
Click Decoration of Bombyx mori Silk Fibroin for Cell Adhesion Control



Abstract
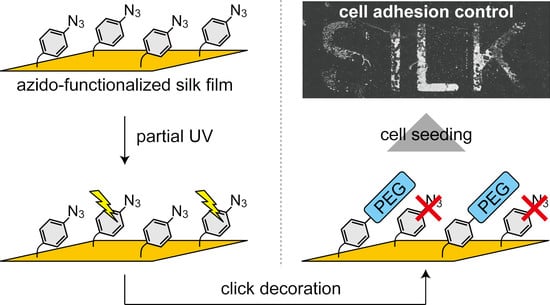
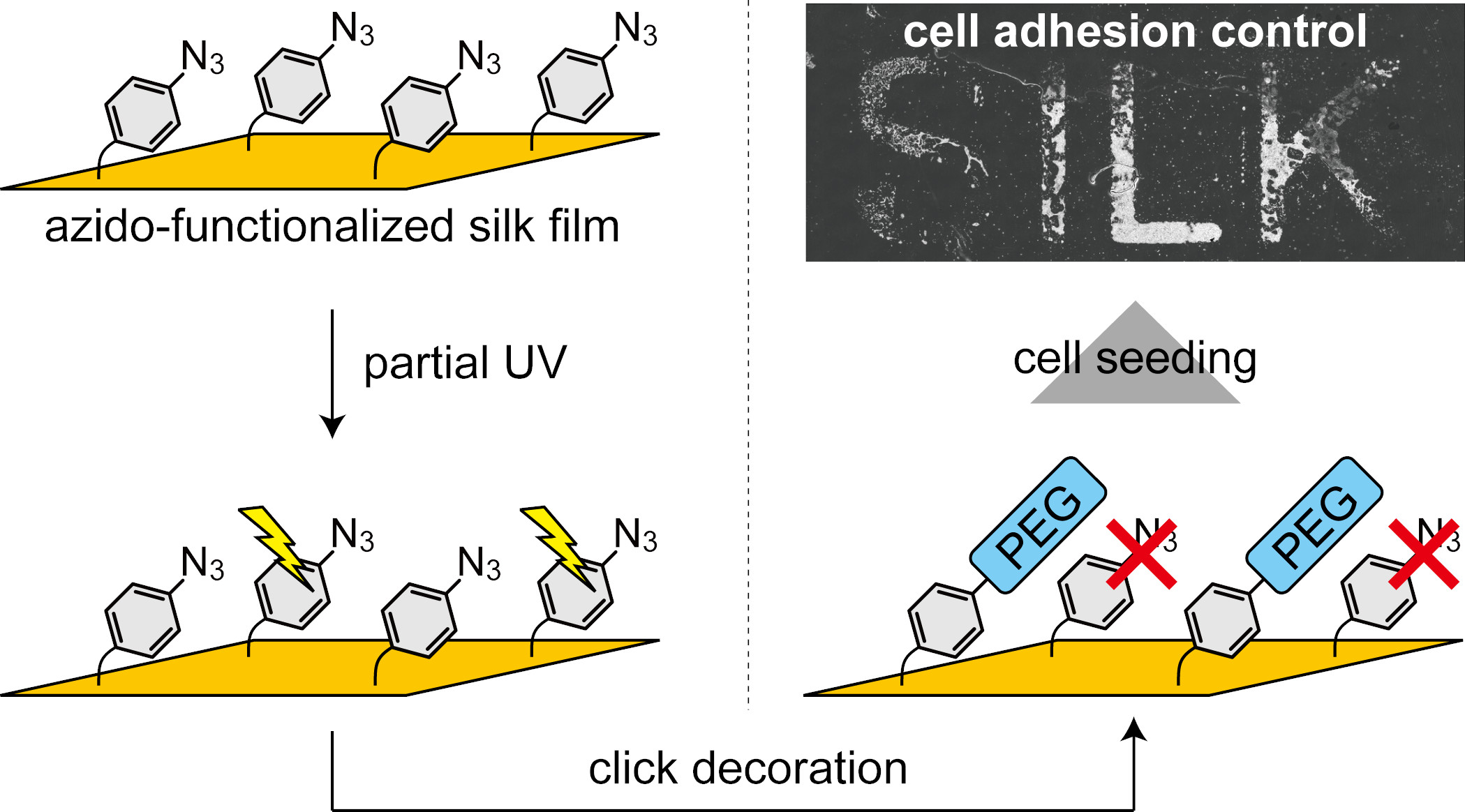{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
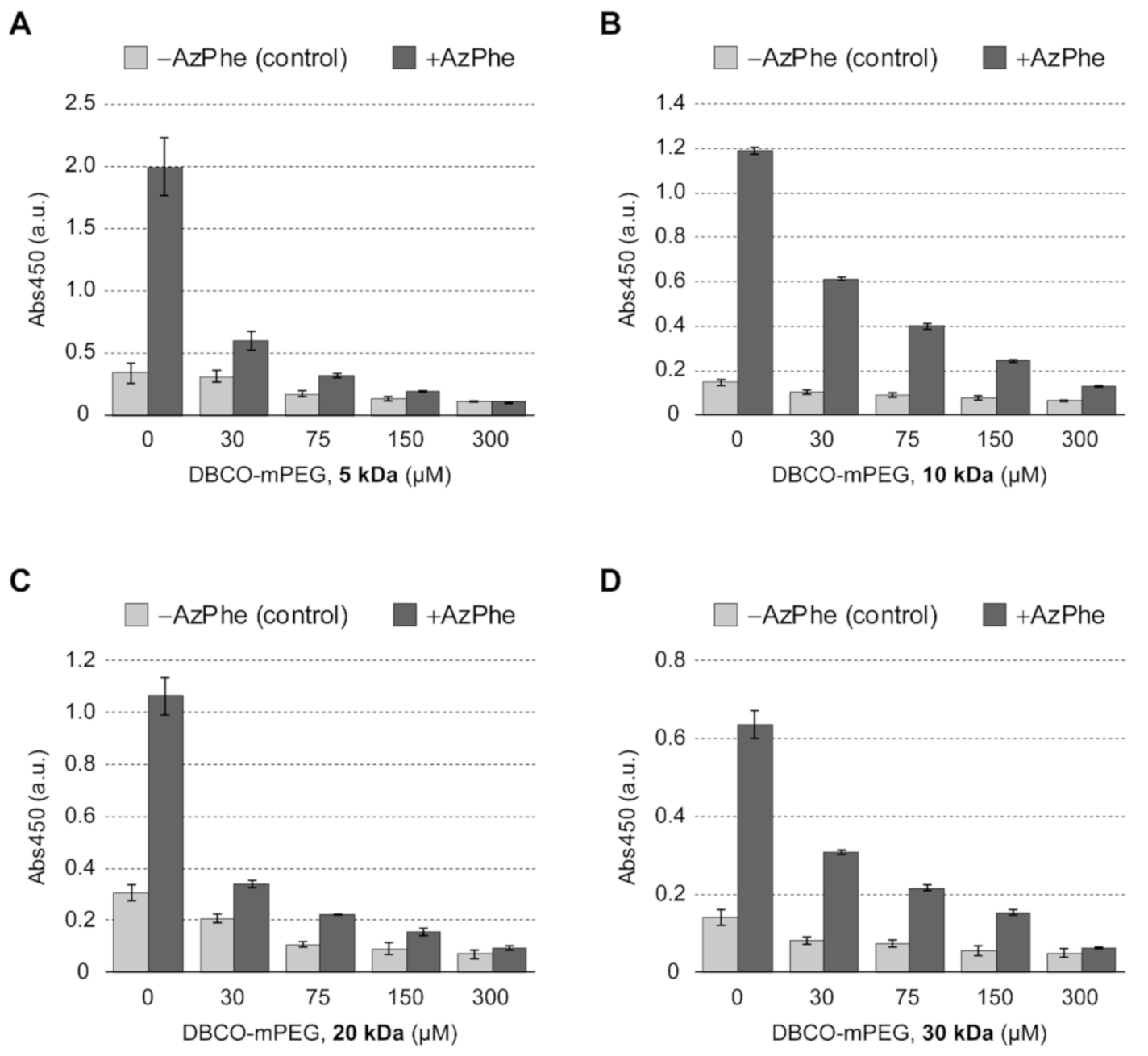
2.1. Click Decoration of Silk Fibroin with PEG
2.2. Cell Adhesion on PEG-Decorated Silk Fibroin Film
2.3. Spatial Patterning of Cells on Partially PEG-Decorated Silk Fibroin Film
3. Materials and Methods
3.1. Materials and Animals
3.2. Production of AzidoSilk
3.3. Preparation of Fibroin Aqueous Solution
3.4. Preparation of Fibroin Film-Coated Cell Culture Plates and Dishes
3.5. Click Decoration Tests
3.6. Cell Adhesion Tests
3.7. Cell Patterning Tests
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhou, C.Z.; Confalonieri, F.; Medina, N.; Zivanovic, Y.; Esnault, C.; Yang, T.; Jacquet, M.; Janin, J.; Duguet, M.; Perasso, R.; et al. Fine organization of Bombyx mori fibroin heavy chain gene. Nucl. Acids Res. 2000, 28, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.; Numata, K.; Rnjak-Kovacina, J.; Seib, F.P. The biomedical use of silk: Past, present, future. Adv. Healthc. Mater. 2019, 8, 1800465. [Google Scholar] [CrossRef] [PubMed]
- Rockwood, D.N.; Preda, R.C.; Yücel, T.; Wang, X.; Lovett, M.L.; Kaplan, D.L. Materials fabrication from Bombyx mori silk fibroin. Nat. Protoc. 2011, 6, 1612–1631. [Google Scholar] [CrossRef] [PubMed]
- Thurber, A.E.; Omenetto, F.G.; Kaplan, D.L. In vivo bioresponses to silk proteins. Biomaterials 2015, 71, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.D.; Kimmerling, E.P.; Cairns, D.M.; Kaplan, D.L. Silk as a biomaterial to support long-term three-dimensional tissue cultures. ACS Appl. Mater. Interfaces 2016, 8, 21861–21868. [Google Scholar] [CrossRef]
- Leal-Egaña, A.; Scheibel, T. Silk-based materials for biomedical applications. Biotechnol. Appl. Biochem. 2010, 55, 155–167. [Google Scholar] [CrossRef]
- Murphy, A.R.; Kaplan, D.L. Biomedical applications of chemically-modified silk fibroin. J. Mater. Chem. 2009, 19, 6443–6450. [Google Scholar] [CrossRef]
- Volkov, V.; Ferreira, A.V.; Cavaco-Paulo, A. On the routines of wild-type silk fibroin processing toward silk-inspired materials: A review. Macromol. Mater. Eng. 2015, 300, 1199–1216. [Google Scholar] [CrossRef]
- Mukai, T.; Lajoie, M.J.; Englert, M.; Söll, D. Rewriting the genetic code. Annu. Rev. Microbiol. 2017, 71, 557–577. [Google Scholar] [CrossRef]
- Dumas, A.; Lercher, L.; Spicer, C.D.; Davis, B.G. Designing logical codon reassignment—Expanding the chemistry in biology. Chem. Sci. 2015, 6, 50–69. [Google Scholar] [CrossRef]
- Chin, J.W. Expanding and reprogramming the genetic code of cells and animals. Annu. Rev. Biochem. 2014, 83, 379–408. [Google Scholar] [CrossRef] [PubMed]
- Lajoie, M.J.; Söll, D.; Church, G.M. Overcoming challenges in engineering the genetic code. J. Mol. Biol. 2016, 428, 1004–1021. [Google Scholar] [CrossRef]
- Brown, W.; Liu, J.; Deiters, A. Genetic code expansion in animals. ACS Chem. Biol. 2018, 13, 2375–2386. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. Engineering the genetic code in cells and animals: Biological considerations and impacts. Acc. Chem. Res. 2017, 50, 2767–2775. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, H.; Kojima, K. Production of Bombyx mori silk fibroin incorporated with unnatural amino acids. Biomacromolecules 2014, 15, 2682–2690. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, H.; Amano, Y.; Iraha, F.; Kojima, K.; Ito, T.; Sakamoto, K. Genetic code expansion of the silkworm Bombyx mori to functionalize silk fiber. ACS Synth. Biol. 2018, 7, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Lang, K.; Chin, J.W. Cellular incorporation of unnatural amino acids and bioorthogonal labeling of proteins. Chem. Rev. 2014, 114, 4764–4806. [Google Scholar] [CrossRef]
- Teramoto, H.; Nakajima, K.; Kojima, K. Azide-incorporated clickable silk fibroin materials with the ability to photopattern. ACS Biomater. Sci. Eng. 2016, 2, 251–258. [Google Scholar] [CrossRef]
- Nikić, I.; Kang, J.H.; Girona, G.E.; Aramburu, I.V.; Lemke, E.A. Labeling proteins on live mammalian cells using click chemistry. Nat. Protoc. 2015, 10, 780. [Google Scholar] [CrossRef]
- Reddington, S.; Watson, P.; Rizkallah, P.; Tippmann, E.; Jones, D.D. Genetically encoding phenyl azide chemistry: New uses and ideas for classical biochemistry. Biochem. Soc. Trans. 2013, 41, 1177–1182. [Google Scholar] [CrossRef]
- Teramoto, H.; Iga, M.; Tsuboi, H.; Nakajima, K. Characterization and scaled-up production of azido-functionalized silk fiber produced by transgenic silkworms with an expanded genetic code. Int. J. Mol. Sci. 2019, 20, 616. [Google Scholar] [CrossRef]
- Ju, J.; Hu, N.; Cairns, D.M.; Liu, H.; Timko, B.P. Photo-cross-linkable, insulating silk fibroin for bioelectronics with enhanced cell affinity. Proc. Natl. Acad. Sci. USA 2020, 117, 15482–15489. [Google Scholar] [CrossRef] [PubMed]
- Kurland, N.E.; Dey, T.; Kundu, S.C.; Yadavalli, V.K. Precise patterning of silk microstructures using photolithography. Adv. Mater. 2013, 25, 6207–6212. [Google Scholar] [CrossRef]
- Xu, M.; Pradhan, S.; Agostinacchio, F.; Pal, R.K.; Greco, G.; Mazzolai, B.; Pugno, N.M.; Motta, A.; Yadavalli, V.K. Easy, scalable, robust, micropatterned silk fibroin cell substrates. Adv. Mater. Interfaces 2019, 6, 1801822. [Google Scholar] [CrossRef]
- Hu, X.; Shmelev, K.; Sun, L.; Gil, E.-S.; Park, S.-H.; Cebe, P.; Kaplan, D.L. Regulation of silk material structure by temperature-controlled water vapor annealing. Biomacromolecules 2011, 12, 1686–1696. [Google Scholar] [CrossRef] [PubMed]
- Tamada, Y.; Ikada, Y. Fibroblast growth on polymer surfaces and biosynthesis of collagen. J. Biomed. Mater. Res. 1994, 28, 783–789. [Google Scholar] [CrossRef]
- Takayama, Y.; Kusamori, K.; Nishikawa, M. Click chemistry as a tool for cell engineering and drug delivery. Molecules 2019, 24, 172. [Google Scholar] [CrossRef]
- Xi, W.; Scott, T.F.; Kloxin, C.J.; Bowman, C.N. Click chemistry in materials science. Adv. Funct. Mater. 2014, 24, 2572–2590. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teramoto, H.; Shirakawa, M.; Tamada, Y. Click Decoration of Bombyx mori Silk Fibroin for Cell Adhesion Control. Molecules 2020, 25, 4106. https://doi.org/10.3390/molecules25184106
Teramoto H, Shirakawa M, Tamada Y. Click Decoration of Bombyx mori Silk Fibroin for Cell Adhesion Control. Molecules. 2020; 25(18):4106. https://doi.org/10.3390/molecules25184106
Chicago/Turabian StyleTeramoto, Hidetoshi, Minori Shirakawa, and Yasushi Tamada. 2020. "Click Decoration of Bombyx mori Silk Fibroin for Cell Adhesion Control" Molecules 25, no. 18: 4106. https://doi.org/10.3390/molecules25184106
APA StyleTeramoto, H., Shirakawa, M., & Tamada, Y. (2020). Click Decoration of Bombyx mori Silk Fibroin for Cell Adhesion Control. Molecules, 25(18), 4106. https://doi.org/10.3390/molecules25184106