
Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review




Abstract
1. Introduction
2. Results
3. Benefits of Natural and Synthetic GSs and GSHP
3.1. Biofumigation/ Biocidal Activity and Pest Management
3.2. Antimicrobial Activity
3.3. Antioxidant Activity
3.4. Anti-Inflammatory Activity
3.5. Cholinesterase (ChE) Inhibitory Activities
3.6. Cytotoxic and Anti-Cancer Activity
3.7. Other Activities
3.8. Effects of GSs/GSHPs Used in Preclinical and Clinical Studies
3.9. Improving Plant GSs, GSHPs Content, Their Bioavailability, and Bioactivity
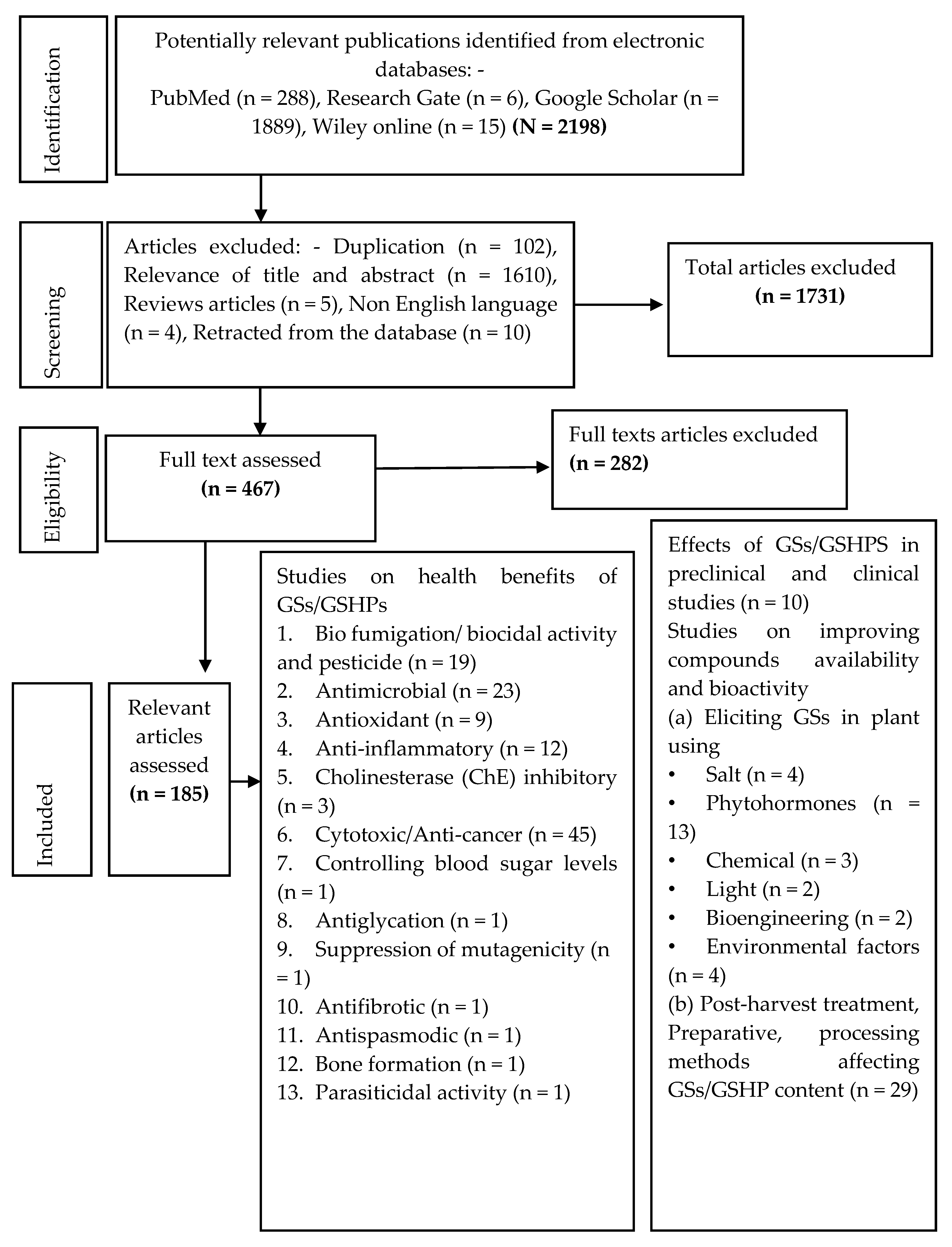
4. Materials and Methods
Search Strategy and Selection Criteria
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Heldt, H.-W.; Piechulla, B. Secondary metabolites fulfill specific ecological functions in plants. Plant Biochem. 2011, 399–408. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef] [PubMed]
- Borgen, B.H.; Ahuja, I.; Thangstad, O.P.; Honne, B.I.; Rohloff, J.; Rossiter, J.T.; Bones, A.M. “Myrosin cells” are not a prerequisite for aphid feeding on oilseed rape (Brassica napus) but affect host plant preferences. Plant Biol. 2012, 14, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Luang-In, V.; Narbad, A.; Nueno-Palop, C.; Mithen, R.; Bennett, M.; Rossiter, J.T. The metabolism of methylsulfinylalkyl- and methylthioalkyl-glucosinolates by a selection of human gut bacteria. Mol. Nutr. Food Res. 2014, 58, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Saladino, F.; Bordin, K.; Luciano, F.B.; Franzón, M.F.; Mañes, J.; Meca, G. Glucosinolates. Glucosinolates 2016. [Google Scholar] [CrossRef]
- Miao, H.; Wang, J.; Cai, C.; Chang, J.; Zhao, Y.; Wang, Q. Accumulation of Glucosinolates in Broccoli. In Glucosinolates; Mérillon, J.M., Ramawat, K.G., Eds.; Springer International Publishing: New York, NY, USA, 2017; pp. 431–461. [Google Scholar] [CrossRef]
- Ahuja, I.; Van Dam, N.M.; Winge, P.; Trælnes, M.; Heydarova, A.; Rohloff, J.; Langaas, M.; Bones, A.M. Plant defence responses in oilseed rape MINELESS plants after attack by the cabbage moth Mamestra brassicae. J. Exp. Bot. 2015, 66, 579–592. [Google Scholar] [CrossRef]
- Falk, K.L.; Kästner, J.; Bodenhausen, N.; Schramm, K.; Europe PMC Funders Group. The role of glucosinolates and the jasmonic acid pathway in resistance of Arabidopsis thaliana against molluskan herbivores. Mol. Ecol. 2016, 23, 1188–1203. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.C.; Ferreira, C.; Saavedra, M.J.; Simões, L.C.; Simões, M. Antibacterial activity and mode of action of selected glucosinolate hydrolysis products against bacterial pathogens. J. Food Sci. Technol. 2015, 52, 4737–4748. [Google Scholar] [CrossRef]
- Abdull Razis, A.F.; Noor, N.M. Sulforaphane is Superior to Glucoraphanin in Modulating Carcinogen-Metabolising Enzymes in Hep G2 Cells. Asian Pacific J. Cancer Prev. 2013, 14, 4235–4238. [Google Scholar] [CrossRef]
- Abdull Razis, A.F.; Bagatta, M.; De Nicola, G.R.; Iori, R.; Ioannides, C. Intact glucosinolates modulate hepatic cytochrome P450 and phase II conjugation activities and may contribute directly to the chemopreventive activity of cruciferous vegetables. Toxicology 2010, 277, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Berhow, M.A.; Angelino, D.; Jeffery, E.H. Camelina sativa defatted seed meal contains both alkyl sulfinyl glucosinolates and quercetin that synergize bioactivity. J. Agric. Food Chem. 2014, 62, 8385–8391. [Google Scholar] [CrossRef]
- Vale, A.P.; Santos, J.; Brito, N.V.; Fernandes, D.; Rosa, E.; Beatriz, M.; Oliveira, P.P. Evaluating the impact of sprouting conditions on the glucosinolate content of Brassica oleracea sprouts. Phytochemistry 2015, 115, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Carmen, S.; Tan, M.G.; Noel, M. Glucosinolates, Degradation Products and Myrosinase Activity in Raphanus sativus Linn. Pharmacogn. J. 2019. [Google Scholar] [CrossRef]
- Fechner, J.; Kaufmann, M.; Herz, C.; Eisenschmidt, D.; Lamy, E.; Kroh, L.W.; Hanschen, F.S. The major glucosinolate hydrolysis product in rocket (Eruca sativa L.), sativin, is 1,3-thiazepane-2-thione: Elucidation of structure, bioactivity, and stability compared to other rocket isothiocyanates. Food Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.R.; Kwak, J.H. Chemical composition and antioxidant activity in different tissues of brassica vegetables. Molecules 2015, 20, 1228–1243. [Google Scholar] [CrossRef]
- Ku, K.M.; Jeffery, E.H.; Juvik, J.A.; Kushad, M.M. Correlation of Quinone Reductase Activity and Allyl Isothiocyanate Formation among Different Genotypes and Grades of Horseradish Roots. J. Agric. Food Chem. 2015, 63, 2947–2955. [Google Scholar] [CrossRef]
- Lee, M.K.; Chun, J.H.; Byeon, D.H.; Chung, S.O.; Park, S.U.; Park, S.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, Y.P.; Kim, S.J. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp. pekinensis) and their antioxidant activity. LWT Food Sci. Technol. 2014, 58, 93–101. [Google Scholar] [CrossRef]
- Hahn, C.; Müller, A.; Kuhnert, N.; Albach, D. Diversity of Kale (Brassica oleracea var. sabellica): Glucosinolate Content and Phylogenetic Relationships. J. Agric. Food Chem. 2016. [Google Scholar] [CrossRef]
- Huseby, S.; Koprivova, A.; Lee, B.R.; Saha, S.; Mithen, R.; Wold, A.B.; Bengtsson, G.B.; Kopriva, S. Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 2013, 64, 1039–1048. [Google Scholar] [CrossRef]
- Kurepina, N.; Kreiswirth, B.N.; Mustaev, A. Growth-inhibitory activity of natural and synthetic isothiocyanates against representative human microbial pathogens. J. Appl. Microbiol. 2013, 115, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Lensing, C.J.; Forred, B.J.; Amolins, M.W.; Aegerter, C.L.; Vitiello, P.F.; Mays, J.R. Differentiating Antiproliferative and Chemopreventive Modes of Activity for Electron-Deficient Aryl Isothiocyanates against Human MCF-7 Cells. ChemMedChem 2018, 13, 1695–1710. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Feng, J.; Lin, K.; Zhang, X. Synthesis and Herbicidal Activity of Substituted Pyrazole Isothiocyanates. Molecules 2012, 17, 12187–12196. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Niu, J.; Wang, W.; Huo, H.; Li, J.; Luo, L.; Cao, Y. p-Aromatic Isothiocyanates: Synthesis and Anti Plant Pathogen Activity. Russ. J. Gen. Chem. 2018, 88, 1252–1257. [Google Scholar] [CrossRef]
- De Santi, M.; Galluzzi, L.; Lucarini, S.; Paoletti, M.F.; Fraternale, A.; Duranti, A.; De Marco, C.; Fanelli, M.; Zaffaroni, N.; Brandi, G.; et al. The indole-3-carbinol cyclic tetrameric derivative CTet inhibits cell proliferation via overexpression of p21/CDKN1A in both estrogen receptor-positive and triple-negative breast cancer cell lines. Breast Cancer Res. 2011, 13, R33. [Google Scholar] [CrossRef]
- Vo, Q.V.; Trenerry, C.; Rochfort, S.; Wadeson, J.; Leyton, C.; Hughes, A.B. Synthesis and anti-inflammatory activity of aromatic glucosinolates. Bioorganic Med. Chem. 2013, 21, 5945–5954. [Google Scholar] [CrossRef]
- Vo, Q.V.; Trenerry, C.; Rochfort, S.; Wadeson, J.; Leyton, C.; Hughes, A.B. Bioorganic & Medicinal Chemistry Synthesis and anti-inflammatory activity of indole glucosinolates. Bioorg. Med. Chem. 2014, 22, 856–864. [Google Scholar] [CrossRef]
- Vo, Q.V.; Rochfort, S.; Nam, P.C.; Nguyen, T.L.; Nguyen, T.T.; Mechler, A. Synthesis of aromatic and indole alpha-glucosinolates. Carbohydr. Res. 2018, 455, 45–53. [Google Scholar] [CrossRef]
- Psurski, M.; Błaewska, K.; Gajda, A.; Gajda, T.; Wietrzyk, J.; Oleksyszyn, J. Synthesis and antiproliferative activity of novel α- And β-dialkoxyphosphoryl isothiocyanates. Bioorganic Med. Chem. Lett. 2011, 21, 4572–4576. [Google Scholar] [CrossRef]
- Seo, M.S.; Jin, M.; Sohn, S.H.; Kim, J.S. Expression profiles of BrMYB transcription factors related to glucosinolate biosynthesis and stress response in eight subspecies of Brassica rapa. FEBS Open Bio 2017, 7, 1646–1659. [Google Scholar] [CrossRef]
- Community, ECommission Regulation (EEC) No 1470 / 68 on the drawing and reduction of Whereas the measures provided for in this Regulation are in accordance with the opinion of the Management Having regard to Council Regula. Off. J. Eur. Commun. 1990, 33, 27–34.
- Guo, L.; Yang, R.; Zhou, Y.; Gu, Z. Heat and hypoxia stresses enhance the accumulation of aliphatic glucosinolates and sulforaphane in broccoli sprouts. Eur. Food Res. Technol. 2016, 242, 107–116. [Google Scholar] [CrossRef]
- Tian, M.; Xu, X.; Liu, Y.; Xie, L.; Pan, S. Effect of Se treatment on glucosinolate metabolism and health-promoting compounds in the broccoli sprouts of three cultivars. Food Chem. 2016, 190, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Kumar, R.; Mahajan, J.; Vig, A.P.; Singh, B.; Singh, B.; Arora, S. 3-Butenyl isothiocyanate: A hydrolytic product of glucosinolate as a potential cytotoxic agent against human cancer cell lines. J. Food Sci. Technol. 2016, 53, 3437–3445. [Google Scholar] [CrossRef] [PubMed]
- Al-Gendy, A.A.; Nematallah, K.A.; Zaghloul, S.S.; Ayoub, N.A. Glucosinolates profile, volatile constituents, antimicrobial, and cytotoxic activities of Lobularia libyca. Pharm. Biol. 2016, 54, 3257–3263. [Google Scholar] [CrossRef]
- Li, Z.Y.; Wang, Y.; Shen, W.T.; Zhou, P. Content determination of benzyl glucosinolate and anti-cancer activity of its hydrolysis product in Carica papaya L. Asian Pac. J. Trop. Med. 2012, 5, 231–233. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Li, Y.; Wade, K.L.; Paonessa, J.D.; Fahey, J.W.; Zhang, Y. Allyl isothiocyanate-rich mustard seed powder inhibits bladder cancer growth and muscle invasion. Carcinogenesis 2010, 31, 2105–2110. [Google Scholar] [CrossRef]
- Boscaro, V.; Boffa, L.; Binello, A.; Amisano, G.; Fornasero, S.; Cravotto, G.; Gallicchio, M. Antiproliferative, proapoptotic, antioxidant and antimicrobial effects of Sinapis nigra L. And Sinapis alba L. Extracts. Molecules 2018, 23, 3004. [Google Scholar] [CrossRef]
- Rajan, T.S.; De Nicola, G.R.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Anticancer activity of glucomoringin isothiocyanate in human malignant astrocytoma cells. Fitoterapia 2016, 110, 1–7. [Google Scholar] [CrossRef]
- Antonini, E.; Iori, R.; Ninfali, P.; Scarpa, E.S. A Combination of Moringin and Avenanthramide 2f Inhibits the Proliferation of Hep3B Liver Cancer Cells Inducing Intrinsic and Extrinsic Apoptosis. Nutr. Cancer 2018, 70, 1159–1165. [Google Scholar] [CrossRef]
- Michl, C.; Vivarelli, F.; Weigl, J.; De Nicola, G.R.; Canistro, D.; Paolini, M.; Iori, R.; Rascle, A. The chemopreventive phytochemical moringin isolated from Moringa oleifera seeds inhibits JAK/STAT signaling. PLoS ONE 2016, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Van Loon, J.; Ruschioni, S.; De Nicola, G.R.; Olsen, C.E.; Iori, R.; Agerbirk, N. Taste detection of the non-volatile isothiocyanate moringin results in deterrence to glucosinolate-adapted insect larvae. Phytochemistry 2015, 118, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; De Nicola, G.R.; Iori, R.; Dell’Utri, P.; Bramanti, P.; Mazzon, E. Antibacterial activity of glucomoringin bioactivated with myrosinase against two important pathogens affecting the health of long-term patients in hospitals. Molecules 2013, 18, 14340–14348. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Rajan, T.S.; De Nicola, G.R.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. The isothiocyanate isolated from moringa oleifera shows potent anti-inflammatory activity in the treatment of murine subacute Parkinson’s disease. Rejuvenation Res. 2017, 20, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Blažević, I.; Đulović, A.; Čulić, V.Č.; Popović, M.; Guillot, X.; Burčul, F.; Rollin, P. Microwave-assisted versus conventional isolation of glucosinolate degradation products from Lunaria annua L. and their cytotoxic activity. Biomolecules 2020, 10, 215. [Google Scholar] [CrossRef]
- Meyer, S.L.F.; Zasada, I.A.; Orisajo, S.B.; Morra, M.J. Mustard seed meal mixtures: Management of Meloidogyne incognita on pepper and potential phytotoxicity. J. Nematol. 2011, 43, 7–15. [Google Scholar]
- Carpenter, E.L.; Le, M.N.; Miranda, C.L.; Reed, R.L.; Stevens, J.F.; Indra, A.K.; Ganguli-Indra, G. Photoprotective properties of isothiocyanate and nitrile glucosinolate derivatives from meadowfoam (Limnanthes alba) against UVB irradiation in human skin equivalent. Front. Pharmacol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Zasada, I.A.; Weiland, J.E.; Reed, R.L.; Stevens, J.F. Activity of meadowfoam (Limnanthes alba) seed meal glucolimnanthin degradation products against soilborne pathogens. J. Agric. Food Chem. 2012, 60, 339–345. [Google Scholar] [CrossRef]
- Intanon, S.; Reed, R.L.; Stevens, J.F.; Hulting, A.G.; Mallory-Smith, C.A. Identification and phytotoxicity of a new glucosinolate breakdown product from meadowfoam (Limnanthes alba) seed meal. J. Agric. Food Chem. 2014, 62, 7423–7429. [Google Scholar] [CrossRef]
- Mastelić, J.; Blaẑević, I.; Kosalec, I. Chemical composition and antimicrobial activity of volatiles from degenia velebitica, a european stenoendemic plant of the brassicaceae family. Chem. Biodivers. 2010, 7, 2755–2765. [Google Scholar] [CrossRef]
- Radonić, A.; Blažević, I.; Mastelić, J.; Zekić, M.; Skočibušić, M.; Maravić, A. Phytochemical analysis and antimicrobial activity of Cardaria draba (L.) desv. volatiles. Chem. Biodivers. 2011, 8, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Blažević, I.; Radonić, A.; Skočibušić, M.; Denicola, G.R.; Montaut, S.; Iori, R.; Rollin, P.; Mastelić, J.; Zekić, M.; Maravić, A. Glucosinolate profiling and antimicrobial screening of aurinia leucadea (Brassicaceae). Chem. Biodivers. 2011, 8, 2310–2321. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Maravić, A.; Čikeš Čulić, V.; Montaut, S.; Rollin, P. Antimicrobial and Cytotoxic Activities of Lepidium latifolium L. Hydrodistillate, Extract and Its Major Sulfur Volatile Allyl Isothiocyanate. Chem. Biodivers. 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.S.; Lazzeri, L.; Atkinson, H.J.; Urwin, P.E. Biofumigation for control of pale potato cyst nematodes: Activity of brassica leaf extracts and green manures on globodera pallida in vitro and in soil. J. Agric. Food Chem. 2011, 59, 7882–7890. [Google Scholar] [CrossRef]
- Hashem, F.A.; Motawea, H.; El-Shabrawy, A.E.; Shaker, K.; El-Sherbini, S. Myrosinase hydrolysates of Brassica oleraceae L. Var. italica reduce the risk of colon cancer. Phyther. Res. 2012, 26, 743–747. [Google Scholar] [CrossRef]
- Liu, A.G.; Juvik, J.A.; Jeffery, E.H.; Berman-Booty, L.D.; Clinton, S.K.; Erdman, J.W. Enhancement of broccoli indole glucosinolates by methyl jasmonate treatment and effects on prostate carcinogenesis. J. Med. Food 2014, 17, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Venkatesan, R.; Kim, S.Y. Neuroprotective and anti-inflammatory activities of allyl isothiocyanate through attenuation of JNK/NF-κB/TNF-α signaling. Int. J. Mol. Sci. 2017, 18, 1423. [Google Scholar] [CrossRef]
- Dekić, M.S.; Radulović, N.S.; Stojanović, N.M.; Randjelović, P.J.; Stojanović-Radić, Z.Z.; Najman, S.; Stojanović, S. Spasmolytic, antimicrobial and cytotoxic activities of 5-phenylpentyl isothiocyanate, a new glucosinolate autolysis product from horseradish (Armoracia rusticana P. Gaertn., B. Mey. & Scherb., Brassicaceae). Food Chem. 2017, 232, 329–339. [Google Scholar] [CrossRef]
- Popović, M.; Maravić, A.; Čulić, V.Č.; Đulović, A.; Burčul, F.; Blažević, I. Biological effects of glucosinolate degradation products from horseradish: A horse that wins the race. Biomolecules 2020, 10, 343. [Google Scholar] [CrossRef]
- Kulisic-Bilusic, T.; Schmöller, I.; Schnäbele, K.; Siracusa, L.; Ruberto, G. The anticarcinogenic potential of essential oil and aqueous infusion from caper (Capparis spinosa L.). Food Chem. 2012, 132, 261–267. [Google Scholar] [CrossRef]
- Lozano-Baena, M.D.; Tasset, I.; Obregón-Cano, S.; De Haro-Bailon, A.; Muñoz-Serrano, A.; Alonso-Moraga, Á. Antigenotoxicity and tumor growing inhibition by leafy Brassica carinata and sinigrin. Molecules 2015, 20, 15748–15765. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, D.; Omirou, M.; Liadaki, K.; Tsikou, D.; Delis, C.; Garagounis, C.; Krokida, A.; Zambounis, A.; Papadopoulou, K.K. Glucosinolate biosynthesis in Eruca sativa. Plant Physiol. Biochem. 2016, 109, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Samson, M.; Porter, N.; Orekoya, O.; Hebert, J.R.; Adams, S.A.; Bennett, C.L.; Steck, S.E. Direct and Indirect Antioxidant Activity of Polyphenol- and Isothiocyanate-Enriched Fractions from Moringa oleifera Tugba. J. Agric. Food Chem. 2017, 155, 3–12. [Google Scholar] [CrossRef]
- Aissani, N.; Tedeschi, P.; Maietti, A.; Brandolini, V.; Garau, V.L.; Caboni, P. Nematicidal activity of allylisothiocyanate from horseradish (Armoracia rusticana) roots against Meloidogyne incognita. J. Agric. Food Chem. 2013, 61, 4723–4727. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Park, H.; Hyun, H.; Kim, J.; Kim, H.; Oh, H.I.; Hwang, H.S.; Kim, D.K.; Kim, H.H. Effects of glucosinolates from Turnip (Brassica rapa L.) root on bone formation by human osteoblast-like MG-63 cells and in normal young rats. Phyther. Res. 2015, 29, 902–909. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Burčul, F.; Ljubenkov, I.; Ruščić, M.; Generalić Mekinić, I. Bunias erucago L.: Glucosinolate profile and in vitro biological potential. Molecules 2019, 24, 741. [Google Scholar] [CrossRef]
- Kestwal, R.M.; Lin, J.C.; Bagal-Kestwal, D.; Chiang, B.H. Glucosinolates fortification of cruciferous sprouts by sulphur supplementation during cultivation to enhance anti-cancer activity. Food Chem. 2011, 126, 1164–1171. [Google Scholar] [CrossRef]
- Ibrahim, M.D.; Kntayya, S.B.; Mohd Ain, N.; Iori, R.; Ioannides, C.; Razis, A.F.A. Induction of apoptosis and cytotoxicity by raphasatin in human breast adenocarcinoma MCF-7 cells. Molecules 2018, 23, 3092. [Google Scholar] [CrossRef]
- Miao, H.Y.; Wang, M.Y.; Chang, J.Q.; Tao, H.; Sun, B.; Wang, Q.M. Effects of glucose and gibberellic acid on glucosinolate content and antioxidant properties of Chinese kale sprouts. J. Zhejiang Univ. Sci. B 2017, 18, 1093–1100. [Google Scholar] [CrossRef]
- Scholl, C.; Eshelman, B.D.; Barnes, D.M.; Hanlon, P.R. Raphasatin Is a More Potent Inducer of the Detoxification Enzymes Than Its Degradation Products. J. Food Sci. 2011, 76, C504–C511. [Google Scholar] [CrossRef]
- Sotelo, T.; Lema, M.; Soengas, P.; Cartea, M.E.; Velasco, P. In Vitro Activity of Glucosinolates and Their Degradation Products against Brassica-Pathogenic Bacteria and Fungi. Appl. Environ. Microbiol. 2015, 81, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Bangarwa, S.K.; Norsworthy, J.K. Herbicidal activity of three isothiocyanates against yellow nutsedge and their dissipation under two plastic mulches. Crop Prot. 2015, 74, 145–149. [Google Scholar] [CrossRef]
- Kaur, T.; Bhat, R.; Khajuria, M.; Vyas, R.; Kumari, A.; Nadda, G.; Vishwakarma, R.; Vyas, D. Dynamics of glucosinolate-myrosinase system during Plutella xylostella interaction to a novel host Lepidium latifolium L. Plant Sci. 2016, 250, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, A.R.; Hulagabali, C.V.; Adhav, A.S.; Joshi, R.S. Mechanistic insight in potential dual role of sinigrin against Helicoverpa armigera. Phytochemistry 2018, 145, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Witzel, K.; Hanschen, F.S.; Schreiner, M.; Krumbein, A.; Ruppel, S.; Grosch, R. Verticillium Suppression Is Associated with the Glucosinolate Composition of Arabidopsis thaliana Leaves. PLoS ONE 2013, 8, e71877. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kurt, Ş.; Güneş, U.; Soylu, E.M. In vitro and in vivo antifungal activity of synthetic pure isothiocyanates against Sclerotinia sclerotiorum. Pest Manag. Sci. 2011, 67, 869–875. [Google Scholar] [CrossRef]
- Rivera-Vega, L.J.; Krosse, S.; de Graaf, R.M.; Garvi, J.; Garvi-Bode, R.D.; van Dam, N.M. Allelopathic effects of glucosinolate breakdown products in Hanza [Boscia senegalensis (Pers.) Lam.] processing waste water. Front. Plant Sci. 2015. [Google Scholar] [CrossRef]
- Hinds, L.; Kenny, O.; Hossain, M.B.; Walsh, D.; Sheehy, E.; Evans, P.; Gaffney, M.; Rai, D.K. Evaluating the Antibacterial Properties of Polyacetylene and Glucosinolate Compounds with Further Identification of Their Presence within Various Carrot (Daucus carota) and Broccoli (Brassica oleracea) Cultivars Using High-Performance Liquid Chromatograph. J. Agric. Food Chem. 2017, 65, 7186–7191. [Google Scholar] [CrossRef]
- Choi, K.D.; Kim, H.Y.; Shin, I.S. Antifungal activity of isothiocyanates extracted from horseradish (Armoracia rusticana) root against pathogenic dermal fungi. Food Sci. Biotechnol. 2017, 26, 847–852. [Google Scholar] [CrossRef]
- Park, H.W.; Choi, K.D.; Shin, I.S. Antimicrobial activity of isothiocyanates(itcs) extracted from horseradish (Armoracia rusticana) root against oral microorganisms. Biocontrol Sci. 2013, 18, 163–168. [Google Scholar] [CrossRef]
- Dias, C.; Aires, A.; Saavedra, M.J. Antimicrobial Activity of Isothiocyanates from Cruciferous Plants against Methicillin-Resistant Staphylococcus aureus (MRSA). IJMS 2014, 15, 19552–19561. [Google Scholar] [CrossRef]
- Conrad, A.; Biehler, D.; Nobis, T.; Richter, H.; Engels, I.; Biehler, K.; Frank, U. Broad spectrum antibacterial activity of a mixture of isothiocyanates from nasturtium (Tropaeoli majoris herba) and horseradish (Armoraciae rusticanae radix). Drug Res. (Stuttg). 2013, 63, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.; Borges, A.; Dias, C.; Aires, A.; Bennett, R.; Rosa, E.; Simões, M. Antimicrobial Activity of Phenolics and Glucosinolate Hydrolysis Products and their Synergy with Streptomycin against Pathogenic Bacteria. Med. Chem. (Los. Angeles). 2012, 6, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.J.; Mutters, N.T.; Blessing, B.; Günther, F. Natural isothiocyanates express antimicrobial activity against developing and mature biofilms of Pseudomonas aeruginosa. Fitoterapia 2017, 119, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Herzallah, S.; Lledó, M.L.; Holley, R. Influence of NaCl and NaNO3 on sinigrin hydrolysis by foodborne bacteria. J. Food Prot. 2011, 74, 2162–2168. [Google Scholar] [CrossRef]
- Hifnawy, M.S.; Salam, R.M.A.; Rabeh, M.A.; Aboseada, M.A. Glucosinolates, Glycosidically Bound Volatiles and Antimicrobial Activity of Brassica oleraceae Var. Botrytis, (Soultany Cultivar). Arch. biochem. Biophys. 2013, 3, 66–82. [Google Scholar]
- Dufour, V.; Alazzam, B.; Ermel, G.; Thepaut, M.; Rossero, A.; Tresse, O.; Baysse, C. Antimicrobial activities of isothiocyanates against Campylobacter jejuni isolates. Front. Cell. Infect. Microbiol. 2012, 2, 53. [Google Scholar] [CrossRef]
- Aires, A.; Dias, C.; Rosa, E.; Saavedra, M. Antimicrobial Susceptibility of Aeromonas Spp. Isolated from Pig Ileum Segments to Natural Isothiocyanates. Med. Chem. (Los. Angeles). 2013, 9, 861–866. [Google Scholar] [CrossRef]
- Blažević, I.; Radonić, A.; Mastelić, J.; Zekić, M.; Skočibušić, M.; Maravić, A. Glucosinolates, glycosidically bound volatiles and antimicrobial activity of Aurinia sinuata (Brassicaceae). Food Chem. 2010, 121, 1020–1028. [Google Scholar] [CrossRef]
- Malabed, R.S.; Noel, M.G.; Aton III, B.C.; Toribio, E.A.F. Characterization of the Glucosinolates and Isothiocyanates in Mustard (Brassica juncea L.) Extracts and Determination of Its Myrosinase Activity and Antioxidant Capacity. In Proceedings of the DLSU Research Congress 2014, Manila, Phillipines, 6–8 March 2014; pp. 1–7. [Google Scholar]
- Biegańska-Marecik, R.; Radziejewska-Kubzdela, E.; Marecik, R. Characterization of phenolics, glucosinolates and antioxidant activity of beverages based on apple juice with addition of frozen and freeze-dried curly kale leaves (Brassica oleracea L. var. acephala L.). Food Chem. 2017, 230, 271–280. [Google Scholar] [CrossRef]
- Lee, H.W.; Lee, C.G.; Rhee, D.K.; Um, S.H.; Pyo, S. Sinigrin inhibits production of inflammatory mediators by suppressing NF-κB/MAPK pathways or NLRP3 inflammasome activation in macrophages. Int. Immunopharmacol. 2017, 45, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Shirai, Y.; Fujita, Y.; Hashimoto, R.; Ohi, K.; Yamamori, H.; Yasuda, Y.; Ishima, T.; Suganuma, H.; Ushida, Y.; Takeda, M.; et al. Dietary intake of sulforaphane-rich broccoli sprout extracts during juvenile and adolescence can prevent phencyclidine-induced cognitive deficits at adulthood. PLoS ONE 2015, 10, e0127244. [Google Scholar] [CrossRef] [PubMed]
- Shiina, A.; Kanahara, N.; Sasaki, T.; Oda, Y.; Hashimoto, T.; Hasegawa, T.; Yoshida, T.; Iyo, M.; Hashimoto, K. An open study of sulforaphane-rich broccoli sprout extract in patients with schizophrenia. Clin. Psychopharmacol. Neurosci. 2015, 13, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Galuppo, M.; Iori, R.; De Nicola, G.R.; Bramanti, P.; Mazzon, E. The protective effects of bioactive (RS)-glucoraphanin on the permeability of the mice blood-brain barrier following experimental autoimmune encephalomyelitis. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 194–204. [Google Scholar]
- Galuppo, M.; Giacoppo, S.; De Nicola, G.R.; Iori, R.; Mazzon, E.; Bramanti, P. RS-Glucoraphanin bioactivated with myrosinase treatment counteracts proinflammatory cascade and apoptosis associated to spinal cord injury in an experimental mouse model. J. Neurol. Sci. 2013, 334, 88–96. [Google Scholar] [CrossRef]
- Blazevic, I.; Burcul, F.; Ruscic, M.; Mastelic, J. Glucosinolates, volatile constituents, and acetylcholinesterase inhibitory activity of Alyssoides utriculata. Chem. Nat. Compd. 2013, 49, 374–378. [Google Scholar] [CrossRef]
- Kadir, N.H.A.; David, R.; Rossiter, J.T.; Gooderham, N.J. The selective cytotoxicity of the alkenyl glucosinolate hydrolysis products and their presence in Brassica vegetables. Toxicology 2015, 334, 59–71. [Google Scholar] [CrossRef]
- Bassan, P.; Bhushan, S.; Kaur, T.; Arora, R.; Arora, S.; Vig, A.P. Extraction, profiling and bioactivity analysis of volatile glucosinolates present in oil extract of Brassica juncea var. raya. Physiol. Mol. Biol. Plants 2018, 24, 399–409. [Google Scholar] [CrossRef]
- Srivastava, M.M.; Bansal, P.; Medhe, S.; Ganesh, N. In vitro anticancer activity of dietary bioagent (isothiocyanates) on HepG2 and B16F10 cell lines: A comparative study. Ann. Plant Sci. 2013, 234–237. [Google Scholar]
- Radošević, K.; Srček, V.G.; Bubalo, M.C.; Rimac Brnčić, S.; Takács, K.; Redovniković, I.R. Assessment of glucosinolates, antioxidative and antiproliferative activity of broccoli and collard extracts. J. Food Compos. Anal. 2017, 61, 59–66. [Google Scholar] [CrossRef]
- Abdull Razis, A.F.; Iori, R.; Ioannides, C. The natural chemopreventive phytochemical R-sulforaphane is a far more potent inducer of the carcinogen-detoxifying enzyme systems in rat liver and lung than the S-isomer. Int. J. Cancer 2011, 128, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Abdull Razis, A.F.; De Nicola, G.R.; Pagnotta, E.; Iori, R.; Ioannides, C. A glucosinolate-rich extract of Japanese Daikon perturbs carcinogen-metabolizing enzyme systems in rat, being a potent inducer of hepatic glutathione S-transferase. Eur. J. Nutr. 2013, 52, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Jakubikova, J.; Cervi, D.; Ooi, M.; Kim, K.; Nahar, S.; Klippel, S.; Cholujova, D.; Leiba, M.; Daley, J.F.; Delmore, J.; et al. Anti-tumor activity and signaling events triggered by the isothiocyanates, sulforaphane and phenethyl isothiocyanate, in multiple myeloma. Haematologica 2011, 96, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Sun, B.; Miao, H.; Cai, C.; Xu, C.; Wang, Q. Variation of glucosinolates and quinone reductase activity among different varieties of Chinese kale and improvement of glucoraphanin by metabolic engineering. Food Chem. 2015, 168, 321–326. [Google Scholar] [CrossRef]
- Ku, K.M.; Kim, M.J.; Jeffery, E.H.; Kang, Y.H.; Juvik, J.A. Profiles of Glucosinolates, Their Hydrolysis Products, and Quinone Reductase Inducing Activity from 39 Arugula (Eruca sativa Mill.) Accessions. J. Agric. Food Chem. 2016, 64, 6524–6532. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Cho, H.J.; Chung, F.L.; Wang, X.; Hoe, H.S.; Park, K.K.; Kim, C.H.; Chang, H.W.; Lee, S.R.; Chang, Y.C. Isothiocyanates suppress the invasion and metastasis of tumors by targeting FAK/MMP-9 activity. Oncotarget 2017, 8, 63949–63962. [Google Scholar] [CrossRef]
- Veprik, A.; Khanin, M.; Linnewiel-Hermoni, K.; Danilenko, M.; Levy, J.; Sharoni, Y. Polyphenols, isothiocyanates, and carotenoid derivatives enhance estrogenic activity in bone cells but inhibit it in breast cancer cells. Am. J. Physiol. Endocrinol. Metab. 2012, 303, 815–824. [Google Scholar] [CrossRef][Green Version]
- Hać, A.; Brokowska, J.; Rintz, E.; Bartkowski, M.; Węgrzyn, G.; Herman-Antosiewicz, A. Mechanism of selective anticancer activity of isothiocyanates relies on differences in DNA damage repair between cancer and healthy cells. Eur. J. Nutr. 2019, 59, 421–1432. [Google Scholar] [CrossRef]
- Vauzour, D.; Buonfiglio, M.; Corona, G.; Chirafisi, J.; Vafeiadou, K.; Angeloni, C.; Hrelia, S.; Hrelia, P.; Spencer, J.P.E. Sulforaphane protects cortical neurons against 5-S-cysteinyl-dopamine-induced toxicity through the activation of ERK1/2, NrF-2 and the upregulation of detoxification enzymes. Mol. Nutr. Food Res. 2010, 54, 532–542. [Google Scholar] [CrossRef]
- Rajendran, P.; Delage, B.; Dashwood, W.M.; Yu, T.W.; Wuth, B.; Williams, D.E.; Ho, E.; Dashwood, R.H. Histone deacetylase turnover and recovery in sulforaphane-treated colon cancer cells: Competing actions of 14-3-3 and Pin1 in HDAC3/SMRT corepressor complex dissociation/reassembly. Mol. Cancer 2011, 10, 68. [Google Scholar] [CrossRef]
- Tafakh, M.S.; Saidijam, M.; Ranjbarnejad, T.; Malih, S.; Mirzamohammadi, S.; Najafi, R. Sulforaphane, a Chemopreventive Compound, Inhibits Cyclooxygenase-2 and Microsomal Prostaglandin e Synthase-1 Expression in Human HT-29 Colon Cancer Cells. Cells Tissues Organs 2019, 206, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Curran, K.M.; Bracha, S.; Wong, C.P.; Beaver, L.M.; Stevens, J.F.; Ho, E. Sulforaphane absorption and histone deacetylase activity following single dosing of broccoli sprout supplement in normal dogs. Vet. Med. Sci. 2018, 4, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Kntayya, S.B.; Ibrahim, M.D.; Ain, N.M.; Iori, R.; Ioannides, C.; Abdull Razis, A.F. Induction of apoptosis and cytotoxicity by isothiocyanate sulforaphene in human hepatocarcinoma HepG2 cells. Nutrients 2018, 10, 718. [Google Scholar] [CrossRef] [PubMed]
- Abdull Razis, A.F.; De Nicola, G.R.; Pagnotta, E.; Iori, R.; Ioannides, C. 4-Methylsulfanyl-3-butenyl isothiocyanate derived from glucoraphasatin is a potent inducer of rat hepatic phase II enzymes and a potential chemopreventive agent. Arch. Toxicol. 2012, 86, 183–194. [Google Scholar] [CrossRef]
- Förster, N.; Mewis, I.; Glatt, H.; Haack, M.; Brigelius-Flohé, R.; Schreiner, M.; Ulrichs, C. Characteristic single glucosinolates from: Moringa oleifera: Induction of detoxifying enzymes and lack of genotoxic activity in various model systems. Food Funct. 2016, 7, 4660–4674. [Google Scholar] [CrossRef]
- Kim, J.; Bang, H.; Ahn, M.; Choi, Y.; Kim, G.O.; Shin, T. Allyl isothiocyanate reduces liver fibrosis by regulating Kupffer cell activation in rats. J. Vet. Med. Sci. 2018, 80, 893–897. [Google Scholar] [CrossRef]
- Awasthi, S.; Saraswathi, N.T. Sinigrin, a major glucosinolate from cruciferous vegetables restrains non-enzymatic glycation of albumin. Int. J. Biol. Macromol. 2016, 83, 410–415. [Google Scholar] [CrossRef]
- Baenas, N.; Piegholdt, S.; Schloesser, A.; Moreno, D.A.; García-Viguera, C.; Rimbach, G.; Wagner, A.E. Metabolic activity of radish sprouts derived isothiocyanates in drosophila melanogaster. Int. J. Mol. Sci. 2016, 17, 251. [Google Scholar] [CrossRef]
- Hanlon, N.; Konsue, N.; Coldham, N.; Sauer, M.J.; Ioannides, C. Exposure to isothiocyanates suppresses urinary mutagenicity in rats treated with heterocyclic amine IQ: Lack of association with CYP1 activity. Nutr. Cancer 2011, 63, 300–305. [Google Scholar] [CrossRef]
- Galletti, S.; Bagatta, M.; Branca, F.; Argento, S.; De Nicola, G.R.; Cianchetta, S.; Iori, R.; Ninfali, P. Isatis canescens is a rich source of glucobrassicin and other health-promoting compounds. J. Sci. Food Agric. 2015, 95, 158–164. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Szaefer, H.; Bartoszek, A.; Baer-Dubowska, W. Modulation of rat hepatic and kidney phase II enzymes by cabbage juices: Comparison with the effects of indole-3-carbinol and phenethyl isothiocyanate. Br. J. Nutr. 2011, 105, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Hajra, S.; Patra, A.R.; Basu, A.; Bhattacharya, S. Prevention of doxorubicin (DOX)-induced genotoxicity and cardiotoxicity: Effect of plant derived small molecule indole-3-carbinol (I3C) on oxidative stress and inflammation. Biomed. Pharmacother. 2018, 101, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Hajra, S.; Basu, A.; Singha Roy, S.; Patra, A.R.; Bhattacharya, S. Attenuation of doxorubicin-induced cardiotoxicity and genotoxicity by an indole-based natural compound 3,3′-diindolylmethane (DIM) through activation of Nrf2/ARE signaling pathways and inhibiting apoptosis. Free Radic. Res. 2017, 51, 812–827. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Schulz, M.; Pagnotta, E.; Ugolini, L.; Yang, T.; Matthes, A.; Lazzeri, L.; Agerbirk, N. The Role of the Glucosinolate-Myrosinase System in Mediating Greater Resistance of Barbarea verna than B. vulgaris to Mamestra brassicae Larvae. J. Chem. Ecol. 2018, 44, 1190–1205. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.; Borges, A.; Simoes, L.; Saavedra, M.; Simoes, M. Antibacterial Activity of Phenyl Isothiocyanate on Escherichia coli and Staphylococcus aureus. Med. Chem. (Los. Angeles) 2013, 9, 756–761. [Google Scholar] [CrossRef]
- Martín, J.C.; Higuera, B.L. Glucosinolate composition of Colombian accessions of mashua (Tropaeolum tuberosum Ruíz & Pavón), structural elucidation of the predominant glucosinolate and assessment of its antifungal activity. J. Sci. Food Agric. 2016, 96, 4702–4712. [Google Scholar] [CrossRef]
- Burčul, F.; Generalić Mekinić, I.; Radan, M.; Rollin, P.; Blažević, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzyme Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef]
- Lee, Y.M.; Cho, H.J.; Ponnuraj, S.P.; Kim, J.; Kim, J.; Kim, G.S.; Park, J.H. Phenethyl isothiocyanate inhibits 12-0-tetradecanoylphorbol-13-acetate-induced inflammatory responses in mouse skin. J. Med. Food 2011, 14, 377–385. [Google Scholar] [CrossRef]
- Rajan, T.S.; Giacoppo, S.; Iori, R.; De Nicola, G.R.; Grassi, G.; Pollastro, F.; Bramanti, P.; Mazzon, E. Anti-inflammatory and antioxidant effects of a combination of cannabidiol and moringin in LPS-stimulated macrophages. Fitoterapia 2016, 112, 104–115. [Google Scholar] [CrossRef]
- Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Shi, Y.; Liu, H.; Panjwani, A.A.; Warrick, C.R.; Olson, M.E. A strategy to deliver precise oral doses of the glucosinolates or isothiocyanates from moringa oleifera leaves for use in clinical studies. Nutrients 2019, 11, 1547. [Google Scholar] [CrossRef]
- Malabed, R.S.; Noel, M.G. Characterization of the glucosinolates and isothiocyanates in Malunggay (Moringa oleifera L.) Extracts and determinationof their myrosinae activity and anticancer properties. In Proceedings of the DLSU Research Congress 2013, Manila, Phillipines, 7–9 March 2013; pp. 1–6. [Google Scholar]
- Yu, C.; Gong, A.Y.; Chen, D.; Solelo Leon, D.; Young, C.Y.; Chen, X.M. Phenethyl isothiocyanate inhibits androgen receptor-regulated transcriptional activity in prostate cancer cells through suppressing PCAF. Mol. Nutr. Food Res. 2013, 57, 1825–1833. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.; Gan, N.; Chung, F.L. Isothiocyanates inhibit proteasome activity and proliferation of multiple myeloma cells. Carcinogenesis 2011, 32, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Larkin, R.P.; Honeycutt, C.W.; Olanya, O.M. Management of Verticillium Wilt of Potato with Disease-Suppressive Green Manures and as Affected by Previous Cropping History. Plant Dis. 2011, 95, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Schlaeppi, K.; Abou-Mansour, E.; Buchala, A.; Mauch, F. Disease resistance of Arabidopsis to Phytophthora brassicae is established by the sequential action of indole glucosinolates and camalexin. Plant J. 2010, 62, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.K.; Tsukamoto, C.; Kim, K.W.; Choi, M.R. Investigation of glucosinolates, and the antioxidant activity of Dolsan leaf mustard kimchi extract using HPLC and LC-PDA-MS/MS. J. Food Biochem. 2017, 41. [Google Scholar] [CrossRef]
- Chester, K.; Zahiruddin, S.; Ahmad, A.; Khan, W.; Paliwal, S.; Ahmad, S. Bioautography-based Identification of Antioxidant Metabolites of Solanum nigrum L. and Exploration. Pharmacogn. Mag. 2017, 15, 104. [Google Scholar] [CrossRef]
- Nguyen Phan, T.X. A Density Functional Theory Study of Antioxidant Activity of Isothiocyanates in Broccoli Sprouts (Brassica Oleracea L.). Vietnam J. Sci. Technol. 2018, 54, 306. [Google Scholar] [CrossRef]
- Eichholzer, M.; Steinbrecher, A.; Kaaks, R.; Teucher, B.; Linseisen, J.; Rohrmann, S. Effects of selenium status, dietary glucosinolate intake and serum glutathione S-transferase α activity on the risk of benign prostatic hyperplasia. BJU Int. 2012, 110, 879–885. [Google Scholar] [CrossRef]
- Psurski, M.; Wietrzyk, J.; Oleksyszyn, J. 1035 Studies over the Natural Isothiocyanates Structure-activity and Structure-mechanism of Action Relationship. Eur. J. Cancer 2012, 48, S249. [Google Scholar] [CrossRef]
- Skugor, S.; Jodaa Holm, H.; Bjelland, A.K.; Pino, J.; Evensen, Ø.; Krasnov, A.; Wadsworth, S. Nutrigenomic effects of glucosinolates on liver, muscle and distal kidney in parasite-free and salmon louse infected Atlantic salmon. Parasites Vectors 2016, 9, 639. [Google Scholar] [CrossRef] [PubMed]
- Tahata, S.; Singh, S.V.; Lin, Y.; Hahm, E.R.; Beumer, J.H.; Christner, S.M.; Rao, U.N.; Sander, C.; Tarhini, A.A.; Tawbi, H.; et al. Evaluation of biodistribution of sulforaphane after administration of oral broccoli sprout extract in melanoma patients with multiple atypical nevi. Cancer Prev. Res. 2018, 11, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Tohidi, M.; Nazeri, P.; Mehran, M.; Azizi, F.; Mirmiran, P. Effect of broccoli sprouts on insulin resistance in type 2 diabetic patients: A randomized double-blind clinical trial. Int. J. Food Sci. Nutr. 2012, 63, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Traka, M.H.; Melchini, A.; Coode-Bate, J.; Kadhi, O.A.; Saha, S.; Defernez, M.; Troncoso-Rey, P.; Kibblewhite, H.; O’Neill, C.M.; Bernuzzi, F.; et al. Transcriptional changes in prostate of men on active surveillance after a 12-mo glucoraphanin-rich broccoli intervention-results from the Effect of Sulforaphane on prostate CAncer PrEvention (ESCAPE) randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1133–1144. [Google Scholar] [CrossRef]
- Alumkal, J.J.; Slottke, R.; Schwartzman, J.; Cherala, G.; Munar, M.; Graff, J.N.; Beer, T.M.; Ryan, C.W.; Koop, D.R.; Gibbs, A.; et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Invest. New Drugs 2015, 33, 480–490. [Google Scholar] [CrossRef]
- Brown, R.H.; Reynolds, C.; Brooker, A.; Talalay, P.; Fahey, J.W. Sulforaphane improves the bronchoprotective response in asthmatics through Nrf2-mediated gene pathways. Respir. Res. 2015, 16, 106. [Google Scholar] [CrossRef]
- Wise, R.A.; Holbrook, J.T.; Criner, G.; Sethi, S.; Rayapudi, S.; Sudini, K.R.; Sugar, E.A.; Burke, A.; Thimmulappa, R.; Singh, A.; et al. Lack of effect of oral sulforaphane administration on Nrf2 expression in COPD: A randomized, double-blind, placebo controlled trial. PLoS ONE 2016, 11, e0163716. [Google Scholar] [CrossRef]
- Atwell, L.L.; Zhang, Z.; Mori, M.; Farris, P.E.; Vetto, J.T.; Naik, A.M.; Oh, K.Y.; Thuillier, P.; Ho, E.; Shannon, J. Sulforaphane bioavailability and chemopreventive activity in women scheduled for breast biopsy. Cancer Prev. Res. 2015, 8, 1184–1191. [Google Scholar] [CrossRef]
- Ma, L.; Liu, G.; Sampson, L.; Willett, W.C.; Hu, F.B.; Sun, Q. Dietary glucosinolates and risk of type 2 diabetes in 3 prospective cohort studies. Am. J. Clin. Nutr. 2018, 107, 617–625. [Google Scholar] [CrossRef]
- Ma, L.; Liu, G.; Zong, G.; Sampson, L.; Hu, F.B.; Willett, W.C.; Rimm, E.B.; Manson, J.E.; Rexrode, K.M.; Sun, Q. Intake of glucosinolates and risk of coronary heart disease in three large prospective cohorts of US men and women. Clin. Epidemiol. 2018, 10, 749–762. [Google Scholar] [CrossRef]
- Banerjee, A.; Variyar, P.S.; Chatterjee, S.; Sharma, A. Effect of post harvest radiation processing and storage on the volatile oil composition and glucosinolate profile of cabbage. Food Chem. 2014, 151, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Bagchi, B.; Misra, L.K.; Deka, B.C. Changes in post-harvest phytochemical qualities of broccoli florets during ambient and refrigerated storage. Food Chem. 2011, 127, 1510–1514. [Google Scholar] [CrossRef]
- Villarreal-García, D.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as biofactories: Postharvest stress-induced accumulation of phenolic compounds and glucosinolates in broccoli subjected to wounding stress and exogenous phytohormones. Front. Plant Sci. 2016, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S. Effect of cooking method on antioxidant compound contents in cauliflower. Prev. Nutr. Food Sci. 2019, 24, 210–216. [Google Scholar] [CrossRef]
- Sut, S.; Boschiero, I.; Solana, M.; Malagoli, M.; Bertucco, A.; Dall’Acqua, S. Supercritical CO2 extraction of eruca sativa using cosolvents: Phytochemical composition by LC-MS analysis. Molecules 2018, 23, 3240. [Google Scholar] [CrossRef]
- Rafińska, K.; Pomastowski, P.; Rudnicka, J.; Krakowska, A.; Maruśka, A.; Narkute, M.; Buszewski, B. Effect of solvent and extraction technique on composition and biological activity of Lepidium sativum extracts. Food Chem. 2019, 289, 16–25. [Google Scholar] [CrossRef]
- Luciano, F.B.; Belland, J.; Holley, R.A. Microbial and chemical origins of the bactericidal activity of thermally treated yellow mustard powder toward Escherichia coli O157:H7 during dry sausage ripening. Int. J. Food Microbiol. 2011, 145, 69–76. [Google Scholar] [CrossRef]
- Yuan, G.; Wang, X.; Guo, R.; Wang, Q. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts. Food Chem. 2010, 121, 1014–1019. [Google Scholar] [CrossRef]
- Guo, R.F.; Yuan, G.F.; Wang, Q.M. Effect of NaCl treatments on glucosinolate metabolism in broccoli sprouts. J. Zhejiang Univ. Sci. B 2013, 14, 124–131. [Google Scholar] [CrossRef]
- Del Carmen Rodríguez-Hernández, M.; Moreno, D.A.; Carvajal, M.; Del Carmen Martínez-Ballesta, M. Genotype influences sulfur metabolism in broccoli (Brassica oleracea L.) under elevated CO2 and NaCl stress. Plant Cell Physiol. 2014, 55, 2047–2059. [Google Scholar] [CrossRef]
- Ku, K.M.; Jeffery, E.H.; Juvik, J.A. Exogenous methyl jasmonate treatment increases glucosinolate biosynthesis and quinone reductase activity in kale leaf tissue. PLoS ONE 2014, 9, e103407. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yang, R.; Gu, Z. Cloning of genes related to aliphatic glucosinolate metabolism and the mechanism of sulforaphane accumulation in broccoli sprouts under jasmonic acid treatment. J. Sci. Food Agric. 2016, 96, 4329–4336. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.C.; Juvik, J.A.; Ku, K.M. Targeted metabolomic and transcriptomic analyses of “red russian” kale (Brassicae napus var. pabularia) following methyl jasmonate treatment and larval infestation by the cabbage looper (Trichoplusia ni hübner). Int. J. Mol. Sci. 2018, 19, 1058. [Google Scholar] [CrossRef] [PubMed]
- Ku, K.M.; Jeffery, E.H.; Juvik, J.A. Optimization of methyl jasmonate application to broccoli florets to enhance health-promoting phytochemical content. J. Sci. Food Agric. 2014, 94, 2090–2096. [Google Scholar] [CrossRef]
- Ku, K.M.; Jeffery, E.H.; Juvik, J.A. Influence of seasonal variation and methyl jasmonate mediated induction of glucosinolate biosynthesis on quinone reductase activity in broccoli florets. J. Agric. Food Chem. 2013, 61, 9623–9631. [Google Scholar] [CrossRef]
- Ku, K.M.; Choi, J.H.; Kushad, M.M.; Jeffery, E.H.; Juvik, J.A. Pre-harvest Methyl Jasmonate Treatment Enhances Cauliflower Chemoprotective Attributes Without a Loss in Postharvest Quality. Plant Foods Hum. Nutr. 2013, 68, 113–117. [Google Scholar] [CrossRef]
- Wiesner, M.; Schreiner, M.; Glatt, H. High mutagenic activity of juice from pak choi (Brassica rapa ssp. chinensis) sprouts due to its content of 1-methoxy-3-indolylmethyl glucosinolate, and its enhancement by elicitation with methyl jasmonate. Food Chem. Toxicol. 2014, 67, 10–16. [Google Scholar] [CrossRef]
- Kim, M.J.; Chiu, Y.C.; Kim, N.K.; Park, H.M.; Lee, C.H.; Juvik, J.A.; Ku, K.M. Cultivar-specific changes in primary and secondary metabolites in pak choi (Brassica rapa, Chinensis group) by methyl jasmonate. Int. J. Mol. Sci. 2017, 18, 1004. [Google Scholar] [CrossRef]
- Zhang, K.; Su, H.; Zhou, J.; Liang, W.; Liu, D.; Li, J. Overexpressing the Myrosinase Gene TGG1 Enhances Stomatal Defense Against Pseudomonas syringae and Delays Flowering in Arabidopsis. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Schiavon, M.; Berto, C.; Malagoli, M.; Trentin, A.; Sambo, P.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Selenium biofortification in radish enhances nutritional quality via accumulation of methyl-selenocysteine and promotion of transcripts and metabolites related to glucosinolates, phenolics amino acids. Front. Plant Sci. 2016. [Google Scholar] [CrossRef]
- Marino, D.; Ariz, I.; Lasa, B.; Santamaría, E.; Fernández-Irigoyen, J.; González-Murua, C.; Aparicio Tejo, P.M. Quantitative proteomics reveals the importance of nitrogen source to control glucosinolate metabolism in Arabidopsis thaliana and Brassica oleracea. J. Exp. Bot. 2016, 67, 3313–3323. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Yang, Z.; Chen, X.; Jin, P.; Wang, X.; Zheng, Y. 6-Benzylaminopurine Delays Senescence and Enhances Health-Promoting Compounds of Harvested Broccoli. J. Agric. Food Chem. 2012, 60, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Chen, X.; Yang, Z.; Jin, P.; Wang, K.; Shang, H.; Wang, X.; Zheng, Y. Maintaining quality and bioactive compounds of broccoli by combined treatment with 1-methylcyclopropene and 6-benzylaminopurine. J. Sci. Food Agric. 2013, 93, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Yao, D.; Xu, F.; Wang, H.; Zheng, Y. Effect of light on quality and bioactive compounds in postharvest broccoli florets. Food Chem. 2015, 172, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Sontowski, R.; Gorringe, N.J.; Pencs, S.; Schedl, A.; Touw, A.J.; van Dam, N.M. Same Difference? Low and High Glucosinolate Brassica rapa Varieties Show Similar Responses Upon Feeding by Two Specialist Root Herbivores. Front. Plant Sci. 2019, 10, 1451. [Google Scholar] [CrossRef]
- Touw, A.J.; Verdecia Mogena, A.; Maedicke, A.; Sontowski, R.; van Dam, N.M.; Tsunoda, T. Both Biosynthesis and Transport Are Involved in Glucosinolate Accumulation During Root-Herbivory in Brassica rapa. Front. Plant Sci. 2020, 10, 1653. [Google Scholar] [CrossRef]
- Rozpadek, P.; Ślesak, I.; Cebula, S.; Waligórski, P.; Dziurka, M.; Skoczowski, A.; Miszalski, Z. Ozone fumigation results in accelerated growth and persistent changes in the antioxidant system of Brassica oleracea L. var. capitata f. alba. J. Plant Physiol. 2013, 170, 1259–1266. [Google Scholar] [CrossRef]
- Stotz, H.U.; Sawada, Y.; Shimada, Y.; Hirai, M.Y.; Sasaki, E.; Krischke, M.; Brown, P.D.; Saito, K.; Kamiya, Y. Role of camalexin, indole glucosinolates, and side chain modification of glucosinolate-derived isothiocyanates in defense of Arabidopsis against Sclerotinia sclerotiorum. Plant J. 2011, 67, 81–93. [Google Scholar] [CrossRef]
- Kato, M.; Imayoshi, Y.; Iwabuchi, H.; Shimomura, K. Kinetic changes in glucosinolate-derived volatiles by heat-treatment and myrosinase activity in nakajimana (Brassica rapa L. cv. nakajimana). J. Agric. Food Chem. 2011, 59, 11034–11039. [Google Scholar] [CrossRef]
- Atwell, L.L.; Hsu, A.; Wong, C.P.; Stevens, J.F.; Bella, D.; Yu, W.; Pereira, C.B.; Löhr, C.V.; Christensen, J.M.; Roderick, H. Multiple rare alleles at LDLR and APOA5 confer risk for early- onset myocardial infarction. Nature 2015, 518, 102–106. [Google Scholar] [CrossRef]
- Okunade, O.; Niranjan, K.; Ghawi, S.K.; Kuhnle, G.; Methven, L. Supplementation of the Diet by Exogenous Myrosinase via Mustard Seeds to Increase the Bioavailability of Sulforaphane in Healthy Human Subjects after the Consumption of Cooked Broccoli. Mol. Nutr. Food Res. 2018, 62, 1–6. [Google Scholar] [CrossRef]
- Ghawi, S.K.; Methven, L.; Niranjan, K. The potential to intensify sulforaphane formation in cooked broccoli (Brassica oleracea var. italica) using mustard seeds (Sinapis alba). Food Chem. 2013, 138, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, R.P.; Wu, C.; Holley, R.A. Contribution of endogenous plant myrosinase to the antimicrobial activity of deodorized mustard against Escherichia coli O157:H7 in fermented dry sausage. Int. J. Food Microbiol. 2014, 189, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Shen, Y.; Zhu, Y.; Mupunga, J.; Zou, L.; Liu, C.; Liu, S.; Mao, J. Broccoli ingestion increases the glucosinolate hydrolysis activity of microbiota in the mouse gut. Int. J. Food Sci. Nutr. 2019, 70, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Totušek, J.; Tříska, J.; Lefnerová, D.; Strohalm, J.; Vrchotová, N.; Zendulka, O.; Průchová, J.; Chaloupková, J.; Novotná, P.; Houška, M. Contents of sulforaphane and total isothiocyanates, antimutagenic activity, and inhibition of clastogenicity in pulp juices from Cruciferous plants. Czech J. Food Sci. 2011, 29, 548–556. [Google Scholar] [CrossRef]
- Okunade, O.A.; Ghawi, S.K.; Methven, L.; Niranjan, K. Thermal and pressure stability of myrosinase enzymes from black mustard (Brassica nigra L. W.D.J. Koch. var. nigra), brown mustard (Brassica juncea L. Czern. var. juncea) and yellow mustard (Sinapsis alba L. subsp. maire) seeds. Food Chem. 2015, 187, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.; Aguilar-Galvez, A.; García-Ríos, D.; Chirinos, R.; Limaymanta, E.; Pedreschi, R. Postharvest storage and cooking techniques affect the stability of glucosinolates and myrosinase activity of Andean mashua tubers (Tropaeolum tuberosum). Int. J. Food Sci. Technol. 2019, 54, 2387–2395. [Google Scholar] [CrossRef]
- Lee, J.G.; Lim, S.; Kim, J.; Lee, E.J. The mechanism of deterioration of the glucosinolate-myrosynase system in radish roots during cold storage after harvest. Food Chem. 2017, 233, 60–68. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Kühn, C.; Nickel, M.; Rohn, S.; Dekker, M. Leaching and degradation kinetics of glucosinolates during boiling of Brassica oleracea vegetables and the formation of their breakdown products. Food Chem. 2018, 263, 240–250. [Google Scholar] [CrossRef]
- Radziejewska-Kubzdela, E.; Olejnik, A.; Biegańska-Marecik, R. Effect of pretreatment on bioactive compounds in wild rocket juice. J. Food Sci. Technol. 2019, 56, 5234–5242. [Google Scholar] [CrossRef]
- Niu, Y.; Rogiewicz, A.; Wan, C.; Guo, M.; Huang, F.; Slominski, B.A. Effect of microwave treatment on the efficacy of expeller pressing of Brassica napus rapeseed and Brassica juncea mustard seeds. J. Agric. Food Chem. 2015, 63, 3078–3084. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Sharma, A.; Rai, P.K.; Kumar, D. Effect of microwave pre-treatment on quality parameters in Indian mustard. J. Food Sci. Technol. 2019, 56, 4956–4965. [Google Scholar] [CrossRef] [PubMed]
- Rybarczyk-Plonska, A.; Hagen, S.F.; Borge, G.I.A.; Bengtsson, G.B.; Hansen, M.K.; Wold, A.B. Glucosinolates in broccoli (Brassica oleracea L. var. italica) as affected by postharvest temperature and radiation treatments. Postharvest Biol. Technol. 2016, 116, 16–25. [Google Scholar] [CrossRef]


{kind=link}
| GSs | Hydrolysis Product | Biological Activity | Source | Organism/Pathogen/ Cell | Reference |
|---|---|---|---|---|---|
| Aliphatic GSs | |||||
| Biocidal/ Biofumigation Activity | |||||
| Glucoraphanin, glucoiberin glucobrassicanapin, glucoerucin | Sulforaphane 4-pentenyl ITCs | Biocide | Synthetic | [72] | |
| Sinigrin | Allyl ITCs/2-propenyl ITCs | Herbicidal Larval feeding deterrent | Synthetic L. latifolium L. | Cyperus esculentus weed Plutella xylostella pest | [73,74] |
| Nematicidal activity | A. rusticana | M. incognita | [65] | ||
| Insecticidal | synthetic | Helicoverpa armigera | [75] | ||
| Biocide | Synthetic | [72] | |||
| Nematicidal activity | B. juncea L. | Globodera pallida | [55] | ||
| Biofumigation | A. thaliana | Verticillium longisporum | [76] | ||
| Fungitoxic activity | Synthetic | Sclerotinia sclerotiorum | [77] | ||
| Gluconapin | Butyl ITCs | Fungitoxic activity | Synthetic | S. sclerotiorum | [77] |
| Biocide | [72] | ||||
| Glucocapparin | Methyl ITCs | Fungitoxic activity | Synthetic | S. sclerotiorum | [77] |
| Weed germination inhibition | B. senegalensis | [78] | |||
| Antimicrobial activity | |||||
| Sinigrin | Antimicrobial | S. alba L. and S. nigra L. | S. aureus, Streptococcus pyogenes, Bacillus cereus, Escherichia coli, P. aeruginosa, Candida albicans | [39] | |
| Antibacterial | B. oleracea | Bacillus cereus, E. coli, Salmonella typhimurium, Methicilin resistant S. aureus | [79] | ||
| Antifungal | A. rusticana | Trichophyton rubrum, Trichophyton mentagrophytes, Microsporum canis, Epidermophyton floccosum | [80] | ||
| Antimicrobial | A. rusticana | C. albicans, Fusobacterium nucleatum | [81] | ||
| Antimicrobial | Cruciferous plant | methicillin resistant S. aureus | [82] | ||
| Antimicrobial and cytotoxic | L. latifolum L. | Listeria monocytogenes, Acinetobacter baumannii, C. albicans, S. aureus, S. Typhimurium, E. coli, P. aeruginosa | [54] | ||
| Synthetic | Haemophilus influenzae, Moraxella catarrhalis, Serratia marcescens, Proteus vulgaris, and Candida species E. coli, P. aeruginosa, S. aureus and L. monocytogenes P. aeruginosa | [10,83,84,85] | |||
| Antimicrobial | Synthetic | E. coli | [86] | ||
| Antimicrobial activity | B. oleracea | E. coli, Klebsiella pneumoniae | [87] | ||
| Antimicrobial | A. rusticana | C. albicans, F. nucleatum | [81] | ||
| Bactericidal | synthetic | Campylobacter jejuni | [88] | ||
| Glucoraphanin | 4-(Methylsulfanyl)butyl ITCs & 5-(methylsulfanyl)pentanenitrile | Antimicrobial | C. draba L. | E. coli, K. pneumonia,Enterobacter sakazakii, P. aeruginosa, Cronobacter spp., S. aureus, Rhizopus stolonifer | [52] |
| Antibacterial | Synthetic | Pig aeromonas intestinal bacteria | [89] | ||
| Gluconapin | 3-Butenyl ITCs | Antimicrobial | A. leucadea | Bacillus cereus, C. albicans, Penicillium sp., R. stolonifera, P. aeruginosa | [53] |
| Antimicrobial activity | B. oleracea | E. coli, K. pneumoniae | [87] | ||
| Antimicrobial | A. rusticana | C. albicans, F. nucleatum | [81] | ||
| Glucoerucin | 4-(Methylsulfanyl)butyl ITCs, 5-(methylsulfanyl)pentanenitrile | Antimicrobial | C. draba L. | E. coli, K. pneumonia,E. sakazakii, P. aeruginosa, Cronobacter spp., S. aureus, R. stolonifer | [52] |
| Antimicrobial activity | B. oleracea | E. coli, K. pneumoniae | [87] | ||
| Antimicrobial | L. libyca | C. albicans and P. aeruoginosa | [36] | ||
| Glucobrassicanapin | Antimicrobial | Aurinia sinuate | Gram positive, negative bacteria and fungi | [90] | |
| A. leucadea | B. cereus, C. albicans, Penicillium sp., R. stolonifera, P. aeruginosa | [53] | |||
| Glucoalyssin | Antimicrobial | A. leucadea | B. cereus, C. albicans, Penicillium sp., R. stolonifera, P. aeruginosa | [53] | |
| Glucoiberverin | Antimicrobial activity | B. oleracea | E. coli, K. pneumoniae | [87] | |
| Glucoberteroin | 6-(Methylsulfanyl) hexanenitrile | Antimicrobial | D. velebitica | [51] | |
| aliphatic ITCs and their derivatives | Antimicrobial | Synthetic | Mycobacterium tuberculosis | [22] | |
| Antioxidant activity | |||||
| Sinigrin | Allyl ITCs | Antioxidant | B. juncea L. | [91] | |
| B. rapa L. | [19] | ||||
| Curly kale leaves | [92] | ||||
| Gluconapin | Antioxidant | B. juncea L. B. rapa L. | [19,91] | ||
| Glucoalyssin, progoitrin, glucobrassicanapin, | Antioxidant | B. rapa L. | [19] | ||
| Glucoiberin | Antioxidant | Curly kale leaves | [92] | ||
| Anti-inflammatory activity | |||||
| Sinigrin | Allyl ITCs | Anti-inflammatory | Synthetic | [93] | |
| W. koreana | [58] | ||||
| Glucoraphanin | Sulforaphane | Inflammatory Prophylactic | Broccoli sprouts | [94] | |
| Anti-inflammatory | Broccoli sprouts | [95] | |||
| Tuscan blackkale | [96] | ||||
| B. oleracea | [97] | ||||
| Neoglucobrassicin | Anti-inflammatory | synthetic | [28] | ||
| Cholinesterase Inhibitory Activity | |||||
| Gluconapin, Glucoerucin Glucoraphanin | But-3-enyl ITCs erucin sulforaphane | Acetylcholinesterase inhibitory activity | Alyssoides utriculata | [98] | |
| Cytotoxic Activity and chemoprevention | |||||
| Sinigrin | Allyl ITCs | Quinone reductase activity | A. rusticana | Hepa1c1c7 murine hepatoma cells | [18] |
| Cytotoxicity | Synthetic | MCL-5 cells | [99] | ||
| Anti-cancer, radical scavenging ability and increase ROS | B. juncea var. raya | human breast cancer (MCF-7, MDA-MB-231), prostate cancer (PC-3), lung cancer (A-549), cervical cancer (HeLa) and colon cancer (HCT 116) cells | [100] | ||
| Anti-cancer | Mustard seed powder | bladder cancer cell lines and orthotopic rats model bladder | [38] | ||
| Cytotoxic | L. latifolum L. | Glioblastoma LN229 cells | [54] | ||
| Anti-cancer, anti-melanoma | E. sativa seed oil | HepG2 human liver carcinoma and BB16F10 mice melanoma cells | [101] | ||
| Anti-tumor | Collard | Human MCF-7, HeLa cells | [102] | ||
| Anti-proliferative | S. alba L. and S. nigra L. | colon HCT 116 and HT-29 cells | [39] | ||
| Anti-tumor | Collard | Human MCF-7 and HeLa cells | [102] | ||
| Glucoraphanin | Sulforaphane | Chemoprevention | B. oleracea | Liver and lung cells | [103] |
| Chemoprevention | Japanese Daikon | Rats liver and lung | [104] | ||
| Anti-multiple myeloma activity | Synthetic | Myeloma cells | [105] | ||
| Chemoprevention | B. oleracea | Human HepG2 hepatoma cells | [11] | ||
| Anti-cancer, anti-melanoma | E. sativa seed oil | HepG2 human liver carcinoma, BB16F10 mice melanoma cells | [101] | ||
| Quinone reductase activity | Chinese kales | Hepa 1c1c7 murine hepatoma cells | [106] | ||
| Quinone reductase activity | E. sativa Mill | murine hepatoma Hepa1c1c7 cells | [107] | ||
| Anti-metastatic | Synthetic | MDA-MB-231(breast), Caski (cervical), A549 (lung), and U2OS (osteosarcoma) cell lines | [108] | ||
| Anti-cancer | Synthetic | breast cancer cell | [109] | ||
| inhibit cancer cells DNA replication | Synthetic | PC-3 prostate cancer cells | [110] | ||
| upregulating detoxification enzymes | Synthetic | Mouse cortical neurons | [111] | ||
| Chemoprevention | Japanese Daikon | Rats liver and lung | [104] | ||
| Chemoprotection | Synthetic | Human colon cancer cells | [112,113] | ||
| histone deacetylase activity | Broccoli sproutssupplement | Dog | [114] | ||
| Glucoraphenin | Sulforaphene | Cytotoxic and cell apoptosis | R. sativus seeds | Human hepatocarcinoma HepG2 cells | [115] |
| Glucoraphasatin | Raphasitin | Enzyme detoxification | R. sativus | HepG2 cells | [71] |
| Chemoprevention | Human breast MCF-7 cells | [69] | |||
| Chemoprevention | Japanese Daikon | Rats liver and lung | [104] | ||
| Chemo preventive | Daikon sprouts | Rats’ hepatic enzymes | [116] | ||
| Gluconapin | 3-Butenyl ITCs | Cytotoxicity | B. juncea L. | Human prostate cancer cells | [35] |
| Cytotoxicity | Synthetic | MCL-5 cells | [99] | ||
| Anti-cancer, radical scavenging ability and increase ROS | B. juncea var. raya | human breast cancer (MCF-7,MDA-MB-231), prostate cancer (PC-3), lung cancer (A-549), cervical cancer (HeLa) and colon cancer (HCT 116) cells | [100] | ||
| Glucoiberin | Cytotoxic activity | B. oleracea | colon cancer cells | [56] | |
| Glucocapparin | Methyl ITCs | Chemoprevention | Capparis spinosa L. | HT-29 cell | [61] |
| Glucoarabin, Glucocamelinin | phase II detoxification enzyme induction, quinone reductase | C. sativa L. Crantz | Hepa1c1c7 cells | [13] | |
| α-4-Rhamnopyranosyloxy-benzyl GSs, & isomers | Induction of detoxifying enzymes | M. oleifera | human hepatocellular carcinoma cell line -HepG2 cells | [117] | |
| Glucoiberverin, Glucoerucin Glucoiberin | Tumor growth inhibition, Antimicrobial | L. libyca | HL60 (human promyelocytic leukaemia cell lines) C. albicans, P. aeruoginosa | [36] | |
| Other activities | |||||
| Sinigrin | Allyl ITCs | Antifibrotic activity | Synthetic | [118] | |
| Antiglycation | Synthetic | [119] | |||
| Glucoraphenin, Glucoraphasatin | Sulforaphene, Raphasatin | Blood sugar control | R. sativus | [120] | |
| Glucoraphanin, Glucoerucin | Sulforaphane, Erucin | Suppress mutagenecity | Synthetic | [121] | |
| progoitrin, glucoraphanin, glucoalyssin, gluconapin, glucoerucin, glucoberteroin, glucobrassicanapin | Bone formation | B. rapa L. | [66] | ||
| Indole GSs | |||||
| Biocidal activity | |||||
| Glucobrassicin | Indole-3 carbinol | Biocide | Synthetic | [72] | |
| Antioxidant activity | |||||
| Glucobrassicin | Antioxidant | B. rapa L. | [19] | ||
| Curly kale leaves | [92] | ||||
| Isatis canescens | [122] | ||||
| Chemoprevention | |||||
| Glucobrassicin | Indole-3-carbinol | Chemoprevention | Synthetic | Rats’ hepatic and kidney mice bone marrow cells | [123,124] |
| 3,3′-Diindolylmethane | Enhances level of reduced-glutathione | Synthetic | Rats heart and bone marrow | [125] | |
| Other activities | |||||
| Glucobrassicin, neoglucobrassicin | Bone formation | B. rapa L. | [66] | ||
| Aromatic GSs | |||||
| Gluconasturtiin | Phenethyl ITCs | Biocide | Synthetic | [72] | |
| Larvicidal | Barbarea verna and Barbarea vulgaris | Mamestra brassicae | [126] | ||
| Glucotropaeolin, Sinalbin | Benzyl ITCs | Biocide | Synthetic | [72] | |
| Glucolimnanthin | 3-ethoxybenzyl ITCs | Bio-herbicide Biopesticide | L. alba | Lettuce Meloidogyne hapla, Pythium irregulare and Verticillium dahliae | [49,50] |
| GMG | Moringin | Insect deterrence | M. oleifera seeds | Pieris napi, Athalia rosae, Pieris brassicae | [43] |
| Phenyl ITCs | Herbicidal | Synthetic | C. esculentus weed | [73] | |
| Antimicrobial activity | |||||
| GMG | Moringin | Antimicrobial | M. oleifera | P. aeruginosa, S. aureus | [44] |
| Gluconasturtiin | Phenylethyl ITCs | Antifungal | A. rusticana | T. rubrum, T. mentagrophytes, M. canis, E. floccosum | [80] |
| Antimicrobial | A. rusticanaroots | C. albicans, F. nucleatum | [81] | ||
| Antibacterial | Synthetic | E. coli, S. aureus | [10,127] | ||
| Antimicrobial | Cruciferous plant | methicillin resistant S. aureus | [82] | ||
| Antimicrobial | Synthetic Tropaeoli majoris, A. rusticanae | Pig aeromonas intestinal bacteria P. aeruginosa H. influenzae, M. catarrhalis, S. marcescens, P. vulgaris and Candida species | [83,84,85,89] | ||
| Antifungal | Synthetic | S. sclerotiorum | [77] | ||
| Glucotropaeolin | Benzyl ITCs | Bactericidal | Synthetic | C. jejuni | [88] |
| Antimicrobial | Synthetic | Antimicrobial | [82] | ||
| Antimicrobial | Pig aeromonas bacteria | [89] | |||
| Antifungal | Synthetic | S. sclerotiorum | [77] | ||
| Glucobarbarin | Larvicidal | B. verna and B. vulgaris | M. brassicae | [126] | |
| p-Methoxybenzyl, p-hydroxybenzyl GS | Antifungal | Tropaeolumtuberosum | Phytophthora infestans, R. solani | [128] | |
| 5- Phenylpentyl ITCs | Antimicrobial, cytotoxic & antispasmodic | A. rusticana | S. aureus, B. subtilis, B. cereus, S. enterica, P. vulgaris, E. coli, C. albicans Aspergillus brasiliensis | [59] | |
| Antioxidant activity | |||||
| GMG | Antioxidant | M. oleifera | [64] | ||
| Anti-inflammatory activity | |||||
| Phenyl ITCs | Anti-inflammatory | Synthetic | [129] | ||
| Gluconasturtiin | Phenethyl ITCs | Anti-inflammatory | Synthetic | [130] | |
| GMG | Moringin | Anti-inflammatory | M. oleifera | [96,131,132] | |
| 3,4-Dimethoxyphenyl | Anti-inflammatory | Synthetic | [27] | ||
| 3-methoxyphenyl ITCS | Anti-inflammatory | synthetic | [129] | ||
| Cytotoxicity and chemoprevention | |||||
| GMG | cytotoxicity | M. oleifera | human colon adenocarcinoma grade II cells | [133] | |
| Inhibit cells proliferation | Hep3B Liver Cancer Cells | [41] | |||
| Anticancer | human malignant astrocytoma cell | [40] | |||
| Gluconasturtiin | Phenethyl ITCs | Anti-cancer, radical scavenging ability and increase ROS | B. juncea var. raya | human breast cancer (MCF-7 and MDA-MB-231), prostate cancer (PC-3), lung cancer (A-549), cervical cancer (HeLa) and colon cancer (HCT 116) lines | [100] |
| Chemoprevention | Synthetic | Rats’ hepatic and kidney | [123] | ||
| Chemoprevention | Synthetic | prostate cancer cells | [134] | ||
| Anti-cancer, anti-melanoma | E. sativa seed oil | HepG2 human liver carcinoma and BB16F10 mice melanoma cell line | [101] | ||
| Inhibit multiple myeloma growth | Synthetic | multiple myeloma cells | [135] | ||
| Glucotropaeolin | Benzyl ITCs | Anticancer activity | Carica papaya L. | human lung cancer H69 cell | [37] |
| Inhibit growth | Synthetic | multiple myeloma cells | [135] | ||
| Benzyl and Phenyl ITCs | Antimetastatic | Synthetic | MDA-MB-231 (breast), Caski (cervical), A549 (lung), and U2OS (osteosarcoma) cell lines | [108] | |
| A, β-dialkoxyphosphoryl alkyl & aralkyl ITCs | Antiproliferative activity | Synthetic | Lung cancer cells | [30] | |
| 3-Methoxybenzyl ITCs, | Photoprotective | L. alba | human skin cells | [48] | |
| 5- Phenylpentyl ITCs | Antimicrobial, cytotoxic & antispasmodic | A. rusticana | S. aureus, B. subtilis, B. cereus, S. enterica, P. vulgaris, E. coli, C. albicans A. brasiliensis | [59] | |
| Other activities | |||||
| Gluconasturtiin | Bone formation | B. rapa L. | [66] | ||
| Phenethyl ITCs | Suppress mutagenecity | Synthetic | [121] | ||
| Phenyl ITCs | Anti-inflammatory, cholinesterase inhibitory | Synthetic | [129] | ||
| Patient’s Condition | Participants Enrolled | Groups and Doses | Duration | Analysed | Clinical Outcomes | Results | Reference |
|---|---|---|---|---|---|---|---|
| Melanoma | 17 | 3 groups, 3 oral doses (50,100, 200 µmoles) of broccoli sprouts extract containing sulforaphane (SFN) | 28 days | 17 | sulforaphane levels in plasma and skin, plasma cytokines, Safety, tissue proteomics | Detectable SFN in plasma, cytokines decreased, extracts tolerated up to 200 µmol, increased tumor suppressors | [145] |
| Type 2 diabetes patients | 81 | 3 groups (2 received 10 g/day (d) broccoli sprout powder (BSP) and 5 g/d BSP and the third received the placebo | 4 weeks | 72 | Insulin concentration, fasting serum glucose, glucose: insulin ratio, homeostasis model of insulin resistance index | 10 g/d BSP significantly decreased serum insulin concentration, and improved insulin resistance in patients | [146] |
| Low and intermediate risk prostate cancer patients | 61 | 3 groups received each 300 mL of different broccoli soup (rich in glucoraphanin) weekly | 12 months | 48 | Tissues examination/prostate pathology and RNA sequencing analysis for gene expression | Reduction of prostate cancer progression. Soups with high glucoraphanin content caused suppression in expression. | [147] |
| Patients with recurrent prostate cancer | 20 | 200 µmoles/day of sulforaphane rich extract broccoli sprouts | 20 weeks | 16 | Safety of doses, Prostate specific antigen (PSA)% levels | Treatment was safe, exhibited anti-tumor potential, majority of patients didn’t get reduced PSA levels | [148] |
| Asthmatic patients | 51 | 100 µmoles/day of sulforaphane rich broccoli sprouts extract | 14 days | 45 | Pulmonary functions, NAD(P)H quinone dehydrogenase-1 gene expression, safety | Enhanced bronchoconstrictor effects, increased gene expression also enhanced broncho-protection | [149] |
| Women with abnormal mammograms | 54 | 2 groups; group1: (2 pills, 3 times/day of 30 mg glucoraphanin), group 2 placebo | 2 to 8 weeks | 48 | Sulforaphane in blood and urine, tissue biomarkers | Safe but not efficient to produce the changes in breast tissue tumor biomarkers | [151] |
| Schizophrenia | 10 | 3 tablets/30 mg of Sulforaphane per day | 8 weeks | 7 | Evaluation of Symptoms using positive and negative syndrome scale and cognitive function | Sulforaphane potentially improved cognitive function | [95] |
| Treatment/factor | Plant | GSs | Biological Activity | Reference |
|---|---|---|---|---|
| Sodium chloride | Radish sprouts Broccoli sprouts | Total GSs, Glucoraphasatin Sulforaphane | Antioxidant activity - | [161,162] |
| Salinity and carbon dioxide | B oleracea | Indolic GSs, aliphatic GSs respectively | - | [163] |
| Sulphur salt supplementation | B. oleracea Var. capitata, R. sativus | Total GSs, progoitrin, glucoerucin, glucobrassicin, glucohirsutin and 4- methoxybrassicin | Antioxidant, anti- proliferative | [68] |
| MeJA | B. napus, B.oleracea | gluconasturtiin, glucobrassicin, neoglucobrassicin, glucoraphanin | Quinone reductase | [164] |
| Broccoli florets | glucobrassicin, neoglucobrassicin, gluconasturtiin | Quinone reductase | [169] | |
| B. rapa ssp. Chinensis B. rapa L. B. napus var. pabularia B. oleracea | 1-methoxy-3-indolylmethyl GSs Indole GSs Total GSs, sulforaphane glucobrassicin, neoglucobrassicin, gluconasturtiin | Mutagenecity - - - | [57,166,171,170] | |
| MeJA, high temperature, water stress | Broccoli cultivars | Total GSs | Quinone reductase | [168] |
| GA3 with glucose | B. oleracea | Indolic GSs | Antioxidant | [70] |
| JA | B. oleracea var. italica | Glucoraphanin | Not evaluated | [165] |
| Selenium | Broccoli sprouts R. sativus | Sulforaphane Glucoraphanin | Not evaluated | [34] [173] |
| Nitrogen nutrition (Ammonium) | A. thaliana and B. oleraceaVar. italica | GSs | Not evaluated | [174] |
| Ozone | Brassica oleracea var. capitata f. alba | Sinigrin | Antioxidant | [180] |
| 6-benzylaminopurine | B. oleracea | Total GSs and sulforaphane | Antioxidant | [175,176] |
| Green LED light | B. oleracea | Total GSs | [177] | |
| Larval infestation Delia radicum root herbivory D. radicum and Delia floralis | B. napus var. pabularia B. rapa L. B. rapa varieties | Indole GSs Benzyl and indole GSs in roots Aliphatic, indole and benzyl GSs in roots of high GSs varieties | [166,178,179] | |
| BoTGG1 Gene | A. thaliana | GSs | Bacterial pathogens attack | [172] |
| Treatment | Source | Effects | Biological Activity | |
|---|---|---|---|---|
| Mild heat treatment | B. rapa L. | Myrosinase enzyme activity | [182] | |
| Continuous consumption | Broccoli | Total ITCs in colon and caecum levels of male mice | Quinone oxidoreductase | [187] |
| Exogenous myrosinase treatment | Broccoli sprouts Cooked broccoli Mustard powder | Sulforaphane in diet Sulforaphane in cooked vegetables Enzyme activity | Sulforaphane Plasma levels Urinary sulforaphane levels Antimicrobial | [183,184,186] |
| Powdered mustard seeds | B. oleracea var. italica | Sulforaphane in processed vegetables | - | [185] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maina, S.; Misinzo, G.; Bakari, G.; Kim, H.-Y. Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules 2020, 25, 3682. https://doi.org/10.3390/molecules25163682
Maina S, Misinzo G, Bakari G, Kim H-Y. Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules. 2020; 25(16):3682. https://doi.org/10.3390/molecules25163682
Chicago/Turabian StyleMaina, Sylvia, Gerald Misinzo, Gaymary Bakari, and Ho-Youn Kim. 2020. "Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review" Molecules 25, no. 16: 3682. https://doi.org/10.3390/molecules25163682
APA StyleMaina, S., Misinzo, G., Bakari, G., & Kim, H.-Y. (2020). Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules, 25(16), 3682. https://doi.org/10.3390/molecules25163682