
The Role of Circulating Lycopene in Low-Grade Chronic Inflammation: A Systematic Review of the Literature


Abstract
1. Introduction
1.1. Low-Grade Chronic Inflammation
1.2. Lycopene
1.2.1. Physicochemical Properties of Lycopene
1.2.2. Lycopene Kinetics after Oral Administration: Absorption, Distribution, Metabolism, Excretion
1.2.3. Mechanism of Action (In Vitro)
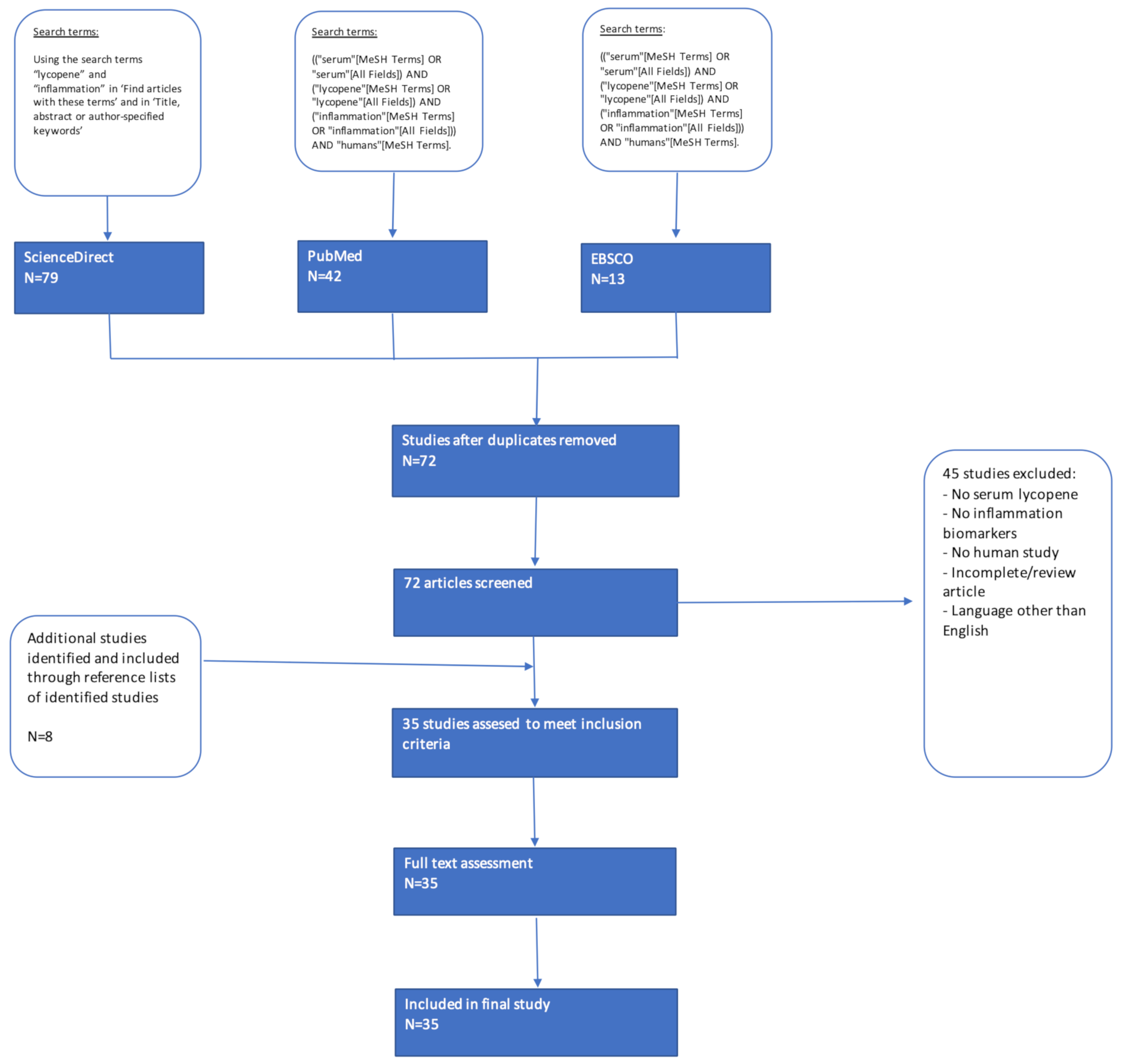
2. Methods
2.1. Literature Search
2.2. Application of Inclusion/Exclusion Criteria
2.3. Data Extraction and Analysis
3. Results
3.1. Study Characteristics
3.2. Cross-Sectional Studies
3.3. Intervention Studies
4. Discussion
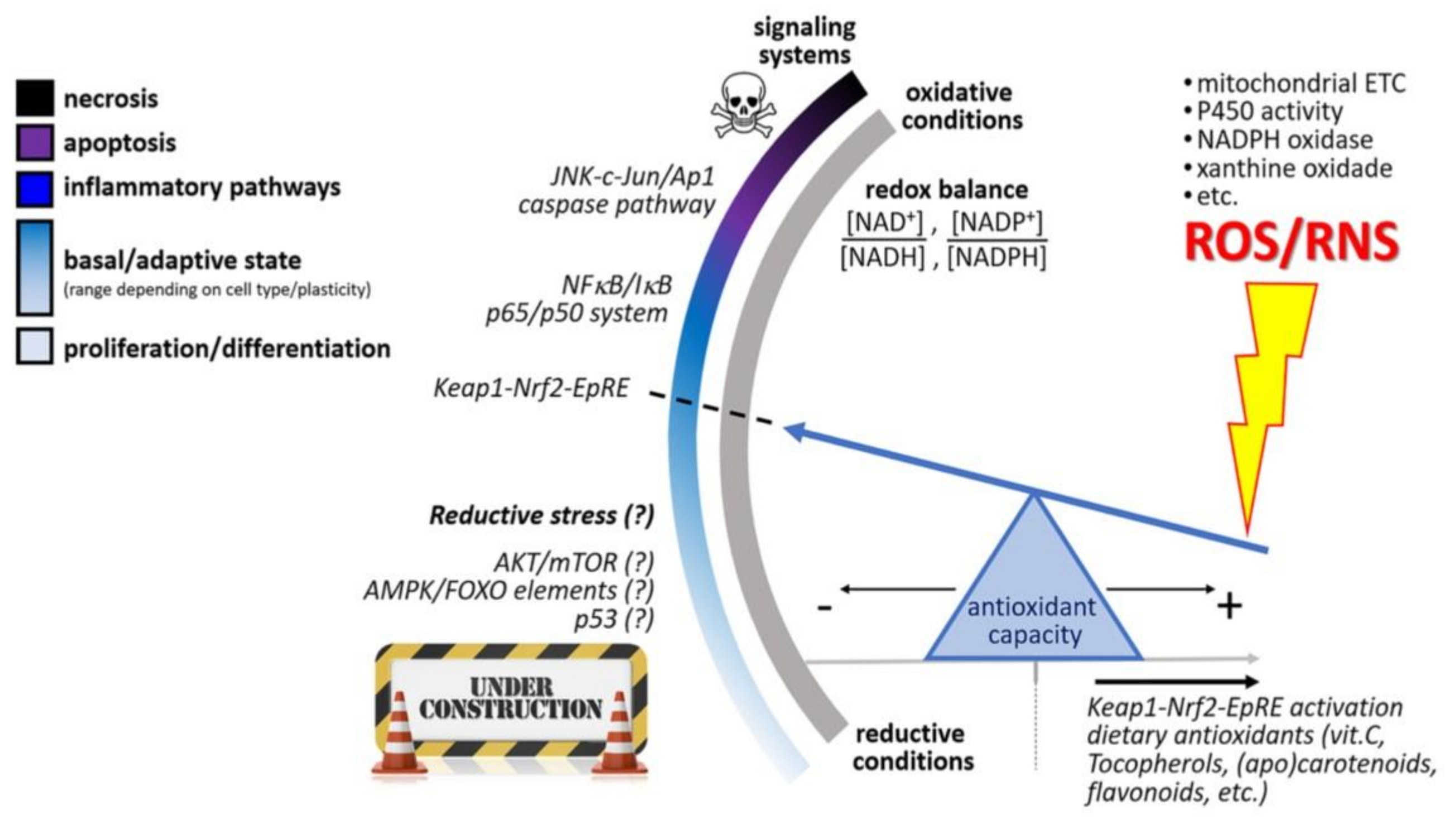
4.1. Molecular Mechanisms of Action
4.2. Dietary Recommendations
4.3. Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Georgiou, N.; Garssen, J.; Witkamp, R. Pharma–nutrition interface: The gap is narrowing. Eur. J. Pharmacol. 2011, 651, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bagetta, D.; Maruca, A.; Lupia, A.; Mesiti, F.; Catalano, R.; Romeo, I.; Moraca, F.; Ambrosio, F.; Costa, G.; Artese, A.; et al. Mediterranean products as promising source of multi-target agents in the treatment of metabolic syndrome. Eur. J. Med. Chem. 2020, 186, 111903. [Google Scholar] [CrossRef] [PubMed]
- Kashi, D.; Shabir, A.; Da Boit, M.; Bailey, S.; Higgins, M. The Efficacy of Administering Fruit-Derived Polyphenols to Improve Health Biomarkers, Exercise Performance and Related Physiological Responses. Nutrients 2019, 11, 2389. [Google Scholar] [CrossRef]
- Maria, L.; Eamon, E.; Mana, S.; Rosa, C. Relation of Fruits and Vegetables with Major Cardiometabolic Risk Factors, Markers of Oxidation, and Inflammation. Nutrients 2019, 11, 2381. [Google Scholar]
- Bosma-den Boer, M.; van Wetten, M.; Pruimboom, L. Chronic inflammatory diseases are stimulated by current lifestyle: How diet, stress levels and medication prevent our body from recovering. Nutr. Metab. 2012, 9, 32. [Google Scholar] [CrossRef]
- de Boer, A.; van de Worp, W.; Hageman, G.; Bast, A. The effect of dietary components on inflammatory lung diseases–a literature review. Int. J. Food Sci. Nutr. 2017, 68, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Núñez, B.; Pruimboom, L.; Dijck-Brouwer, D.; Muskiet, F. Lifestyle and nutritional imbalances associated with Western diseases: Causes and consequences of chronic systemic low-grade inflammation in an evolutionary context. J. Nutr. Biochem. 2013, 24, 1183–1201. [Google Scholar] [CrossRef] [PubMed]
- Todoric, J.; Antonucci, L.; Karin, M. Targeting Inflammation in Cancer Prevention and Therapy. Cancer Prev. Res. 2016, 9, 895–905. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New Adventures of an Old Flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Calder, P.; Ahluwalia, N.; Albers, R.; Bosco, N.; Bourdet-Sicard, R.; Haller, D.; Holgate, S.; Jönsson, L.; Latulippe, M.; Marcos, A.; et al. A Consideration of Biomarkers to be Used for Evaluation of Inflammation in Human Nutritional Studies. Br. J. Nutr. 2013, 109, S1–S34. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Del Giudice, M.; Gangestad, S. Rethinking IL-6 and CRP: Why they are more than inflammatory biomarkers, and why it matters. Brain Behav. Immun. 2018, 70, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.; Vinoy, S.; Russell, W.; Baka, A.; Roche, H.; Tuohy, K.; Teeling, J.; Blaak, E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Lacourt, T.; Vichaya, E.; Chiu, G.; Dantzer, R.; Heijnen, C. The High Costs of Low-Grade Inflammation: Persistent Fatigue as a Consequence of Reduced Cellular-Energy Availability and Non-adaptive Energy Expenditure. Front. Behav. Neurosci. 2018, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Abrams, N.; Carrick, D.; Chander, P.; Dwyer, J.; Hamlet, M.; Macchiarini, F.; PrabhuDas, M.; Shen, G.; Tandon, P.; et al. Biomarkers of chronic inflammation in disease development and prevention: Challenges and opportunities. Nat. Immunol. 2017, 18, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.; Cruz, M.; Gualillo, O. Editorial: The Physiology of Inflammation—The Final Common Pathway to Disease. Front. Physiol. 2018, 9, 1741. [Google Scholar] [CrossRef]
- Kaluza, J.; Håkansson, N.; Harris, H.; Orsini, N.; Michaëlsson, K.; Wolk, A. Influence of anti-inflammatory diet and smoking on mortality and survival in men and women: Two prospective cohort studies. J. Int. Med. 2018, 285, 75–91. [Google Scholar] [CrossRef]
- Khoo, H.; Prasad, K.; Kong, K.; Jiang, Y.; Ismail, A. Carotenoids and Their Isomers: Color Pigments in Fruits and Vegetables. Molecules 2011, 16, 1710–1738. [Google Scholar] [CrossRef]
- Schieber, A.; Carle, R. Occurrence of carotenoid cis-isomers in food: Technological, analytical, and nutritional implications. Trends Food Sci. Technol. 2005, 16, 416–422. [Google Scholar] [CrossRef]
- Rao, A.; Ray, M.; Rao, L. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164. [Google Scholar]
- Palozza, P.; Parrone, N.; Catalano, A.; Simone, R. Tomato Lycopene and Inflammatory Cascade: Basic Interactions and Clin.Implications. Curr. Med. Chem. 2010, 17, 2547–2563. [Google Scholar] [CrossRef] [PubMed]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Parker, R. Absorption, metabolism, and transport of carotenoids. FASEB J. 1996, 10, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Lycopene: A Biologically Important Carotenoid for Humans? Arch. Biochem. Biophys. 1996, 336, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, C.; Stahl, W.; Sies, H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am. J. Clin. Nutr. 1997, 66, 116–122. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.; Xu, X.; Viana, M.; Chen, L.; Stacewicz-Sapuntzakis, M.; Duncan, C.; Bowen, P.; Sharifi, R. Liquid Chromatography−Mass Spectrometry of cis and all-trans-Lycopene in Human Serum and Prostate Tissue after Dietary Supplementation with Tomato Sauce. J. Agric. Food Chem. 2002, 50, 2214–2219. [Google Scholar] [CrossRef]
- Zaripheh, S.; Erdman, J. The Biodistribution of a Single Oral Dose of [14C]-Lycopene in Rats Prefed Either a Control or Lycopene-Enriched Diet. J. Nutr. 2005, 135, 2212–2218. [Google Scholar] [CrossRef]
- Wang, X. Lycopene metabolism and its biological significance. Am. J. Clin. Nutr. 2012, 96, S1214–S1222. [Google Scholar] [CrossRef]
- Nagao, A. Oxidative Conversion of Carotenoids to Retinoids and Other Products. J. Nutr. 2004, 134, S237–S240. [Google Scholar] [CrossRef]
- Huang, C.; Fan, Y.; Lin, C.; Hu, M. Lycopene inhibits matrix metalloproteinase-9 expression and down-regulates the binding activity of nuclear factor-kappa B and stimulatory protein-1. J. Nutr. Biochem. 2007, 18, 449–456. [Google Scholar] [CrossRef]
- Fenni, S.; Hammou, H.; Astier, J.; Bonnet, L.; Karkeni, E.; Couturier, C.; Tourniaire, F.; Landrier, J. Lycopene and tomato powder supplementation similarly inhibit high-fat diet induced obesity, inflammatory response, and associated metabolic disorders. Mol. Nutr. Food Res. 2017, 61, 1601083. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Kim, W.; Ha, A.; Kim, M.; Chang, M. Anti-inflammatory effect of lycopene in SW480 human colorectal cancer cells. Nutr. Res. Pract. 2017, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Senkus, K.; Tan, L.; Crowe-White, K. Lycopene and Metabolic Syndrome: A Systematic Review of the Literature. Adv. Nutr. 2018, 10, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Koutsidis, G.; Lodge, J.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and Cardiovasc. risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef]
- Rowles, J.; Ranard, K.; Smith, J.; An, R.; Erdman, J. Increased dietary and circulating lycopene are associated with reduced prostate cancer risk: A systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2017, 20, 361–377. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Michalek, J.; Mesa, R.; Parma, D.; Rodriguez, R.; Mansour, A.; Svatek, R.; Tucker, T.; Ramirez, A. Carotenoid Intake and Circulating Carotenoids Are Inversely Associated with the Risk of Bladder Cancer: A Dose-Response Meta-analysis. Adv. Nutr. 2019, 11, 630–643. [Google Scholar] [CrossRef]
- Nakkeeran, M.; Periasamy, S.; Inmozhi, S.R.; Santha, K.; Sethupathy, S. Increased Levels of Inflammatory Marker hsCRP, MDA and Lipid Profile in Non-obese Hypertension Subjects. Anal. Biochem. 2017, 6, 4. [Google Scholar]
- Zhang, Y.; Zhang, J.; Sheng, H.; Li, H.; Wang, R. Acute phase reactant serum amyloid A in inflammation and other diseases. Adv. Clin.Chem. 2019, 90, 25–80. [Google Scholar] [PubMed]
- Cumpston, M.; Li, T.; Page, M.; Chandler, J.; Welch, V.; Higgins, J.; Thomas, J. Updated guidance for trusted systematic reviews: A new edition of the Cochrane Handbook for Systematic Reviews of Interventions. Cochrane Database Syst. Rev. 2019, 10, ED000142. [Google Scholar] [CrossRef]
- Tacconelli, E. Systematic reviews: CRD’s guidance for undertaking reviews in health care. Lancet Infect. Dis. 2010, 10, 226. [Google Scholar] [CrossRef]
- Mazidi, M.; Kengne, A.; Katsiki, N.; Mikhailidis, D.; Banach, M. Inverse association between serum antioxidant levels and inflammatory markers is moderated by adiposity: A report based on a large representative population sample of American adults. Br. J. Nutr. 2018, 120, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Sanjuán, J.; Calvo-Nieves, M.; Aguirre-Gervás, B.; Herreros-Rodríguez, J.; Velayos-Jiménez, B.; Castro-Alija, M.; Muñoz-Moreno, M.; Sánchez, D.; Zamora-González, N.; Bajo-Grañeras, R.; et al. Early Detection of High Oxidative Activity in Patients with Adenomatous Intestinal Polyps and Colorectal Adenocarcinoma: Myeloperoxidase and Oxidized Low-Density Lipoprotein in Serum as New Markers of Oxidative Stress in Colorectal Cancer. Lab. Med. 2015, 46, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.; Yoe, H.; Kim, H.; Park, J.; Kim, J.; Lee, S.; Lee, J.; Lee, K.; Jang, Y.; Lee, J. Independent inverse relationship between serum lycopene concentration and arterial stiffness. Atherosclerosis 2010, 208, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G.; D’Orazio, N.; Palumbo, N.; Bucciarelli, V.; di Ilio, E.; Bazzano, L.; Bucciarelli, T. Relationship between plasma antioxidant concentrations and carotid intima-media thickness: The Asymptomatic Carotid Atherosclerotic Disease in Manfredonia Study. Eur. J. Cardiol. Prev. Rehabil. 2009, 16, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Hozawa, A.; Jacobs, D.; Steffes, M.; Gross, M.; Steffen, L.; Lee, D. Relationships of Circulating Carotenoid Concentrations with Several Markers of Inflammation, Oxidative Stress, and Endothelial Dysfunction: The Coronary Artery Risk Development in Young Adults (CARDIA)/Young Adult Longitudinal Trends in Antioxidants (YALTA) Study. Clin. Chem. 2007, 53, 447–455. [Google Scholar]
- Walston, J.; Xue, Q.; Semba, R.; Ferrucci, L.; Cappola, A.; Ricks, M.; Guralnik, J.; Fried, L. Serum Antioxidants, Inflammation, and Total Mortality in Older Women. Am. J. Epidemiol. 2005, 163, 18–26. [Google Scholar] [CrossRef]
- Eboumbou, C.; Steghens, J.; Abdallahi, O.; Mirghani, A.; Gallian, P.; van Kappel, A.; Qurashi, A.; Gharib, B.; De Reggi, M. Circulating markers of oxidative stress and liver fibrosis in Sudanese subjects at risk of schistosomiasis and hepatitis. Acta Trop. 2005, 94, 99–106. [Google Scholar] [CrossRef]
- van Herpen-broekmans, W.; Klöpping-ketelaars, I.; Michiel, B.; Cornelis, K.; Hans, P.; Hendriks, F.; Tijburg, L.; van Poppel, G.; Kardinaal, A. Serum carotenoids and vitamins in relation to markers of endothelial. Eur. J. Epidemiol. 2004, 19, 915–921. [Google Scholar] [CrossRef]
- Jonasson, L.; Wikby, A.; Olsson, A. Low serum β-carotene reflects immune activation in patients with coronary artery disease. Nutr. Metab. Cardiovasc. Dis. 2003, 13, 120–125. [Google Scholar] [CrossRef]
- Yadav, D.; Hertan, H.; Schweitzer, P.; Norkus, E.; Pitchumoni, C. Serum and liver micronutrient antioxidants and serum oxidative stress in patients with chronic hepatitis C. Am. J. Gastroenterol. 2002, 97, 2634–2639. [Google Scholar] [CrossRef]
- Kritchevsky, S.; Bush, A.; Pahor, M.; Gross, M. Serum Carotenoids and Markers of Inflammation in Nonsmokers. Am. J. Gastroenterol. 2000, 152, 1065–1071. [Google Scholar]
- McMillan, D.; Sattar, N.; Talwar, D.; O’Reilly, D.; McArdle, C. Changes in micronutrient concentrations following anti-inflammatory treatment in patients with gastrointestinal cancer. Nutrition 2000, 16, 425–428. [Google Scholar]
- Boosalis, M.; Snowdon, D.; Tully, C.; Gross, M. Acute phase response and plasma carotenoid concentrations in older women: Findings from the nun study. Nutrition 1996, 12, 475–478. [Google Scholar]
- Almushatat, A.; Talwar, D.; McArdle, P.; Williamson, C.; Sattar, N.; O’Reilly, D.; Underwood, M.; McMillan, D. Vitamin antioxidants, lipid peroxidation and the systemic inflammatory response in patients with prostate cancer. Int. J. Cancer 2005, 118, 1051–1053. [Google Scholar]
- McMillan, D.; Talwar, D.; Sattar, N.; Underwood, M.; O’Reilly, D.S.J.; McArdle, C. The relationship between reduced vitamin antioxidant concentrations and the systemic inflammatory response in patients with common solid tumours. Clin. Nutr. 2002, 21, 161–164. [Google Scholar] [PubMed]
- Chang, C.; Chen, J.; Ke, D.; Hu, M. Plasma levels of lipophilic antioxidant vitamins in acute ischemic stroke patients: Correlation to inflammation markers and neurological deficits. Nutrition 2005, 21, 987–993. [Google Scholar] [PubMed]
- Chung, R.; Leanderson, P.; Lundberg, A.; Jonasson, L. Lutein exerts anti-inflammatory effects in patients with coronary artery disease. Atherosclerosis 2017, 262, 87–93. [Google Scholar]
- Quasim, T.; McMillan, D.; Talwar, D.; Sattar, N.; O’Reilly, D.; Kinsella, J. Lower concentrations of carotenoids in the critically ill patient are related to a systemic inflammatory response and increased lipid peroxidation. Clin. Nutr. 2003, 22, 459–462. [Google Scholar]
- Nieman, D.; Capps, C.; Capps, C.; Shue, Z.; McBride, J. Effect of 4-Week Ingestion of Tomato-Based Carotenoids on Exercise-Induced Inflammation, Muscle Damage, and Oxidative Stress in Endurance Runners. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 266–273. [Google Scholar]
- Li, Y.; Chang, Y.; Huang, H.; Wu, Y.; Yang, M.; Chao, P. Tomato juice supplementation in young women reduces inflammatory adipokine levels independently of body fat reduction. Nutrition 2015, 31, 691–696. [Google Scholar]
- Biddle, M.; Lennie, T.; Bricker, G.; Kopec, R.; Schwartz, S.; Moser, D. Lycopene Dietary Intervention. J. Cardiovasc. Nurs. 2015, 30, 205–212. [Google Scholar] [PubMed]
- McEneny, J.; Wade, L.; Young, I.; Masson, L.; Duthie, G.; McGinty, A.; McMaster, C.; Thies, F. Lycopene intervention reduces inflammation and improves HDL functionality in moderately overweight middle-aged individuals. J. Nutr. Biochem. 2013, 24, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Petyaev, I.; Dovgalevsky, P.; Klochkov, V.; Chalyk, N.; Pristensky, D.; Chernyshova, M.; Udumyan, R.; Kocharyan, T.; Kyle, N.; Lozbiakova, M.; et al. Effect of lycopene supplementation on Cardiovasc. parameters and markers of inflammation and oxidation in patients with coronary vascular disease. Food Sci. Nutr. 2018, 6, 1770–1777. [Google Scholar] [PubMed]
- Gajendragadkar, P.; Hubsch, A.; Mäki-Petäjä, K.; Serg, M.; Wilkinson, I.; Cheriyan, J. Effects of Oral Lycopene Supplementation on Vascular Function in Patients with Cardiovasc. Disease and Healthy Volunteers: A Randomised Controlled Trial. PLoS ONE 2014, 9, e99070. [Google Scholar]
- Kim, J.; Paik, J.; Kim, O.; Park, H.; Lee, J.; Jang, Y.; Lee, J. Effects of lycopene supplementation on oxidative stress and markers of endothelial function in healthy men. Atherosclerosis 2011, 215, 189–195. [Google Scholar] [CrossRef]
- Markovits, N.; Ben Amotz, A.; Levy, Y. The effect of tomato-derived lycopene on low carotenoids and enhanced systemic inflammation and oxidation in severe obesity. Isr. Med. Assoc. J. 2009, 11, 598–601. [Google Scholar]
- Thies, F.; Masson, L.; Rudd, A.; Vaughan, N.; Tsang, C.; Brittenden, J.; Simpson, W.; Duthie, S.; Horgan, G.; Duthie, G. Effect of a tomato-rich diet on markers of Cardiovasc. disease risk in moderately overweight, disease-free, middle-aged adults: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 95, 1013–1022. [Google Scholar] [CrossRef]
- Upritchard, J.; Sutherland, W.; Mann, J. Effect of supplementation with tomato juice, vitamin E, and vitamin C on LDL oxidation and products of inflammatory activity in type 2 diabetes. Diabetes Care 2000, 23, 733–738. [Google Scholar]
- Jacob, K.; Periago, M.; Böhm, V.; Berruezo, G. Influence of lycopene and vitamin C from tomato juice on biomarkers of oxidative stress and inflammation. Bri. J. Nutr. 2007, 99, 137–146. [Google Scholar]
- Williams, E.; Baines, K.; Smart, J.; Gibson, P.; Wood, L. Rosuvastatin, lycopene and omega-3 fatty acids: A potential treatment for systemic inflammation in COPD; a pilot study. JNIM 2016, 5, 86–95. [Google Scholar]
- Rydén, M.; Leanderson, P.; Kastbom, K.; Jonasson, L. Effects of simvastatin on carotenoid status in plasma. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 66–71. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hurtado-Barroso, S.; Martínez-Huélamo, M.; Rinaldi de Alvarenga, J.; Quifer-Rada, P.; Vallverdú-Queralt, A.; Pérez-Fernández, S.; Lamuela-Raventós, R. Acute Effect of a Single Dose of Tomato Sofrito on Plasmatic Inflammatory Biomarkers in Healthy Men. Nutrients 2019, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- Colmán-Martínez, M.; Martínez-Huélamo, M.; Valderas-Martínez, P.; Arranz-Martínez, S.; Almanza-Aguilera, E.; Corella, D.; Estruch, R.; Lamuela-Raventós, R. Trans-Lycopene from tomato juice attenuates inflammatory biomarkers in human plasma samples: An intervention trial. Mol. Nutr. Food Res. 2017, 61, 1600993. [Google Scholar] [CrossRef]
- Wood, L.; Garg, M.; Smart, J.; Scott, H.; Barker, D.; Gibson, P. Manipulating antioxidant intake in asthma: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 96, 534–543. [Google Scholar] [CrossRef]
- Yeon, J.; Kim, H.; Sung, M. Diets rich in fruits and vegetables suppress blood biomarkers of metabolic stress in overweight women. Prev. Med. 2012, 54, S109–S115. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, A.; Mendez, R.; Zhang, K. Cross Talk Between ER Stress, Oxidative Stress, and Inflammation in Health and Disease. Methods Mol. Biol. 2015, 205–214. [Google Scholar]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Langham, M.; Zhou, Y.; Chirico, E.; Magland, J.; Sehgal, C.; Englund, E.; Mohler, E.; Guo, W.; Barhoum, S.; Wehrli, F. Effects of age and smoking on endothelial function assessed by quantitative Cardiovasc. magnetic resonance in the peripheral and central vasculature. J.Cardiovasc. Magn. Reson. 2015, 17, 1. [Google Scholar] [CrossRef]
- Amirkhizi, F.; Siassi, F.; Minaie, S.; Djalali, M.; Rahimi, A.; Chamari, M. Is obesity associated with increased plasma lipid peroxidation and oxidative stress in women? ARYA Atheroscler. 2010, 2, 189–192. [Google Scholar]
- Yan, Y.; Guo, T.; Zhu, C. Effects of nonsteroidal anti-inflammatory drugs on serum proinflammatory cytokines in the treatment of ankylosing spondylitis. Biochem. Cell Biol. 2018, 96, 450–456. [Google Scholar] [CrossRef]
- Gallelli, L.; Galasso, O.; Falcone, D.; Southworth, S.; Greco, M.; Ventura, V.; Romualdi, P.; Corigliano, A.; Terracciano, R.; Savino, R.; et al. The effects of nonsteroidal anti-inflammatory drugs on Clin.outcomes, synovial fluid cytokine concentration and signal transduction pathways in knee osteoarthritis. A randomized open label trial. Osteoarthr. Cartil. 2013, 21, 1400–1408. [Google Scholar] [PubMed]
- Fielding, J.M.; Rowley, K.G.; Cooper, P.; O’ Dea, K. Increases in plasma lycopene concentration after consumption of tomatoes cooked with olive oil. Asia Pac. J. Clin. Nutr. 2005, 14, 131–136. [Google Scholar] [PubMed]
- Raikos, V. Food matrix: Natural barrier or vehicle for effective delivery of carotenoids from processed foods? Nutr. Metabol. Insights 2017, 1, 1–6. [Google Scholar]
- Crowe, K.M. Designing Functional Foods with Bioactive Polyphenols: Highlighting Lessons Learned from Original Plant Matrices. J. Hum. Nutr. Food Sci. 2014, 1, 1018. [Google Scholar]
- Krinsky, N.; Yeum, K. Carotenoid–radical interactions. Biochem. Biophys. Res. Commun. 2003, 305, 754–760. [Google Scholar]
- Young, A.; Lowe, G. Antioxidant and Prooxidant Properties of Carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar]
- Barros, M.; Rodrigo, M.; Zacarias, L. Dietary Carotenoid Roles in Redox Homeostasis and Human Health. J. Agric. Food Chem. 2018, 66, 5733–5740. [Google Scholar]
- Niess, A. Response and adaptation of skeletal muscle to exercise-the role of reactive oxygen species. Front. Biosci. 2007, 12, 4826. [Google Scholar]
- Halliwell, B. The antioxidant paradox: Less paradoxical now? Br. J. Clin. Pharmacol. 2013, 75, 637–644. [Google Scholar]
- Di Pierro, F. Antioxidants and cancer: A debate on prevention, progression, hormesis, and cruciferous vegetables. Nutrafoods 2015, 14, 175–179. [Google Scholar]
- Tiwari, A. The antioxidant paradox. Pharmacogn. Mag. 2019, 15, 173. [Google Scholar] [CrossRef]
- Biswas, S. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 12, 1–9. [Google Scholar] [CrossRef]
- Bhakkiyalakshmi, E.; Sireesh, D.; Rajaguru, P.; Paulmurugan, R.; Ramkumar, K. The emerging role of redox-sensitive Nrf2–Keap1 pathway in diabetes. Pharmacol. Res. 2015, 91, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, K.; Sthijns, M.; van der Vijgh, W.; Bast, A.; Haenen, G. The antioxidant flavonoid monoHER provides efficient protection and induces the innate Nrf2 mediated adaptation in endothelial cells subjected to oxidative stress. PharmaNutrition 2014, 2, 69–74. [Google Scholar] [CrossRef]
- Bryan, H.; Olayanju, A.; Goldring, C.; Park, B. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Valgimigli, L.; Iori, R. Antioxidant and pro-oxidant capacities of ITCs. Environ. Mol. Mutagen. 2009, 50, 222–237. [Google Scholar] [CrossRef]
- Tanigawa, S.; Fujii, M.; Hou, D. Action of Nrf2 and Keap1 in ARE-mediated NQO1 expression by quercetin. Free Radic. Biol. Med. 2007, 42, 1690–1703. [Google Scholar] [CrossRef]
- Jacobs, H.; Moalin, M.; Bast, A.; van der Vijgh, W.; Haenen, G. An Essential Difference between the Flavonoids MonoHER and Quercetin in Their Interplay with the Endogenous Antioxidant Network. PLoS ONE 2010, 5, e13880. [Google Scholar] [CrossRef]
- Nespolo, M. Free Radicals in Biology and Medicine; Halliwell, B., Gutteridge, J.M.C., Eds.; University Press: New York, NY, USA, 2007. [Google Scholar]
- Kim, Y.; Young, M.; Bobe, G.; Colburn, N.; Milner, J. Bioactive Food Components, Inflammatory Targets, and Cancer Prevention. Cancer Prev. Res. 2009, 2, 200–208. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
| Study (Ref) | Study Population | Final n | Lycopene Measurement | Inflammation Biomarkers | Conclusions |
|---|---|---|---|---|---|
| Mazidi et al. [41] | Participants divided in quartiles depending on CRP and Fibrinogen | Q1: (n = 193) Q2: (n = 190) Q3: (n = 183) Q4: (n = 199) | Serum trans-Lycopene (μmol/L) Q1: 0.431 ± 0.007 Q2: 0.425 ± 0.007 Q3: 0.421 ± 0.005 Q4: 0.387 ± 0.009 | CRP (mg/dL) Q1: 0.03 ± 0.01 Q2: 0.14 ± 0.04 Q3: 0.33 ± 0.07 Q4: 1.2 ± 0.89 | A higher trans-lycopene level for each μmol/L correlated with 0.067 mg/dL lower CRP and 0.048 mg/dL Fibrinogen |
| Crespo-Sanjuán et al. [42] | Control subjects (n = 14) Patients with intestinal polyps (n = 39) Patients with colorectal adenocarcinoma (CRC) (n = 128) | Control (n = 14) Patients (n = 167) | Plasma Lycopene (μg/L) Control: 194.33 ± 66.17 Carc. in Situ: 138.57 ± 106.62 Cancer IV: 100.42 ± 71.20 | Plasma CRP (mg/L) Control: 2.05 ± 2.33 Carc. in Situ: 13.93 ± 26.53 Cancer IV: 41.83 ± 62.01 | Levels of lycopene were higher in the control group and low in the stage-IV group (p = 0.03), and were inversely correlated with CRP (p = 0.005, R = −0.215). We found a consistent relationship between high lycopene and absence of atherosclerosis (p = 0.002). |
| Kim et al. [43] | Healthy women (31–75 yrs) classified into tertiles according to serum lycopene concentration (n = 264) | T1 (n = 88) T2 (n = 88) T3 (n = 88) | Serum Lycopene (mmol/L) T1: 0.029 ± 0.000 T2: 0.039 ± 0.000 T3: 0.052 ± 0.001 | hs-CRP (mg/dL) T1: 1.27 ± 0.24 T2: Data not shown T3: 0.80 ± 0.25 | Subjects in T3 showed lower C-reactive protein (hs-CRP) (0.80 ± 0.25 mg/dL vs. 1.27 ± 0.24 mg/dL, p = 0.015), compared with those in T1. |
| Riccioni et al. [44] | Participants asymptomatic with respect to carotid artery disease divided over 3 groups based on Carotid intima-media thickness (n = 640) | C1 (n = 291) C2 (n = 232) C3 (n = 117) | Plasma Lycopene (μmol/L)C1: 0.82 ± 0.33C2: 0.33 ± 0.63C3: 0.34 ± 0.21 | CRP (g/dL)C1: 2.90 ± 1.30 C2: 3.84 ± 1.75 C3: 4.86 ± 2.20 | Elevated CIMT was significantly associated with having a low concentration of all antioxidants evaluated (vitamin A, vitamin E, lycopene, and b-carotene) and a higher concentration of inflammatory factors including serum uric acid, CRP, and fibrinogen. |
| Hozawa et al. [45] | Men and women in the Coronary Artery Risk Development in Young Adults study (18–30 years) divided in quartiles depending on Lycopene levels (n = 4580) | Q1: (n = 1144) Q2: (n = 1144) Q3: (n = 1144) Q4: (n = 1148) | Serum Lycopene (nmol/L) Q1: 24.2 Q2: 44.1 Q3: 62.0 Q4: 91.8 | CRP (mg/L)Q1: 1.04Q2: 1.11Q3: 0.99Q4: 1.11 | Serum total and individual carotenoids, with the exception of lycopene, were inversely associated with markers of inflammation |
| Walston et al. [46] | Subjects were disabled women aged >65 years (n = 619) | (n = 619) | Serum Lycopene (μmol/L) 0.56 ± 0.31 | IL-6 (pg/mL) 5.51 ± 12.69 | Persons with the highest levels of b-carotene, lycopene, lutein/zeaxanthin, b-cryptoxanthin, and retinol were also significantly less likely to be in the highest interleukin-6 tertile. |
| Eboumbou et al. [47] | Sudanese subjects exposed and not exposed to Schistosoma infection and French control subjects | Rural Sudan: (n = 35) Urban Sudan: (n = 27) French: (n = 34) | Serum Lycopene (μM)/Lycopene:B-carotene ratio RS: 0.21 (0.04)/1.10 US: 0.68 (0.10)/5.11 F: 1.10 (0.25)/4.52 | Hyaluronic acid (HA)/Malondialdehyde (MDA) around 60 μg/L/200 nM | Drastic decrease of lycopene levels in the subjects exposed to schistosomiasis in comparison with non-exposed Sudanese and French control subjects |
| van Herpen-Broekmans et al. [48] | Healthy men and women (n = 379) | Men: (n = 178) Women: (n = 201) Total: (n = 379) | Serum Lycopene (μmol/L) Men: 0.35 ± 0.18 Women: 0.37 ± 0.18 Total: 0.36 ± 0.18 | CRP (mg/L) Men: 0.9 (0.2–5.9) Women: 1.4 (0.2–7) Total: 1.1 (0.2–6.7) | An inverse relation between lycopene and CRP (−1.14 ± 0.54 per umol/l; p = 0.04) was found in men and not in women (0.50 ± 0.50 per umol/l; p = 0.32) |
| Jonasson et al. [49] | Men with stable angina and angiographically verified CAD and healthy controls (n = 113) | Patients: (n = 44) Controls: (n = 69) | Serum Lycopene (nmol/L) Patients: 177 (115–242) Controls: 298 (212–408) | CRP (mg/L) Patients: 2.30 (1.35–4.41) Controls: 1.22 (0.66–2.16) | Compared with controls, patients had signs of an enhanced inflammatory activity assessed by significantly increased levels of CRP. Patients also had significantly lower B-carotene and lycopene levels. |
| Dhiraj et al. [50] | Patients with Chronic Hepatitis C and controls (n = 42) | Patients: (n = 20) Controls: (n = 22) | Serum Lycopene (μg/dL) Patients: 6.2 ± 3 Controls: 59 ± 28 | MDA (μM) Patients: 1.62 ± 0.57 Controls: 0.23 ± 0.15 | Serum MDA levels were significantly higher in CHC patients compared with controls (1.62 ± 0.57 vs. 0.23 ± 0.15 μmol/L) Serum levels of lycopene were significantly decreased in CHC patients. |
| Kritchevsky et al. [51] | Nonsmoking participants aged 25–55 years (n = 4557) divided in tertiles depending on CRP levels | C1 (n = 3180) C2 (n = 924) C3 (n = 453) | Serum Lycopene (μmol/L) C1: 0.46 ± 0.004 C2: 0.45 ± 0.006 C3: 0.41 ± 0.010 | CRP (mg/dL) C1: < 0.21 C2: 0.22–0.88 C3: >0.88–12.8 | Lycopene is significantly lower in higher CRP tertile |
| McMillan et al. [52] | Healthy control subjects and patients with gastrointestinal cancer (n = 24) | Patients: (n = 12) Controls: (n = 12) | Plasma Lycopene (μmol/L) Patients: <0.02 ( <0.02–0.10) Controls: 0.37 (0.15–0.76) | CRP (mg/L) Patients: 91 (5–182) Controls: <5 ( <5–10) | The cancer group had significantly higher C-reactive protein concentrations (p < 0.001) and concentrations of lycopene were significantly lower (p < 0.001) |
| Boosalis et al. [53] | Catholic sisters (nuns) age 77–99 years (n = 85) divided in 2 groups depending on CRP levels | Elevated CRP: (n = 10) Normal CRP: (n = 75) | Plasma Lycopene (μg/dL) Elevated CRP: 9.0 ± 4.0 Normal CRP: 16.6 ± 10.6 | Serum CRP (mg/dL) Elevated CRP: > 1.5 mg/dL Normal CRP: < 1.5 mg/dL | Results showed that the presence of elevated CRP resulted in a significant decrease of lycopene concentrations (p = 0.03) |
| Almushatat et al. [54] | Healthy subjects (C) Patients with benign prostate hyperplasia (B) Localized (L) Metastatic prostate cancer (M) (n = 112) | C: (n = 14) B: (n = 20) L: (n = 40) M: (n = 38) | Plasma Lycopene (μg/L) C: 127 (17–320) B: 128 (18–223) L: 83 (14–687) M: 42 ( <10–226) | MDA (μmol/L) C: 0.73 (0.50–1.40) B: 0.74 (0.35–1.48) L: 0.93 (0.47–2.93) M: 1.01 (0.44–4.67) | Prostate cancer patients had higher concentrations of malondialdehyde (p < 0.05) and lower circulating concentrations of lycopene (p < 0.001). There was a negative correlation between MDA concentrations and lycopene |
| McMillan et al. [55] | Healthy subjects (C) Breast cancer patients (B) Prostate (p)Colorectal (R) (n = 71) | C: (n = 30) B: (n = 15) p: (n = 15) R: (n = 11) | Plasma Lycopene (μg/100 mL) C: 18.0 (6.0–41.0) B: 1.8 ( <1.0–14.6) p: 6.7 (1.5–47.1) R: <1.0 ( <1.0–5.6) | CRP (mg/L) C: 2.0 (0.2–8.5) B: 3.9 (0.29–14.0) p: 8.0 (4.0–123) R: 70 (5.0–182) | Concentrations of CRP were higher and vitamin antioxidants lower in the cancer patients. In normal subjects and cancer patients, CRP concentrations were inversely correlated with circulating concentrations of lycopene. |
| Chang et al. [56] | Healthy controls (H) Ischemic stroke patients, small (S) or large artery (L) (n = 109) | H: (n = 41) S: (n = 35) L: (n = 33) | Plasma Lycopene (μmol/L) H: 0.13 ± 0.09 S: 0.10 ± 0.07 L: 0.09 ± 0.07 | hs-CRP (mg/L) H: 1.6 ± 1.7 S: 6.0 ± 7.0 L: 8.4 ± 15.4 | hs-CRP concentrations are significantly higher in patients with acute ischemic stroke than in healthy controls. Plasma lycopene, was inversely and significantly correlated with CRP. |
| Chung et al. [57] | Patients with stable angina (SA) or acute coronary syndrome (ACS) (n = 193) | SA: (n = 134) ACS: (n = 59) | Plasma Lycopene (μM) SA: 0.41 (0.25–0.65) ACS: 0.37 (0.26–0.58) | IL-6 (pg/mL) SA: 2.21 (1.45–3.03) ACS: 5.01 (2.68–9.36) | Only lutein + zeaxanthin was inversely correlated with IL-6 in SA patients at baseline |
| Quasim et al. [58] | Healthy controls (H) and critically-ill patients (C) (n = 67) | H: (n = 24) C: (n = 43) | Plasma Lycopene (μg/L) H: 189.0 (62.0–465.0) C: 15.5 ( <10.0–137.0) | CRP (mg/L) H: <5 C: 204 (6–345) | Systemic inflammatory response is associated with low carotenoid concentrations |
| Study (Ref) | Study Population | Intervention | Final n | Lycopene Measurement | Inflammation Biomarkers | Conclusions |
|---|---|---|---|---|---|---|
| Nieman et al. [59] | Healthy runners (n = 20) | Lycopene capsule (5 mg/d) or placebo for 4 weeks | (n = 20) | Plasma Lycopene (ng/mL) Pre-supplement: around 500 Post-supplement: around 750 | CRP (mg/L) Pre-supplement: 1.21 ± 1.2 Post-supplement: 1.28 ± 1.0 | Plasma lycopene increased significantly in intervention group compared to placebo (p < 0.001). No alterations in post-exercise measures of oxidative stress and inflammation were found. |
| Li et al. [60] | Healthy young Taiwanese females (n = 25) | 100% pure tomato juice, containing 11.6 mg of lycopene per 100 mL 280 mL/day for 56 days | (n = 25) | Serum Lycopene (μM) Pre-supplement: 0.72 ± 0.36 Post-supplement: 1.94 ± 0.74 | Adiponectin (μg/mL) Pre-supplement: 11.5 ± 5.8 Post-supplement: 14.4 ± 5.2 MCP-1 (pg/mL)Pre-supplement: 126 ± 36 Post-supplement: 97.3 ± 17.9 TBARS (nM) Pre-supplement: 2.35 ± 1.11 Post-supplement: 1.84 ± 0.89 | Tomato juice supplementation resulted in a decrease in levels of the inflammatory adipokine MCP-1, and an increase in levels of the anti-inflammatory adipokine adiponectin. |
| Biddle et al. [61] | Patients NYHA class II or III (n = 40) | V8 juice containing 29.4 mg of lycopene/day for 30 days | Control (n = 18) Intervention (n = 22) | Plasma Lycopene (μmol/L) Control, pre-supl: 0.56 Control, post-supl: 0.58 Intervention, pre-supl: 0.51 Intervention, post-supl: 0.76 | Serum CRP (mg/L) Control, pre-supl: 4.8 ± 3.4 Control, post-supl: 4.5 ± 3.8 Intervention, pre-supl: 3.4 ± 3.1 Intervention, post-supl: 3.1 ± 2.8 | C-reactive protein levels decreased significantly in the intervention group in women and but not in men (p = 0.04). |
| McEneny et al. [62] | Moderately overweight, middle-aged individuals (n = 54) | Control diet ( <10 mg lycopene/week)lycopene-rich diet (224–350 mg/week) lycopene supplement (70 mg/week)for 12 weeks | Control diet (n = 18) Lycopene diet (n = 18) Lycopene supl (n = 18) | Serum Lycopene (mmol/L) Baseline Control: 0.26 (0.03) Lycopene diet: 0.41 (0.04) Lycopene supl: 0.29 (0.03) Week 12 Control: 0.27 (0.03) Lycopene diet: 1.14 (0.05) Lycopene supl: 0.87 (0.06) | Serum Amyloid A (SAA) (μg/L) Baseline Control: 16,269 Lycopene diet: 15,566 Lycopene supl: 16,899 Week 12 Control: 18,882 Lycopene diet: 17,038 Lycopene supl: 12,070 | Lycopene supplement tended to produce a greater response in reducing SAA concentrations and in influencing HDL’s function compared to the high-tomato diet. |
| Petyaev et al. [63] | Patients with coronary vascular disease (n = 142) | 7 mg of lycopene/day for 1 month, two different lycopene supplements | Lactolycopene (L1) (n = 68) Lycosome GA (L2) (n = 74) | Serum Lycopene (ng/mg cholesterol) Baseline L1: 58.0L2: 55.0 Week 4 L1: 87.0 L2: 237.0 | CRP (mg/L)/MDA (μM) Baseline L1: 6.0/141.0 L2: 6.8/154.0 Week 4 L1: 6.2/156.0 L2: 6.1/51.0 | Lycopene supplementation had no impact on serum CRP level. Lactolycopene did not affect inflammatory markers by the end of the interventional period, whereas lycosome-formulated lycopene significantly reduced MDA |
| Gajendragadkar et al. [64] | Statin treated CVD patients and healthy controls (n = 72) | 7 mg lycopene (1) or placebo (2)/day for 2 months Patients (p) and Healthy (H) | P1: (n = 24) P2: (n = 12) H1: (n = 24) H2: (n = 12) | Serum Lycopene (μg/L) Baseline/Day 56 P1: 146/275 P2: 128/178 H1: 170/267 H2: 182/160 | hsCRP (mg/L)/IL-6 (pg/mL)/TNF-a (pg/mL) Baseline P1: 2.13/1.54/2.13 P2: 1.45/1.20/5.55 H1: 1.15/1.32/5.39 H2: 2.83/0.92/5.55 Day 56 P1: 2.37/1.51/2.37 P2: 1.68/0.92/5.65 H1: 1.87/1.02/4.92 H2: 1.65/0.84/5.32 | hsCRP, IL-6 and TNF-a levels were unchanged for lycopene vs. placebo treatment groups in the CVD arm as well as the HV arm |
| Kim et al. [65] | Healthy men (n = 126) | Placebo (p) Low lycopene, 6 mg/d (L) High lycopene, 15 mg/d (H) For 8 weeks | p: (n = 38) L: (n = 41) H: (n = 37) | Serum Lycopene (μg/mL) Baseline/8 weeksp: 0.2/0.2 L: 0.2/0.26 H: 0.2/0.33 | hsCRP (mg/dL)Baseline/8 weeks p: 1.14 ± 0.22/1.10 ± 0.27 L: 1.39 ± 0.33/1.40 ± 0.37 H: 1.25 ± 0.44/0.54 ± 0.10 | A reduction in hs-CRP in the 15-mg lycopene/day group and the inverse correlation between changes in lycopene and changes in hs-CRP in this study, suggest that lycopene may play a role in inflammatory processes by interfering the action of cytokines. |
| Markovits et al. [66] | Obese patients (p) and healthy controls (C) (n = 16) | Patients received Lyc-o-mato, 30 mg/d for 4 weeks | p: (n = 8) C: (n = 8) | Serum Lycopene (μg/mL) C: 0.14 ± 0.07 p;baseline: 0.23 ± 0.22 p;supple: 1.15 ± 0.21 | CRP (mg/L)/IL-6 (pg/mL)/TNF-a (pg/mL) Baseline C: 1.1/1.0/1.4 p: 6.5/3.6/1.4 Week 4 p; placebo: 5.5/3.5/1.4 p; supple: 5.6/4.7/1.5 | CRP and IL-6 levels were significantly higher in obese vs. controls. Following lycopene treatment, a significant elevation of lycopene (1.15 vs. 0.23 μg/mL) (p < 0.001) occurred in the treatment vs. the placebo group. Markers of inflammation were not altered by lycopene. |
| Thies et al. [67] | Moderately overweight, disease-free, middle-aged adults (n = 225) | Control diet (C) High-tomato diet (H) Lycopene capsules (10 mg/d) (L) for 12 weeks | C: (n = 76) H: (n = 81) L: (n = 68) | Plasma Lycopene (μg/mL) Baseline/12 weeks C: 0.4/0.4 H: 0.4/1.1 L: 0.4/0.85 | hsCRP (mg/L) Baseline/12 weeks C: 3.18/2.08 H: 1.51/1.37 L: 2.27/2.16 IL-6 (pg/L) Baseline/12 weeks C: 1.37/1.38 H: 1.21/1.15 L: 1.44/1.31 | None of the inflammatory markers changed significantly after the dietary intervention. These data indicate that a relatively high daily consumption of tomato-based products (equivalent to 32–50 mg lycopene/d) or lycopene supplements (10 mg/d) is ineffective at reducing conventional CVD risk markers in moderately overweight, healthy, middle-aged individuals. |
| Upritchard et al. [68] | Patients with well-controlled type 2 diabetes aged <75 years (n = 57) | Placebo (C) Tomato juice 500 mL/d (T) for 4 weeks | C: (n = 13) T: (n = 15) | Plasma Lycopene (μmol/L) Baseline/4 weeks C: 0.31/0.28 T: 0.39/1.08 | Plasma CRP (mg/L) Baseline/4 weeks C: 3.1/3.1 T: 3.8/4.1 | Plasma lycopene levels increased nearly three-fold (p = 0.001) and no significant decreases in plasma levels of CRP |
| Jacob et al. [69] | Healthy subjects (n = 24) | 2 weeks depletion followed by 2 weeks tomato juice 500 mL/d (41 mg/L lycopene, 90 mg/L Vitamin C) (L) or enriched with Vitamin C (870 mg/L) (LC) | T-2: baseline T0: after depl. T + 2: after inter. L: (n = 12) LC: (n = 12) | Plasma Lycopene (μmol/L) L/LC T-2: 0.72/0.71 T0: 0.42/0.34 T + 2: 1.05/0.91 | L/LC CRP (ug/L) T-2: 336.2/349.5 T0: 315.6/319.2 T + 2: 262.3/247.1 IL-1 B (ng/L) T-2: 3.45/12.59 T0: 3.87/10.68 T + 2: 4.39/6.40 TNF-a (ng/L) T-2: 6.97/2.93 T0: 6.01/3.35 T + 2: 3.45/3.28 MDA (μmol/L)T-2: 0.55/0.60 T0: 0.54/0.56 T + 2: 0.53/0.50 | The consumption of tomato juice led to a reduction of CRP in both groups. All other markers were affected to a lesser extent or remained unchanged. |
| Williams et al. [70] | COPD patients (n = 11) | Rosuvastatin (20 mg/day) for 4 weeks then a combination of rosuvastatin (20 mg/day), DHA and EPA (1.5 g/day) and lycopene (45 mg/day) for 8 weeks. | T1: baseline T2: rosuvastatin T3: lycopene | Plasma Lycopene (mg/L) T1: 0.30 (0.13–0.54) T2: 0.56 (0.14–0.77) T3: 0.50 (0.22–0.96) | CRP (mg/L) T1: 3.9 (1.9–7.9) T2: 3.3 (0.7–7.6) T3: 3.8 (1.3–8.9) IL-6 (pg/mL) T1: 2.2 (1.6–3.0) T2: 3.2 (2.3–5.1) T3: 3.1 (1.6–4.8) | Treatment interventions did not significantly change plasma carotenoid levels. However, there was a trend for increased lycopene concentration at visit 2 and 3. Following the interventions, plasma IL-6 and CRP were unchanged. |
| Rydén et al. [71] | Middle-aged men with mild to moderate hypercholesterolemia (n = 76) | Placebo (p) Simvastatin 40 mg (S)for 6 weeks | p: (n = 39) S: (n = 37) | Plasma Lycopene (nmol/L/cholesterol) Baseline p: 116 (89–149) S: 100 (75–142) Week 6 p: 125 (98–160) S: 147 (104–182) | CRP (mg/L)/IL-6 (pg/mL) Baseline p: 1.1/1.2 S: 1.3/1.5 Week 6 p: 1.0/1.3 S: 0.9/1.4 | Simvastatin use was associated with significant reductions in CRP and reduced plasma levels of lycopene. However, when adjusted for lipids, lycopene showed significant increases after simvastatin therapy. |
| Hurtado-Barroso et al. [72] | Healthy male subjects (n = 22) | Single dose of sofrito (240 g/70 kg) | T1: baseline T2: intervention | Plasma Lycopene (μmol/L) Baseline/After consumption trans-lycopene: 2.15 ± 0.30/6.33 ± 1.53 5-cis-lycopene: 1.87 ± 0.28/7.93 ± 2.73 13-cis-lycopene: 0.21 ± 0.11/2.08 ± 0.78 9-cis-lycopene: n.d./0.90 ± 0.58 | CRP (mg/dL) T1: 0.1 T2: 0.08 IL-6 (pg/mL) T1: 1.4 T2: 1.0TNF-a (pg/mL) T1: 1.0 T2: 0.8 | After the sofrito intake, a significant decrease in CRP (p = 0.010) and TNF-α (p = 0.011) was observed. |
| Colmán-Martínez et al. [73] | Subject at high Cardiovasc. risk (n = 28) | Tomato Juice HD 400 mL/d LD 200 mL/d Control: Water for 4 weeks | C: (n = 28) LD: (n = 28) HD: (n = 28) | Plasma Lycopene (μmol/L) trans-lycopeneC: 0.70 ± 0.44 LD: 4.04 ± 0.39 HD: 6.67 ± 0.38 5-cis-lycopeneC: 1.13 ± 0.28 LD: 2.38 ± 0.27 HD: 4.08 ± 0.26 13-cis-lycopeneC: 1.07 ± 0.39 LD: 1.90 ± 0.30 HD: 4.01 ± 0.29 9-cis-lycopeneC: 0.42 ± 0.43 LD: 1.05 ± 0.29 HD: 1.92 ± 0.21 | CRP (ng/mL) C: 546 ± 46 LD: 442 ± 44 HD: 530 ± 43 IL-8 (pg/mL) C: 40 ± 17 LD: 23 ± 16 HD: 24 ± 15 | Plasma lycopene increased significantly in intervention group compared to placebo (p < 0.001). No significant alterations in CRP and IL-8 were found. |
| Wood et al. [74] | Asthmatic adults (n = 137) | High-antioxidant diet (HAO) or a low-antioxidant diet (LAO) for 14 d Subjects who consumed the low-antioxidant diet received placebo or tomato extract (45 mg lycopene/d). | HAO: (n = 46) LAO: (n = 91) | Plasma Lycopene (mg/L) Baseline/day 14 HAO: 0.15/0.18 LAO: 0.20/0.13 | hsCRP (mg/L)/IL-6 (pg/mL)/TNF-a (pg/L) HAO baseline: 4.2/1.9/1.3 HAO day 14: 3.0/1.9/1.3 LAO baseline: 2.5/1.9/1.4 LAO day 14: 3.3/2.0/1.5 | After 14 d of dietary modification, a significant decrease from baseline in plasma lycopene concentrations was observed in the LAO diet group, which was significantly different from the increase in the HAO. No effect of the lycopene-rich supplement compared with placebo was observed. Subjects in the low-antioxidant diet group had increased plasma C-reactive protein at week 14. |
| Yeon et al. [75] | Overweight women (n = 22) | High-Vegetable/Fruit (VF) diet (12 servings of VF/day) or low-VF diet (2 servings of VF/day) for 2 weeks, 2 weeks wash-out, 2 weeks | Low base (LB): (n = 22) Low post (LP): (n = 22) High base (HB): (n = 22) High post (HP): (n = 22) | Plasma Lycopene (μmol/L) LB: 0.39 ± 0.18 LP: 0.32 ± 0.14 HB: 0.31 ± 0.19 HP: 0.38 ± 0.32 | CRP (μg/mL)/IL-6 (pg/mL) LB: 0.54 ± 0.44/3.65 ± 1.51 LP: 0.75 ± 0.70/3.08 ± 0.35 HB: 0.56 ± 0.61/3.52 ± 1.08 HP: 0.40 ± 0.40/3.44 ± 0.83 | Results from this study showed that the low-VF diet decreased the average plasma carotenoids by 26%, and the high-VF diet increased the average plasma carotenoids by 32% compared to the baseline values. Changes in plasma lycopene were inversely correlated with changes in plasma IL-6 concentrations when the subjects consumed the low-VF diet. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Steenwijk, H.P.; Bast, A.; de Boer, A. The Role of Circulating Lycopene in Low-Grade Chronic Inflammation: A Systematic Review of the Literature. Molecules 2020, 25, 4378. https://doi.org/10.3390/molecules25194378
van Steenwijk HP, Bast A, de Boer A. The Role of Circulating Lycopene in Low-Grade Chronic Inflammation: A Systematic Review of the Literature. Molecules. 2020; 25(19):4378. https://doi.org/10.3390/molecules25194378
Chicago/Turabian Stylevan Steenwijk, Hidde P., Aalt Bast, and Alie de Boer. 2020. "The Role of Circulating Lycopene in Low-Grade Chronic Inflammation: A Systematic Review of the Literature" Molecules 25, no. 19: 4378. https://doi.org/10.3390/molecules25194378
APA Stylevan Steenwijk, H. P., Bast, A., & de Boer, A. (2020). The Role of Circulating Lycopene in Low-Grade Chronic Inflammation: A Systematic Review of the Literature. Molecules, 25(19), 4378. https://doi.org/10.3390/molecules25194378