
Extract of Boehmeria nivea Suppresses Mast Cell-Mediated Allergic Inflammation by Inhibiting Mitogen-Activated Protein Kinase and Nuclear Factor-κB

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
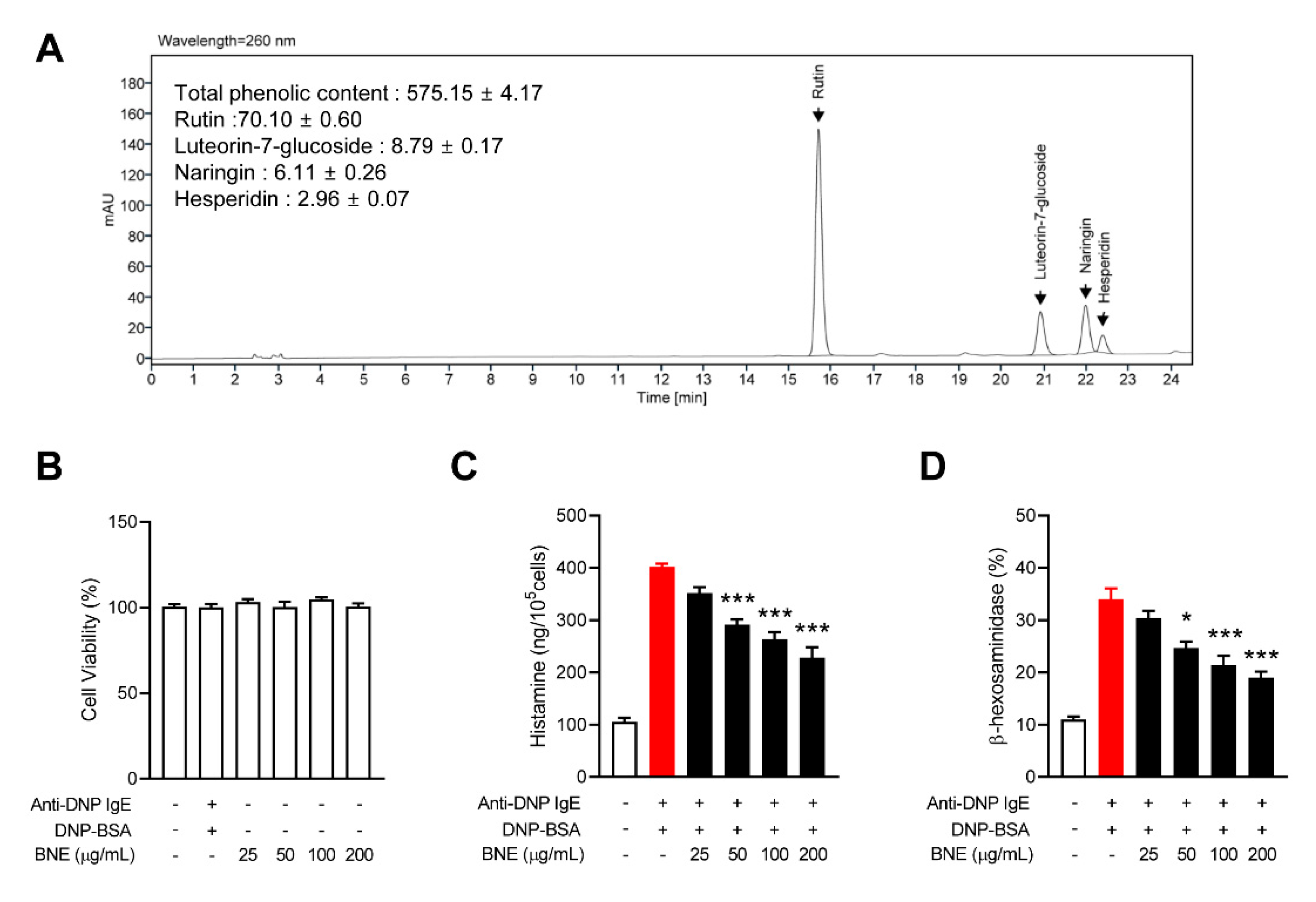
2.1. BNE Inhibits Mast Cell Degranulation
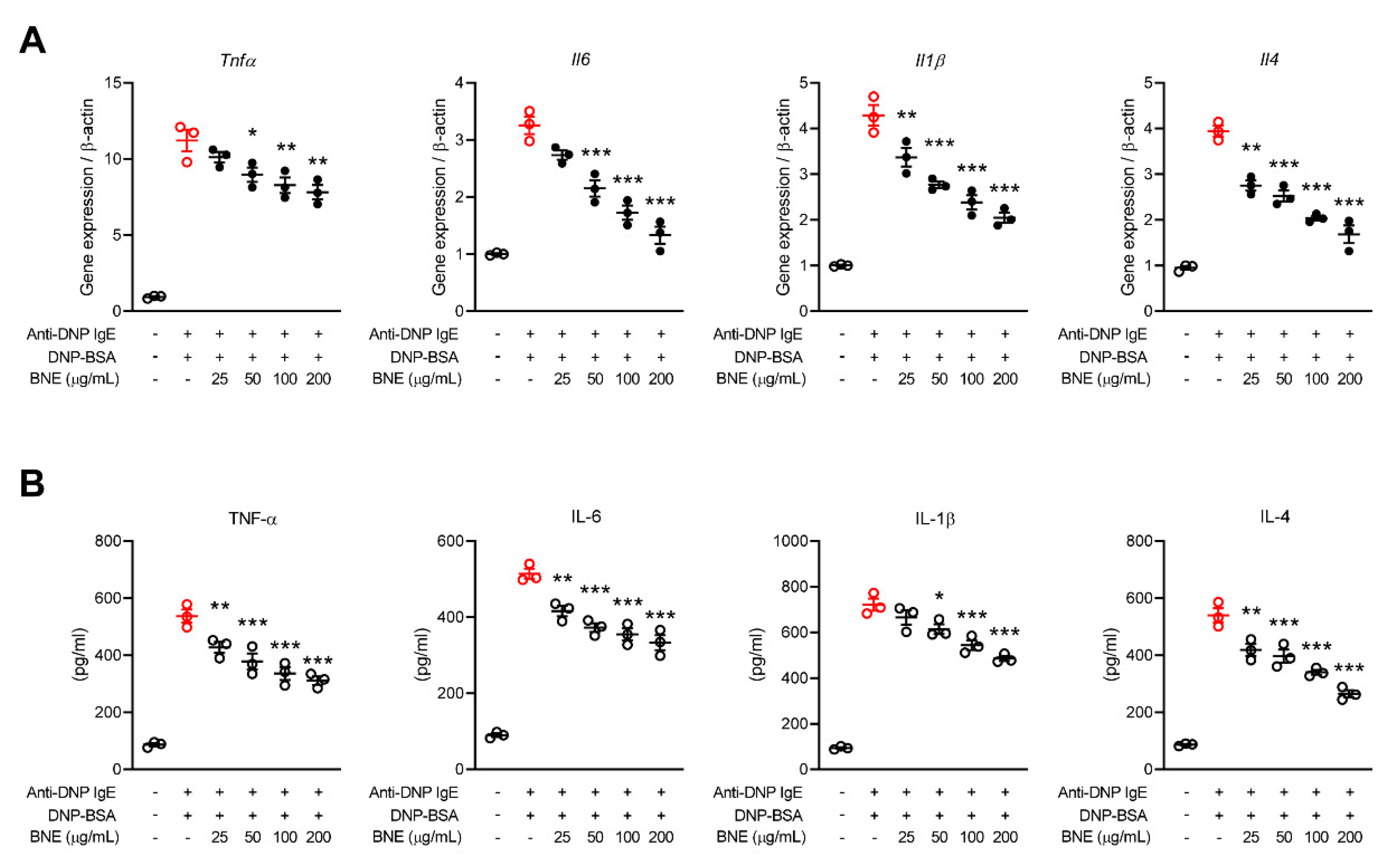
2.2. BNE Reduces Mast Cell-Mediated Inflammatory Cytokine Production
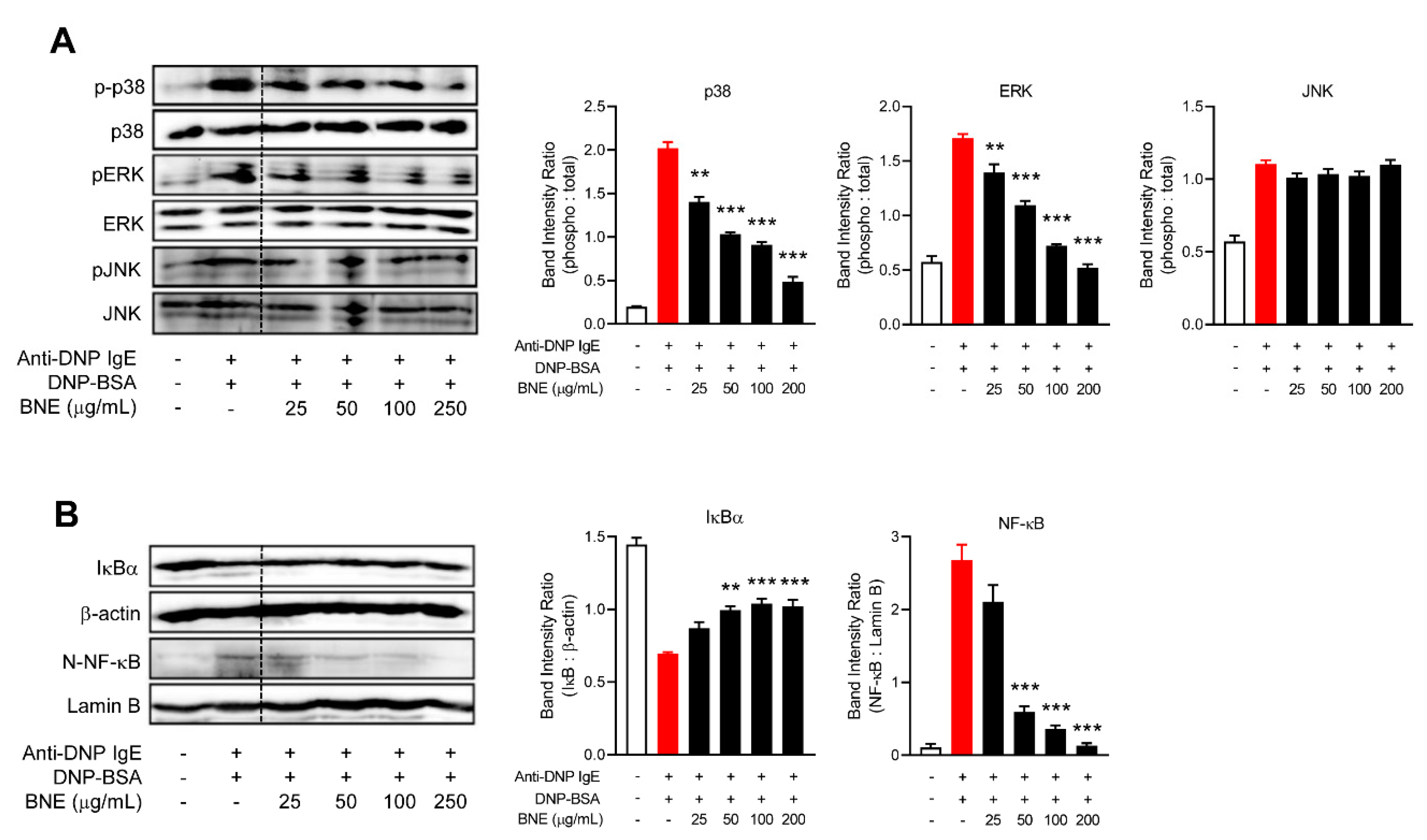
2.3. BNE Suppresses Inflammatory Cytokines by Downregulating p38, ERK MAPK, and Nuclear Factor-κB (NF-κB) Pathways
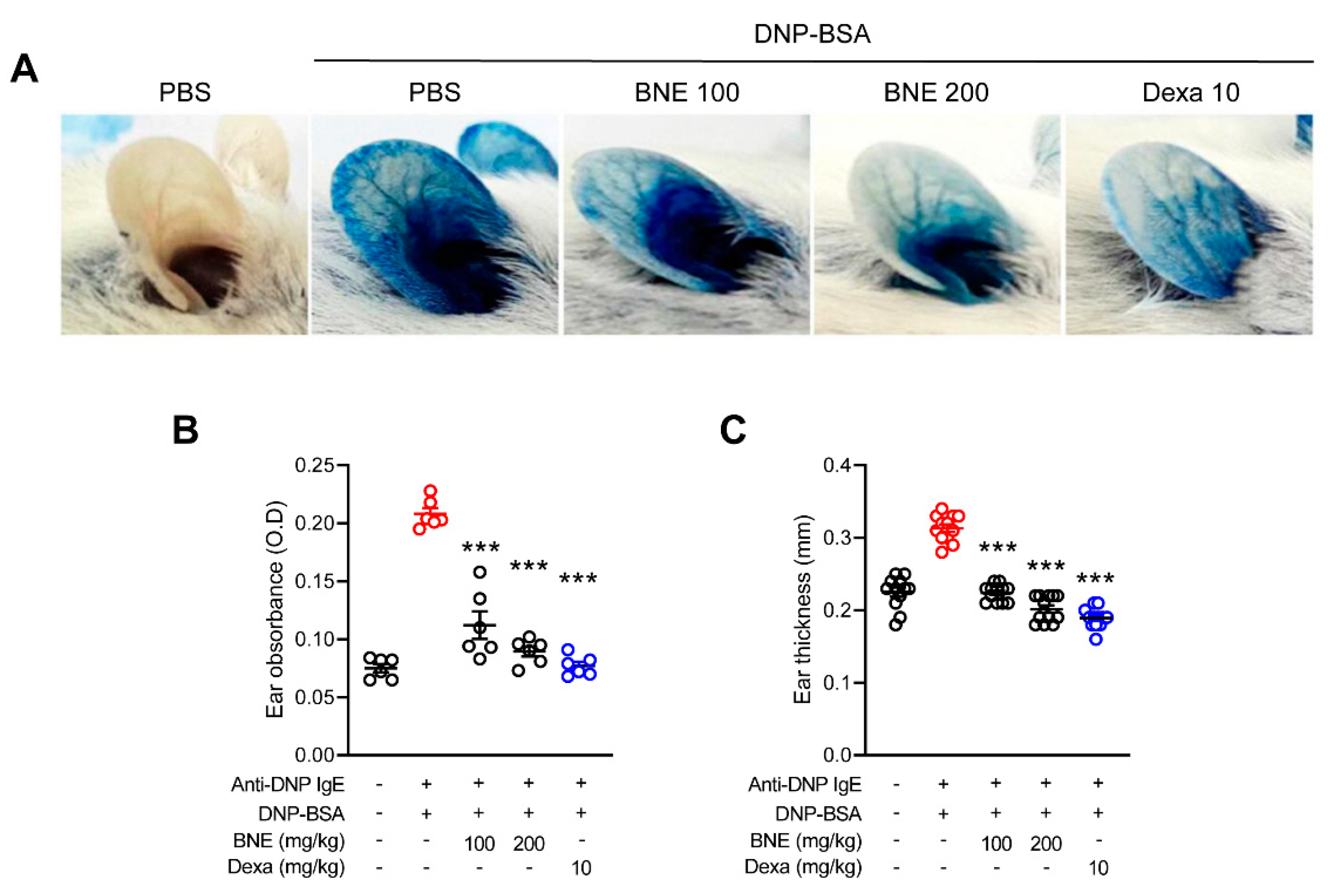
2.4. BNE Attenuates IgE-Mediated Local Cutaneous Anaphylaxis Reaction
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparing BNE and HPLC Analysis
4.3. Animals
4.4. IgE-Mediated Passive Cutaneous Anaphylaxis
4.5. MTT Assay
4.6. β-Hexosaminidase and Histamine Release
4.7. Inflammatory Cytokine Assay
4.8. Real-Time Polymerase Chain Reaction
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TNF | Tumor necrosis factor |
| IL | Interleukin |
| MAPK | Mitogen-activated protein kinase |
| NF-κB | Nuclear factor kappa B |
| BSA | Bovine serum albumin |
| PCA | Passive cutaneous anaphylaxis |
References
- Bischoff, S.C. Role of mast cells in allergic and non-allergic immune responses: Comparison of human and murine data. Nat. Rev. Immunol. 2007, 7, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Akin, C. Mast cell activation disorders. J. Allergy Clin. Immunol. Pract. 2014, 2, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, D.D.; Peavy, R.D.; Gilfillan, A.M. Mechanisms of mast cell signaling in anaphylaxis. J. Allergy Clin. Immunol. 2009, 124, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Lin, L.; Su, X.; Qin, S.; Xu, Q.; Tang, Z.; Deng, Y.; Zhou, Y.; He, S. Anti-hepatitis B virus activity of Boehmeria nivea leaf extracts in human HepG2.2.15 cells. Biomed. Rep. 2014, 2, 147–151. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sung, M.J.; Davaatseren, M.; Kim, S.H.; Kim, M.J.; Hwang, J.T. Boehmeria nivea attenuates LPS-induced inflammatory markers by inhibiting p38 and JNK phosphorylations in RAW264.7 macrophages. Pharm. Biol. 2013, 51, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Sung, M.J.; Yang, H.J.; Kim, M.S.; Hwang, J.T. Boehmeria nivea attenuates the development of dextran sulfate sodium-induced experimental colitis. Mediat. Inflamm. 2014, 2014, 231942. [Google Scholar] [CrossRef] [PubMed]
- Semwal, D.K.; Rawat, U.; Semwal, R.; Singh, R.; Krishan, P.; Singh, M.; Singh, G.J. Chemical constituents from the leaves of Boehmeria rugulosa with antidiabetic and antimicrobial activities. J. Asian Nat. Prod. Res. 2009, 11, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.F.; Jin, X.; Lee, D.; Yang, Y.T.; Lee, K.; Hong, Y.S.; Lee, J.H.; Lee, J.J. Phenanthroquinolizidine alkaloids from the roots of Boehmeria pannosa potently inhibit hypoxia-inducible factor-1 in AGS human gastric cancer cells. J. Nat. Prod. 2006, 69, 1095–1097. [Google Scholar] [CrossRef] [PubMed]
- Sancheti, S.; Sancheti, S.; Bafna, M.; Kim, H.R.; You, Y.H.; Seo, S.Y. Evaluation of antidiabetic, antihyperlipidemic and antioxidant effects of Boehmeria nivea root extract in streptozotocin-induced diabetic rats. Rev. Bras. Farmacogn. 2011, 21, 146–154. [Google Scholar] [CrossRef]
- Lee, T.T.; Chen, C.L.; Shieh, Z.H.; Lin, J.C.; Yu, B. Study on antioxidant activity of Echinacea purpurea L. extracts and its impact on cell viability. Afr. J. Biotechnol. 2009, 8, 5097–5105. [Google Scholar]
- Chirumbolo, S. Dietary assumption of plant polyphenols and prevention of allergy. Curr. Pharm. Des. 2014, 20, 811–839. [Google Scholar] [CrossRef] [PubMed]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The role of histamine and histamine receptors in mast cell-mediated allergy and inflammation: The hunt for new therapeutic targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Alysandratos, K.D.; Angelidou, A.; Delivanis, D.A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast cells and inflammation. Biochim. Biophys. Acta 2012, 1822, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, G.; Wang, H.; Cheng, C.; Zang, G.; Guo, X.; Liu, R.H. Phytochemical profiles and antioxidant activities in six species of ramie leaves. PLoS ONE 2014, 9, e108140. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qiu, C.; Chen, L.; Abbasi, A.M.; Guo, X.; Liu, R.H. Comparative study of phenolic profiles, antioxidant and antiproliferative activities in different vegetative parts of ramie (Boehmeria nivea L.). Molecules 2019, 24, 1551. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Lee, J.Y.; Cho, C.H.; Kim, C.J. Anti-asthmatic action of quercetin and rutin in conscious guinea-pigs challenged with aerosolized ovalbumin. Arch. Pharm. Res. 2007, 30, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.K.; Kim, S.H. Rutin suppresses atopic dermatitis and allergic contact dermatitis. Exp. Biol. Med. (Maywood) 2013, 238, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Kilic, K.; Sakat, M.S.; Yildirim, S.; Kandemir, F.M.; Gozeler, M.S.; Dortbudak, M.B.; Kucukler, S. The amendatory effect of hesperidin and thymol in allergic rhinitis: An ovalbumin-induced rat model. Eur. Arch. Otorhinolaryngol. 2019, 276, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, A.R.; Lee, J.J. Ramie leaf extracts suppresses adipogenic differentiation in 3T3-L1 cells and pig preadipocytes. Asian Australas. J. Anim. Sci. 2016, 29, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Sung, M.J.; Park, J.H.; Yang, H.J.; Hwang, J.T. Boehmeria nivea stimulates glucose uptake by activating peroxisome proliferator-activated receptor gamma in C2C12 cells and improves glucose intolerance in mice fed a high-fat diet. Evid. Based. Complement. Alternat. Med. 2013. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S.; Baker, D.L.; Baker, A. Cellular and Molecular Immunology, 9th ed.; Elsevier: Philadelphia, PA, USA, 2018; p. 565. [Google Scholar]
- Choi, Y.A.; Lee, S.; Choi, J.K.; Kang, B.C.; Kim, M.J.; Dhakal, H.; Kwon, T.K.; Khang, D.; Kim, S.H. The suppressive effect of dabrafenib, a therapeutic agent for metastatic melanoma, in IgE-mediated allergic inflammation. Int. Immunopharmacol. 2020, 83, 106398. [Google Scholar] [CrossRef] [PubMed]
- Roitt, I.M.; Brostoff, J.; Male, D.K. Immunology, 9th ed.; Elsevier: Philadelphia, PA, USA, 2020; p. 416. [Google Scholar]
- Azzolina, A.; Guarneri, P.; Lampiasi, N. Involvement of p38 and JNK MAPKs pathways in substance P-induced production of TNF-alpha by peritoneal mast cells. Cytokine 2002, 18, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Guma, M.; Kashiwakura, J.; Crain, B.; Kawakami, Y.; Beutler, B.; Firestein, G.S.; Kawakami, T.; Karin, M.; Corr, M. JNK1 controls mast cell degranulation and IL-1{beta} production in inflammatory arthritis. Proc. Natl. Acad. Sci. USA 2010, 107, 22122–22127. [Google Scholar] [CrossRef] [PubMed]
- Rao, A. NF-ATp: A transcription factor required for the co-ordinate induction of several cytokine genes. Immunol. Today 1994, 15, 274–281. [Google Scholar] [CrossRef]
- Lin, A.; Minden, A.; Martinetto, H.; Claret, F.X.; Lange-Carter, C.; Mercurio, F.; Johnson, G.L.; Karin, M. Identification of a dual specificity kinase that activates the Jun kinases and p38-Mpk2. Science 1995, 268, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and allergy: From basic mechanisms to clinical applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef] [PubMed]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bulow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. {Alaskhar Alhamwe, 2018 #201}and their role in epigenetics of atopy and allergic diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [PubMed]
- Marwick, J.A.; Tudor, C.; Khorasani, N.; Michaeloudes, C.; Bhavsar, P.K.; Chung, K.F. Oxidants induce a corticosteroid-insensitive phosphorylation of histone 3 at serine 10 in monocytes. PLoS ONE 2015, 10, e0124961. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.K.; Jang, Y.H.; Lee, S.; Lee, S.R.; Choi, Y.A.; Jin, M.; Choi, J.H.; Park, J.H.; Park, P.H.; Choi, H.; et al. Chrysin attenuates atopic dermatitis by suppressing inflammation of keratinocytes. Food Chem. Toxicol. 2017, 110, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, Y.Y.; Choi, Y.A.; Baek, M.C.; Lee, B.; Park, P.H.; Shin, T.Y.; Kwon, T.K.; Khang, D.; Kim, S.H. Elaeocarpusin inhibits mast cell-mediated allergic inflammation. Front. Pharmacol. 2018, 9, 591. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.Y.; Lee, S.; Jang, H.J.; Hur, G.; Lee, S.W.; Jung, K.; Lee, S.J.; Kim, S.H.; Rho, M.C. Cynanchum atratum ameliorates airway inflammation via maintaining alveolar barrier and regulating mast cell-mediated inflammatory responses. Am. J. Chin. Med. 2019, 47, 1795–1814. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, D.L.; Walker, L.L. Dependence of mast cell IgE-mediated cytokine production on nuclear factor-kappaB activity. J. Allergy Clin. Immunol. 2000, 105, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Kempuraj, D.; Di Gioacchino, M.; Boucher, W.; Letourneau, R.; Kandere, K.; Barbacane, R.C.; Reale, M.; Felaco, M.; Frydas, S.; et al. Interleukin-6 and mast cells. Allergy Asthma Proc. 2002, 23, 331–335. [Google Scholar] [PubMed]
- Nam, S.T.; Kim, H.W.; Kim, H.S.; Park, Y.H.; Lee, D.; Lee, M.B.; Min, K.Y.; Kim, Y.M.; Choi, W.S. Furaltadone suppresses IgE-mediated allergic response through the inhibition of Lyn/Syk pathway in mast cells. Eur. J. Pharmacol. 2018, 828, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.; Ha, J.H.; Lee, M.G.; Kim, J.E.; et al. {Park, 2008 #136}and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [PubMed]
- Park, S.H.; Park, E.K.; Kim, D.H. Citrus unshiu and Poncirus trifoliata. Planta Med. 2005, 71, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Takano, S.; Masuda, M.; Iinuma, M.; Matsuda, H. Anti-degranulating activity in rat basophil leukemia RBL-2H3 cells of flavanone glycosides and their aglycones in citrus fruits. J. Nat. Med. 2013, 67, 643–646. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.-Y.; Lee, J.-H.; Lee, B.-R.; Kim, M.A.; Lee, Y.-M.; Kim, D.-K.; Choi, J.K. Extract of Boehmeria nivea Suppresses Mast Cell-Mediated Allergic Inflammation by Inhibiting Mitogen-Activated Protein Kinase and Nuclear Factor-κB. Molecules 2020, 25, 4178. https://doi.org/10.3390/molecules25184178
Lim J-Y, Lee J-H, Lee B-R, Kim MA, Lee Y-M, Kim D-K, Choi JK. Extract of Boehmeria nivea Suppresses Mast Cell-Mediated Allergic Inflammation by Inhibiting Mitogen-Activated Protein Kinase and Nuclear Factor-κB. Molecules. 2020; 25(18):4178. https://doi.org/10.3390/molecules25184178
Chicago/Turabian StyleLim, Ji-Ye, Ji-Hyun Lee, Bo-Ri Lee, Mi Ae Kim, Young-Mi Lee, Dae-Ki Kim, and Jin Kyeong Choi. 2020. "Extract of Boehmeria nivea Suppresses Mast Cell-Mediated Allergic Inflammation by Inhibiting Mitogen-Activated Protein Kinase and Nuclear Factor-κB" Molecules 25, no. 18: 4178. https://doi.org/10.3390/molecules25184178
APA StyleLim, J.-Y., Lee, J.-H., Lee, B.-R., Kim, M. A., Lee, Y.-M., Kim, D.-K., & Choi, J. K. (2020). Extract of Boehmeria nivea Suppresses Mast Cell-Mediated Allergic Inflammation by Inhibiting Mitogen-Activated Protein Kinase and Nuclear Factor-κB. Molecules, 25(18), 4178. https://doi.org/10.3390/molecules25184178