
New Real-Time PCRs to Differentiate Rickettsia spp. and Rickettsia conorii

 ,
,
Abstract
1. Introduction
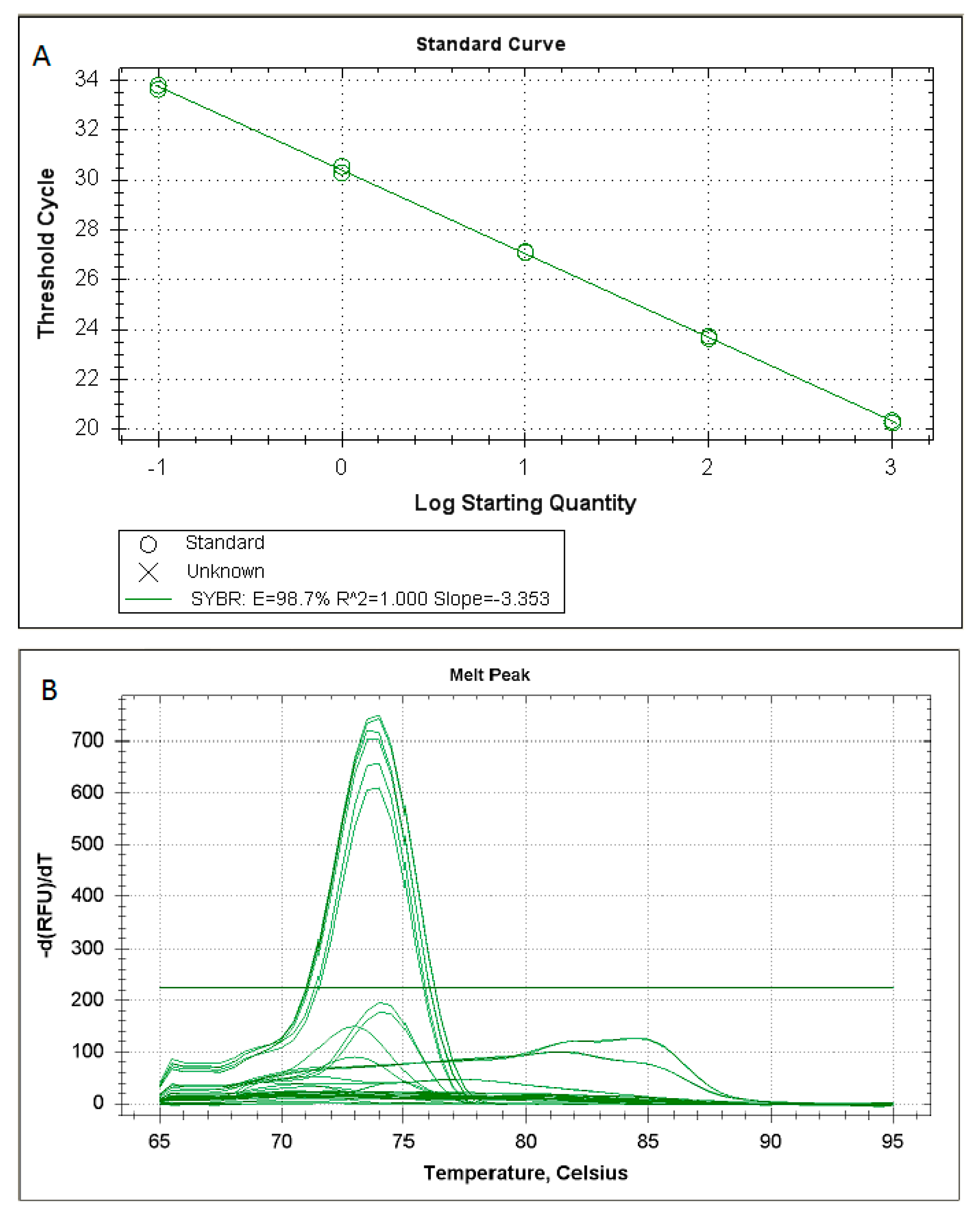
2. Results
3. Discussion
4. Materials and Methods
4.1. Rickettsia Conorii Malish 7 Strain Culture
4.2. Field Samples for Specificity Analyses
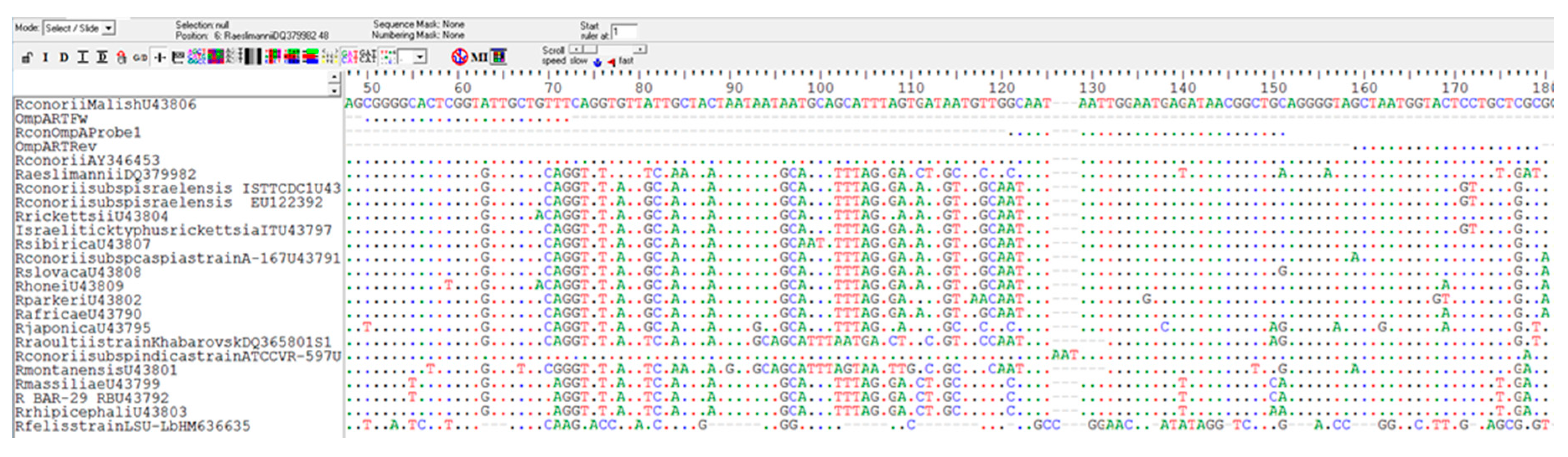
4.3. Primer Design and Selection
4.4. Cloning and Sequencing of PCR Products
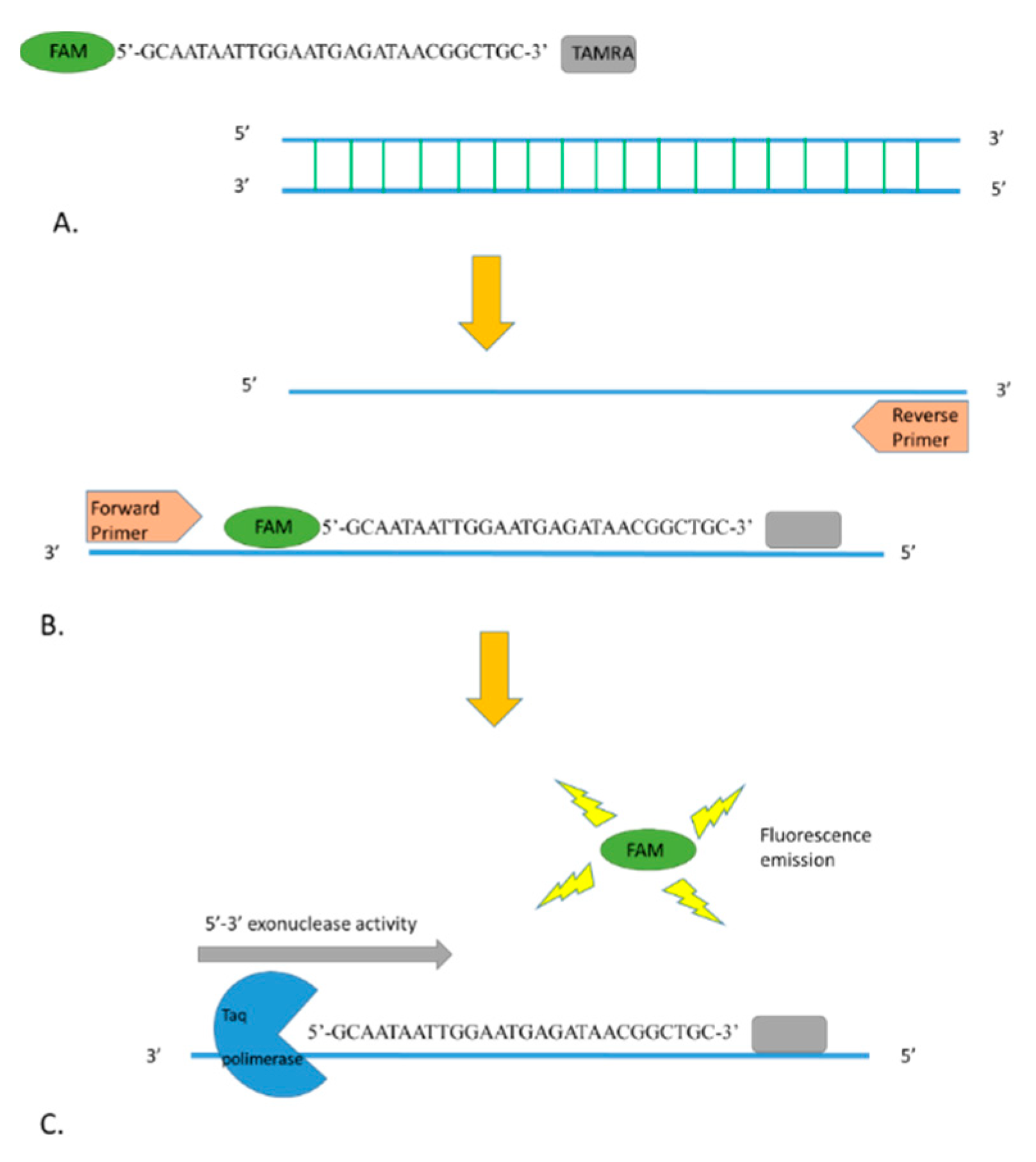
4.5. Real-Time PCR Optimization
4.6. Specificity and Sensitivity of the New Assays
Author Contributions
Funding
Conflicts of Interest
References
- Portillo, A.; de Sousa, R.; Santibáñez, S.; Duarte, A.; Oteo, J.A. Guidelines for the Detection of Rickettsia spp. Vector-Borne Zoonotic Dis. 2017, 17, 23–32. [Google Scholar] [CrossRef]
- Merhej, V.; Angelakis, E.; Socolovschi, C.; Raoult, D. Genotyping, evolution and epidemiological findings of Rickettsia species. Infect. Genet. Evol. 2014, 25, 122–137. [Google Scholar] [CrossRef]
- Fernández de Mera, I.G.; Blanda, V.; Torina, A.; Dabaja, M.F.; Ali, E.R.; Cabezas-Cruz, A.; de la Fuente, J. Identification and molecular characterization of spotted fever group rickettsiae in ticks collected from farm ruminants in Lebanon. Ticks Tick Borne Dis. 2018, 9, 104–108. [Google Scholar] [CrossRef]
- Oteo, J.A.; Portillo, A. Tick-borne rickettsioses in Europe. Ticks Tick Borne Dis. 2012, 3, 271–278. [Google Scholar] [CrossRef]
- Dieme, C.; Bechah, Y.; Socolovschi, C.; Audoly, G.; Faye, O. Transmission potential of Rickettsia felis infection by Anopheles gambiae mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 8088–8093. [Google Scholar] [CrossRef]
- Adem, P.V. Emerging and re-emerging rickettsial infections. Semin Diagn Pathol. 2019, 36, 146–151. [Google Scholar] [CrossRef]
- Merhej, V.; Raoult, D. Rickettsial evolution in the light of comparative genomics. Biol. Rev. Camb. Philos. Soc. 2011, 86, 379–405. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Raoult, D. Tick-Borne Rickettsioses around the World: Emerging Diseases Challenging Old Concepts. Clin. Microbiol. Rev. 2005, 18, 719–756. [Google Scholar] [CrossRef]
- Parola, P.; Socolovschi, C.; Raoult, D. Deciphering the relationships between Rickettsia conorii conorii and Rhipicephalus sanguineus in the ecology and epidemiology of Mediterranean spotted fever. Ann. N. Y. Acad. Sci. 2009, 1166, 49–54. [Google Scholar] [CrossRef]
- Duque, V.; Ventura, C.; Seixas, D.; da Cunha, S.; Duque, V.; Melico-Silvestre, A.; Barai, A.; Mendonca, N.; Martins, J. Mediterranean spotted fever and encephalitis: A case report and review of the literature. J. Infect. Chemother. 2012, 18, 105–108. [Google Scholar] [CrossRef]
- Blanda, V.; Torina, A.; La Russa, F.; D’Agostino, R.; Randazzo, K.; Scimeca, S.; Giudice, E.; Caracappa, S.; Cascio, A.; de la Fuente, J. A retrospective study of the characterization of Rickettsia species in ticks collected from humans. Ticks Tick Borne Dis. 2017, 8, 610–614. [Google Scholar] [CrossRef]
- Parola, P.; Socolovschi, C.; Jeanjean, L.; Bitam, I.; Fournier, P.-E.; Sotto, A.; Labauge, P.; Raoult, D. Warmer weather linked to tick attack and emergence of severe rickettsioses. PLoS Negl. Trop. Dis. 2008, 2, e338. [Google Scholar] [CrossRef]
- Torina, A.; Villari, S.; Blanda, V.; Vullo, S.; La Manna, M.P.; Azgomi, M.S.; Liberto, D.D.; de la Fuente, J.; Sireci, G. Innate Immune Response to Tick-Borne Pathogens: Cellular and Molecular Mechanisms Induced in the Hosts. Int. J. Mol. Sci. 2020, 21, 5437. [Google Scholar] [CrossRef]
- Rovery, C.; Brouqui, P.; Raoult, D. Questions on Mediterranean spotted fever a century after its discovery. Emerg. Infect. Dis. 2008, 14, 1360–1367. [Google Scholar] [CrossRef]
- Raoult, D.; Weiller, P.J.; Chagnon, A.; Chaudet, H.; Gallais, H.; Casanova, P. Mediterranean spotted fever: Clinical, laboratory and epidemiological features of 199 cases. Am. J. Trop. Med. Hyg. 1986, 35, 845–850. [Google Scholar] [CrossRef]
- Giammanco, G.M.; Vitale, G.; Mansueto, S.; Capra, G.; Caleca, M.P.; Ammatuna, P. Presence of Rickettsia conorii subsp. israelensis, the causative agent of Israeli spotted fever, in Sicily, Italy, ascertained in a retrospective study. J. Clin. Microbiol. 2005, 43, 6027–6031. [Google Scholar] [CrossRef]
- Torina, A.; Fernández de Mera, I.G.; Alongi, A.; Mangold, A.J.; Blanda, V.; Scarlata, F.; Di Marco, V.; de la Fuente, J. Rickettsia conorii Indian tick typhus strain and R. slovaca in humans, Sicily. Emerg. Infect. Dis. 2012, 18, 1008–1010. [Google Scholar] [CrossRef]
- Cascio, A.; Torina, A.; Valenzise, M.; Blanda, V.; Camarda, N.; Bombaci, S.; Iaria, C.; De Luca, F.; Wasniewska, M. Scalp eschar and neck lymphadenopathy caused by Rickettsia massiliae. Emerg. Infect Dis. 2013, 19, 836–837. [Google Scholar] [CrossRef]
- Kernif, T.; Leulmi, H.; Raoult, D.; Parola, P. Emerging Tick-Borne Bacterial Pathogens. Microbiol. Spectr. 2016, 4, 295–310. [Google Scholar] [CrossRef]
- Ciceroni, L.; Pinto, A.; Ciarrocchi, S.; Ciervo, A. Current knowledge of rickettsial diseases in Italy. Ann. N. Y. Acad. Sci. 2006, 1078, 143–149. [Google Scholar] [CrossRef]
- Gomez-Barroso, D.; Vescio, M.F.; Bella, A.; Ciervo, A.; Busani, L.; Rizzo, C.; Rezzo, G.; Pezzotti, P. Mediterranean spotted fever rickettsiosis in Italy, 2001–2015: Spatio-temporal distribution based on hospitalization records. Ticks Tick Borne Dis. 2019, 10, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, M.G.; Caprì, A.; Solano-Gallego, L.; Lombardo, G.; Torina, A.; Masucci, M. Prevalence of antibodies against Rickettsia conorii, Babesia canis, Ehrlichia canis, and Anaplasma phagocytophilum antigens in dogs from the Stretto di Messina area (Italy). Ticks Tick Borne Dis. 2012, 3, 315–318. [Google Scholar] [CrossRef]
- Kidd, L.; Maggi, R.; Diniz, P.P.V.P.; Hegarty, B.; Tucker, M.; Breitschwerdt, E. Evaluation of conventional and real-time PCR assays for detection and differentiation of Spotted Fever Group Rickettsia in dog blood. Vet. Microbiol. 2008, 129, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Stenos, J.; Graves, S.R.; Unsworth, N.B. A highly sensitive and specific real-time PCR assay for the detection of spotted fever and typhus group Rickettsiae. Am. J. Trop Med. Hyg. 2005, 73, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Segura, F.; Pons, I.; Sanfeliu, I.; Nogueras, M.M. Shell-vial culture, coupled with real-time PCR, applied to Rickettsia conorii and Rickettsia massiliae-Bar29 detection, improving the diagnosis of the Mediterranean spotted fever. Ticks Tick Borne Dis. 2016, 7, 457–461. [Google Scholar] [CrossRef]
- Fournier, P.E.; Dumler, J.S.; Greub, G.; Zhang, J.; Wu, Y.; Raoult, D. Gene sequence-based criteria for identification of new rickettsia isolates and description of Rickettsia heilonjiangensis sp. Nov. J. Clin. Microbiol. 2003, 41, 5456–5465. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Raoult, D. Identification of rickettsial isolates at the species level using multi-spacer typing. BMC Microbiol. 2007, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, M.E.; Bosserman, E.A.; Demma, L.J.; Zambrano, M.L.; Blau, D.M.; Dasch, G.A. Isolation and identification of Rickettsia massiliae from Rhipicephalus sanguineus ticks collected in Arizona. Appl. Environ. Microbiol. 2006, 72, 5569–5577. [Google Scholar] [CrossRef]
- Scarpulla, M.; Barlozzari, G.; Marcario, A.; Salvato, L.; Blanda, V.; De Liberato, C.; D’Agostini, C.; Torina, A.; Macri, G. Molecular detection and characterization of spotted fever group rickettsiae in ticks from Central Italy. Ticks Tick Borne Dis. 2016, 7, 1052–1056. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Segal, E.; Rubinstein, E. Glucose-6-phosphate dehydrogenase deficiency: Possible determinant for a fulminant course of Israeli spotted fever. Isr. Med. Assoc. J. 2000, 2, 781–782. [Google Scholar]
- Beati, L.; Roux, V.; Ortuno, A.; Castella, J.; Segura Porta, F.; Raoult, D. Phenotypic and genotypic characterization of spotted fever group rickettsiae isolated from Catalan Rhipicephalus sanguineus ticks. J. Clin. Microbiol. 1996, 34, 2688–2694. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.M.; Maurin, M.; Vestris, G.; Raoult, D. In vitro susceptibilities of 27 rickettsiae to 13 antimicrobials. Antimicrob. Agents Chemother. 1998, 42, 1537–1541. [Google Scholar] [CrossRef] [PubMed]
- Vanrompay, D.; Nguyen, T.L.A.; Cutler, S.J.; Butaye, P. Antimicrobial Resistance in Chlamydiales, Rickettsia, Coxiella, and Other Intracellular Pathogens. Microbiol. Spectr. 2018, 6, 485–500. [Google Scholar] [CrossRef]
- Rolain, J.M. Antimicrobial susceptibility of rickettsial agents. In Rickettsial Diseases; Raoult, D., Parola, P., Eds.; Informa Healthcare: New York, NY, USA, 2007; pp. 361–369. [Google Scholar] [CrossRef]
- Vitorino, L.; Chelo, I.M.; Bacellar, F.; Zé-Zé, L. Rickettsiae phylogeny: A multigenic approach. Microbiology 2007, 153, 160–168. [Google Scholar] [CrossRef]
- Leitner, M.; Yitzhaki, S.; Rzotkiewicz, S.; Keysary, A. Polymerase chain reaction-based diagnosis of Mediterranean spotted fever in serum and tissue samples. Am. J. Trop. Med. Hyg. 2002, 67, 166–169. [Google Scholar] [CrossRef]
- Paris, D.H.; Blacksell, S.D.; Stenos, J.; Graves, S.R.; Unsworth, N.B.; Phetsouvanh, R.; Newton, P.N.; Day, N.P.J. Real-time multiplex PCR assay for detection and differentiation of rickettsiae and orientiae. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 186–193. [Google Scholar] [CrossRef]
- Giulieri, S.; Jaton, K.; Cometta, A.; Trellu, L.T.; Greub, G. Development of a duplex real-time PCR for the detection of Rickettsia spp. and typhus group rickettsia in clinical samples. FEMS Immunol. Med. Microbiol. 2012, 64, 92–97. [Google Scholar] [CrossRef]
- Kato, C.Y.; Chung, I.H.; Robinson, L.K.; Austin, A.L.; Dasch, G.A.; Massung, R.F. Assessment of real-time PCR assay for detection of Rickettsia spp. and Rickettsia rickettsii in banked clinical samples. J. Clin. Microbiol. 2013, 51, 314–317. [Google Scholar] [CrossRef]
- Denison, A.M.; Amin, B.D.; Nicholson, W.L.; Paddock, C.D. Detection of Rickettsia rickettsii, Rickettsia parkeri, and Rickettsia akari in skin biopsy specimens using a multiplex real-time polymerase chain reaction assay. Clin. Infect. Dis. 2014, 59, 635–642. [Google Scholar] [CrossRef]
- Tomassone, L.; De Meneghi, D.; Adakal, H.; Rodighiero, P.; Pressi, G.; Grego, E. Detection of Rickettsia aeschlimannii and Rickettsia africae in ixodid ticks from Burkina Faso and Somali Region of Ethiopia by new real-time PCR assays. Ticks Tick Borne Dis. 2016, 7, 1082–1088. [Google Scholar] [CrossRef]
- Jiang, J.; You, B.J.; Liu, E.; Apte, A.; Yarina, T.R.; Myers, T.E.; Lee, J.S.; Francesconi, S.C.; O’Guinn, M.L.; Tsertsvadze, N.; et al. Development of three quantitative real-time PCR assays for the detection of Rickettsia raoultii, Rickettsia slovaca, and Rickettsia aeschlimannii and their validation with ticks from the country of Georgia and the Republic of Azerbaijan. Ticks Tick Borne Dis. 2012, 3, 327–331. [Google Scholar] [CrossRef]
- Roux, V.; Fournier, P.E.; Raoult, D. Differentiation of spottedfever group Rickettsiae by sequencing and analysis of restriction fragment length polymorphism of PCR-amplified DNA of the gene encoding the protein rOmpA. J. Clin. Microbiol. 1996, 34, 2058–2065. [Google Scholar] [CrossRef]
- Giudice, E.; Di Pietro, S.; Alaimo, A.; Blanda, V.; Lelli, R.; Francaviglia, F.; Caracappa, S.; Torina, A. A molecular survey of Rickettsia felis in fleas from cats and dogs in Sicily (Southern Italy). PLoS ONE 2014, 9, e106820. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach [published correction appears in Clin Microbiol Rev. 2014 Jan;27(1):166]. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef]
- Holland, C.A.; Kiechle, F.L. Point-of-care molecular diagnostic systems-past, present and future. Curr. Opin. Microbiol. 2005, 8, 504–509. [Google Scholar] [CrossRef]
- Renvoisé, A.; Rolain, J.M.; Socolovschi, C.; Raoult, D. Widespread use of real-time PCR for rickettsial diagnosis. FEMS Immunol. Med. Microbiol. 2012, 64, 126–129. [Google Scholar] [CrossRef]
- Oteo, J.A.; Portillo, A.; Santibáñez, S.; Blanco, J.R.; Pérez-Martínez, L.; Ibarra, V. Cluster of cases of human Rickettsia felis infection from Southern Europe (Spain) diagnosed by PCR. J. Clin. Microbiol. 2006, 44, 2669–2671. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, S.H.; Park, K.H.; Koh, Y.S.; Lee, K.H.; Baik, H.S.; Choi, M.S.; Kim, I.S.; Jang, W.J. Evaluation of PCR-based assay for diagnosis of spotted fever group rickettsiosis in human serum samples. Clin. Diagn. Lab. Immunol. 2005, 12, 759–763. [Google Scholar] [CrossRef][Green Version]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef]
- Torina, A.; Agnone, A.; Blanda, V.; Alongi, A.; D’Agostino, R.; Caracappa, S.; Marino, A.M.F.; Marco, V.D.; La Fuente, J. Development and validation of two PCR tests for the detection of and differentiation between Anaplasma ovis and Anaplasma marginale. Ticks Tick Borne Dis. 2012, 3, 283–287. [Google Scholar] [CrossRef]
- Naranjo, V.; Ruiz-Fons, F.; Höfle, U.; De Mera, I.G.F.; Villanua, D.; Almazan, C.; Torina, A.; Caracappa, S.; Kocan, K.M.; Gortazar, C.; et al. Molecular epidemiology of human and bovine anaplasmosis in southern Europe. Ann. N. Y. Acad. Sci. 2006, 1078, 95–99. [Google Scholar] [CrossRef]
- Stuen, S.; Nevland, S.; Moum, T. Fatal cases of Tick-borne fever (TBF) in sheep caused by several 16S rRNA gene variants of Anaplasma phagocytophilum. Ann. N. Y. Acad. Sci. 2003, 990, 433–434. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Madigan, J.E.; Dumler, J.S.; Goodman, J.L.; Hayes, S.F.; Barlough, J.E.; Nelson, C.M.; Kurtti, T.J. Isolation of the equine granulocytic ehrlichiosis agent, Ehrlichia equi, in tick cell culture. J. Clin. Microbiol. 1996, 34, 664–670. [Google Scholar] [CrossRef]
- Gubbels, J.M.; de Vos, A.P.; van der Weide, M.; Viseras, J.; Schouls, L.M.; De Vries, E.; Jongejan, F. Simultaneous detection of bovine Theileria and Babesia species by reverse line blot hybridization. J. Clin. Microbiol. 1999, 37, 1782–1789. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds in this study are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rickettsia spp. RT-PCR (ompB) | R. conorii RT-PCR (ompA) | |
|---|---|---|
| R. conorii | + | + |
| R. aeschlimannii | + | - |
| R. massiliae | + | - |
| R. raoultii | + | - |
| R. monacensis | + | - |
| R. helvetica | + | - |
| R. slovaca | + | - |
| R. felis | + | - |
| Rickettsia endosymbiont of Haemaphysalis sulcata | + | - |
| Candidatus Rickettsia hoogstraalii | + | - |
| Babesia bigemina | - | - |
| Babesia bovis | - | - |
| Theieleria annulata | - | - |
| Ehrlichia canis | - | - |
| Anaplasma marginale | - | - |
| Anaplasma ovis | - | - |
| Anaplasma phagocytophilum | - | - |
| Rickettsia Species | ompB GenBank Acc. n. | ompA GenBank Acc. n. |
|---|---|---|
| R. conorii conorii | AF123721.1 | U43806 |
| R. conorii Indian tick typhus | AF123726.1 | 597U437 |
| R. conorii subsp. Caspia | AY643093.1 | 167U4379 |
| R. conorii Israeli tick typhus | AF123712.1 | U43797 |
| R. massiliae | AF123714.1 | U43799 |
| R. raoultii | DQ365798.1 | DQ365801S1 |
| R. aeschlimannii | AF123705.1 | DQ379982 |
| R. slovaca | AF123723.2 | U43808 |
| R. monacensis | EF380356.1 | FJ919650 |
| R. helvetica | AF123725.1 | |
| R. rhipicephali | AF123719.1 | U43803 |
| R. felis | GQ385243 | HM636635 |
| Candidatus R. hoogstraalii | EF629536.1 | |
| Rickettsia endosimbiont of I. scapularis | EF433951.1 | |
| R. mongolotimonae | DQ097083.1 | DQ097082.1 |
| R. sibirica | AF123722.1 | U43807 |
| R. africae | AF123706.1 | U43790 |
| R. honei | AF123711.1 | U43809 |
| R. rickettsii | X16353.1 | U43804 |
| R. montanensis | AF123716.1 | U43801 |
| R. parkeri | AF123717.1 | U43802 |
| R. japonica | AB003681.1 | U43795 |
| R. prowazekii | DQ926859.1 | |
| R. australis | AF123709.1 | |
| R. heilongjiangensis | AY260451.1 | |
| R BAR-29 | RBU43792 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanda, V.; D’Agostino, R.; Giudice, E.; Randazzo, K.; La Russa, F.; Villari, S.; Vullo, S.; Torina, A. New Real-Time PCRs to Differentiate Rickettsia spp. and Rickettsia conorii. Molecules 2020, 25, 4431. https://doi.org/10.3390/molecules25194431
Blanda V, D’Agostino R, Giudice E, Randazzo K, La Russa F, Villari S, Vullo S, Torina A. New Real-Time PCRs to Differentiate Rickettsia spp. and Rickettsia conorii. Molecules. 2020; 25(19):4431. https://doi.org/10.3390/molecules25194431
Chicago/Turabian StyleBlanda, Valeria, Rosalia D’Agostino, Elisabetta Giudice, Kety Randazzo, Francesco La Russa, Sara Villari, Stefano Vullo, and Alessandra Torina. 2020. "New Real-Time PCRs to Differentiate Rickettsia spp. and Rickettsia conorii" Molecules 25, no. 19: 4431. https://doi.org/10.3390/molecules25194431
APA StyleBlanda, V., D’Agostino, R., Giudice, E., Randazzo, K., La Russa, F., Villari, S., Vullo, S., & Torina, A. (2020). New Real-Time PCRs to Differentiate Rickettsia spp. and Rickettsia conorii. Molecules, 25(19), 4431. https://doi.org/10.3390/molecules25194431