silveira, D.; Gomes-Copeland, K.K.P. Influence of in vitro micropropagation on lycorine biosynthesis and anticholinesterase activity in Hippeastrum goianum. Rev. Bras. Farmacogn. 2019, 2, 262–265. [Google Scholar] [CrossRef]
- Pavlov, A. Plant cells and algae in bioreactors. Eng. Life Sci. 2009, 9, 154–155. [Google Scholar] [CrossRef]
- Pavlov, A. Plant cells and algae in bioreactors II. Eng. Life Sci. 2014, 14, 548–549. [Google Scholar]
- Pavlov, A. Plant cell and algae in bioreactors III. Eng. Life Sci. 2019, 19, 828–829. [Google Scholar] [CrossRef]
- Pavlov, A.; Bley, T. Bioprocessing of Plant In Vitro Systems, Reference Series in Phytochemistry; Springer International Publishing: Geneva, Switzerland, 2018; ISBN 978-3-319-54599-8. [Google Scholar]
- Diop, M.F.; Hehn, A.; Ptak, A.; Chretien, F.; Doerper, S.; Gontier, E.; Bourgaud, F.; Henry, M.; Chapleur, Y.; Laurain-Mattar, D. Hairy root and tissue cultures of Leucojum aestivum L.—Relationships to galanthamine content. Phytochem. Rev. 2007, 6, 137–141. [Google Scholar] [CrossRef]
- Gussev, C.; Bosseva, Y.; Pandova, B.; Yanev, S.; Stanilova, M. Resource assessment of Leucojum aestivum L. (Amaryllidaceae) populations in Bulgaria. Bocconea 2007, 21, 405–411. [Google Scholar]
- Bogdanova, Y.; Stoeva, T.; Yanev, S.; Pandova, B.; Molle, E.; Burrus, M.; Stanilova, M. Influence of plant origin on propagation capacity and alkaloid biosynthesis during long-term in vitro cultivation of Leucojum aestivul L. Vitro Cell. Dev. Biol. Plant. 2009, 45, 458–465. [Google Scholar] [CrossRef]
- Ptak, A.; El Tahchy, A.; Skrzypek, E.; Wójtowicz, T.; Laurain-Mattar, D. Influence of auxins on somatic embryogenesis and alkaloid accumulation in Leucojum aestivum callus. Cent. Eur. J. Biol. 2013, 8, 591–599. [Google Scholar] [CrossRef]
- Steingroewer, J.; Bley, T.; Georgiev, V.; Ivanov, I.; Lenk, F.; Marchev, A.; Pavlov, A. Bioprocessing of differentiated plant in vitro systems. Eng. Life Sci. 2013, 13, 26–38. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Kutchan, T.M. The amaryllidaceae alkaloids: Biosynthesis and methods for enzyme discovery. Phytochem. Rev. 2016, 15, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Desgagné-Penix, I. Biosynthesis of alkaloids in Amaryllidaceae plants: A review. Phytochem. Rev. 2020. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chapter two—Chemodiversity, chemotaxonomy and chemoecology of amaryllidaceae alkaloids. In The alkaloids: Chemistry and Biology; Knölker, H.-J., Ed.; Academic Press: New York, NY, USA, 2020; Volume 83, pp. 113–185. [Google Scholar]
- Maeda, H.A. Evolutionary diversification of primary metabolism and its contribution to plant chemical diversity. Front. Plant Sci. 2019, 10, 881. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plan. 2010, 3, 2–20. [Google Scholar] [CrossRef]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From central to specialized metabolism: An overview of some secondary compounds derived from the primary metabolism for their role in conferring nutritional and organoleptic characteristics to fruit. Front. Plant Sci. 2019, 10, 835. [Google Scholar] [CrossRef]
- Li, W.; Yang, Y.; Qiao, C.; Zhang, G.; Luo, Y. Functional characterization of phenylalanine ammonia-lyase- and cinnamate 4-hydroxylase-encoding genes from lycoris radiata, a galanthamine-producing plant. Int. J. Biol. Macromol. 2018, 117, 1264–1279. [Google Scholar] [CrossRef]
- Hotchandani, T.; de Villers, J.; Desgagné-Penix, I. Developmental regulation of the expression of Amaryllidaceae alkaloid biosynthetic genes in Narcissus papyraceus. Genes 2019, 10, 594. [Google Scholar] [CrossRef]
- Singh, A.; Desgagné-Penix, I. Transcriptome and metabolome profiling of Narcissus pseudonarcissus ‘king alfred’ reveal components of Amaryllidaceae alkaloid metabolism. Sci. Rep. 2017, 7, 17356. [Google Scholar] [CrossRef]
- Singh, A.; Massicotte, M.-A.; Garand, A.; Tousignant, L.; Ouellette, V.; Bérubé, G.; Desgagné-Penix, I. Cloning and characterization of norbelladine synthase catalyzing the first committed reaction in Amaryllidaceae alkaloid biosynthesis. BMC Plant Biol. 2018, 18, 338. [Google Scholar] [CrossRef] [PubMed]
- Schenck, C.A.; Maeda, H.A. Tyrosine biosynthesis, metabolism, and catabolism in plants. Phytochemestry 2018, 149, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Han, X.; Xu, S.; Xia, B.; Jiang, Y.; Xue, Y.; Wang, R. Cloning and characterization of a tyrosine decarboxylase involved in the biosynthesis of galanthamine in Lycoris aurea. Peer J. 2019, 7, e6729. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Baek, S.-A.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of Lycoris radiata. Biology 2019, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Kilgore, M.B.; Holland, C.K.; Jez, J.M.; Kutchan, T.M. Identification of a noroxomaritidine reductase with Amaryllidaceae alkaloid biosynthesis related activities. J. Biol. Chem. 2016, 291, 16740–16752. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Qiao, C.; Pang, J.; Zhang, G.; Luo, Y. The versatile o-methyltransferase lromt catalyzes multiple o-methylation reactions in Amaryllidaceae alkaloids biosynthesis. Int. J. Biol. Macromol. 2019, 141, 680–692. [Google Scholar] [CrossRef]
- Sun, B.; Wang, P.; Wang, R.; Li, Y.; Xu, S. Molecular cloning and characterization of a meta/para-o-methyltransferase from Lycoris aurea. Int. J. Mol. Sci. 2018, 19, 1911. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Augustin, M.M.; May, G.D.; Crow, J.A.; Kutchan, T.M. Cyp96t1 of Narcissus sp. Aff. Pseudonarcissus catalyzes formation of the para-para’ c-c phenol couple in the Amaryllidaceae alkaloids. Front. Plant Sci. 2016, 7, 225. [Google Scholar] [CrossRef]
- Pyne, M.E.; Kevvai, K.; Grewal, P.S.; Narcross, L.; Choi, B.; Bourgeois, L.; Dueber, J.E.; Martin, V.J.J. A yeast platform for high-level synthesis of natural and unnatural tetrahydroisoquinoline alkaloids. bioRxiv 2019. [Google Scholar] [CrossRef]
- Ehrenworth, A.M.; Peralta-Yahya, P. Accelerating the semisynthesis of alkaloid-based drugs through metabolic engineering. Nat. Chem. Biol. 2017, 13, 249–258. [Google Scholar] [CrossRef]
- Diamond, A.; Desgagné-Penix, I. Metabolic engineering for the production of plant isoquinoline alkaloids. Plant Biotechn. J. 2016, 14, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Endoma-Arias, M.A.A.; Hudlicky, T. Chemoenzymatic total synthesis of (+)-galanthamine and (+)-narwedine from phenethyl acetate. Chem. A Eur. J. 2016, 22, 14540–14543. [Google Scholar] [CrossRef]
- van Otterlo, W.A.L.; Green, I.R. A review on recent syntheses of Amaryllidaceae alkaloids and isocarbostyrils (time period mid-2016 to 2017). Nat. Prod. Commu. 2018, 13, 255–277. [Google Scholar] [CrossRef]
- Ptak, A.; Moranska, E.; Skrzypek, E.; Warchoł, M.; Spina, R.; Laurain-Mattar, D.; Simlat, M. Carbohydrates stimulated Amaryllidaceae alkaloids biosynthesis in Leucojum aestivum L. plants cultured in RITA bioreactor. Peer J. 2020, 8, e8688. [Google Scholar] [CrossRef] [PubMed]
- Ptak, A.; Simlat, M.; Morańska, E.; Skrzypek, E.; Warchoł, M.; Tarakemeh, A.; Laurain-Mattar, D. Exogenous melatonin stimulated Amaryllidaceae alkaloid biosynthesis in in vitro cultures of Leucojum aestivum L. Ind. Crop. Prod. 2019, 138, 111458. [Google Scholar] [CrossRef]
- Saliba, S.; Ptak, A.; Boisbrun, M.; Spina, R.; Dupire, F.; Laurain-Mattar, D. Stimulating effect of both 4′-O-methylnorbelladine feeding and temporary immersion conditions on galanthamine and lycorine production by Leucojum aestivum L. bulblets. Eng. Life Sci. 2016, 16, 731–739. [Google Scholar] [CrossRef]
- Akita, M.; Ohta, Y. A simple bioreactor system for production of storage organs of Chinese yam (Dioscorea opposite Thumb.). Plant. Biotech. 2002, 9, 353–356. [Google Scholar] [CrossRef]
- Firoozabady, E.; Gutterson, N. Cost-effective in vitro propagation methods for pineapple. Plant Cell Rep. 2003, 21, 844–850. [Google Scholar] [CrossRef]
- Bondarev, N.; Reshetnyak, O.; Nosov, A. Features of development of Stevia rebaudiana shoots cultivated in the roller bioreactor and their production of steviol glycosides. Plant. Med. 2002, 68, 759–762. [Google Scholar] [CrossRef]
- Paek, K.-Y.; Hahn, E.-J.; Son, S.-H. Application of bioreactors for large-scale micropropagation systems of plants. Vitro Cell Dev Plant. 2001, 37, 149–157. [Google Scholar] [CrossRef]
- Paek, K.Y.; Chakrabarty, D.; Hahn, E.J. Application of bioreactor systems for large scale production of horticultural and medicinal plants. Plant. Cell Tissue Org. Cult. 2005, 81, 287–300. [Google Scholar] [CrossRef]
- Ziv, M. Simple bioreactors for mass propagation of plants. Plant. Cell Tissue Org. Cult. 2005, 81, 277–285. [Google Scholar] [CrossRef]
- Jo, U.; Murthy, H.; Hahn, E.; Paek, K. Micropropagation of Alocasia amazonica using semisolid and liquid cultures. Vitro Cell Dev. Plant. 2008, 44, 26–32. [Google Scholar] [CrossRef]
- Liu, C.-Z.; Guo, C.; Wang, Y.-C.; Ouyang, F. Comparison of various bioreactors on growth and artemisinin biosynthesis of Artemisia annua L. shoot cultures. Process. Biochem. 2003, 39, 45–49. [Google Scholar] [CrossRef]
- Takayama, S.; Akita, M. The types of bioreactors used for shoots and embryos. Plant. Cell Tissue Org. Cult. 1994, 39, 147–156. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Type of In Vitro Systems | Amaryllidaceae Alkaloids | Accumulated Concentrations | References |
|---|---|---|---|---|
| Narcissus pseuddonarcissus cv. Carlton | callus shoots bulbs | Galanthamine Galanthamine Galanthamine | 0–7 µg/g FW 40–130 µg/g FW 10–215 µg/g FW | [22] |
| Narcissus tazetta var. Meskin | callus | Galanthamine Lycorine | 0.5–1.9 µg/g DW 1.2–21.5 µg/g DW | [38] |
| bulblet | Galanthamine Lycorine | 15–80 µg/g DW 731–1900 µg/g DW | ||
| roots | Galanthamine Lycorine | 5.2 µg/g DW 131 µg/g DW | [39] | |
| Narcissus confusus | shoots | Galanthamine Tazettine Haemanthamine | 13.07 mg/g DW 0.75 mg/g DW 3.16 mg/g DW | [40] |
| Hymenocallis littoralis | callus | Lycorine | 0.1–2.6 µg/g extract | [41] |
| Rhodophiala bifida | roots | Montanine | 1.19 mg/g | [42] |
| bulbs | Montanine | 2.21 mg/g | ||
| leaves | Montanine | 2.10 mg/g | ||
| Pancratium maritimum | shoots | Lycorine | 2.90 mg/g DW | [23] |
| shoots | Haemanthamine Lycorine | 900 µg/g DW 800 µg/g DW | [43] | |
| Leucojum aestivum | calli | Galanthamine Lycorine | 10–13 µg/g DW 10–30µg/g DW | [36,44] |
| shoots | Galanthamine Lycorine | 15–454 µg/g DW 100 µg/g DW | [36,44,45] | |
| Crinum moorei | bulbs | Crinamidine Crinine Powelline Undulatine | 130–200 mg/100g DW 25–60 mg/100g DW 25–50 mg/100g DW 25–80 mg/100g DW | [46] |
| Hippeastrum vittatum | bulbs | Montanine Pancracine 11-Hydroxyvitattine Vitattine | Data not presented | [47] |
| Hippeastrum goianum | bulbs | Lycorine | 0.075–0.125 µg/mL | [48] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgiev, V.; Ivanov, I.; Pavlov, A. Recent Progress in Amaryllidaceae Biotechnology. Molecules 2020, 25, 4670. https://doi.org/10.3390/molecules25204670
Georgiev V, Ivanov I, Pavlov A. Recent Progress in Amaryllidaceae Biotechnology. Molecules. 2020; 25(20):4670. https://doi.org/10.3390/molecules25204670
Chicago/Turabian StyleGeorgiev, Vasil, Ivan Ivanov, and Atanas Pavlov. 2020. "Recent Progress in Amaryllidaceae Biotechnology" Molecules 25, no. 20: 4670. https://doi.org/10.3390/molecules25204670
APA StyleGeorgiev, V., Ivanov, I., & Pavlov, A. (2020). Recent Progress in Amaryllidaceae Biotechnology. Molecules, 25(20), 4670. https://doi.org/10.3390/molecules25204670