
Apoferritin and Apoferritin-Capped Metal Nanoparticles Inhibit Arginine Kinase of Trypanosoma brucei


 ,
,  and
and 
Abstract
1. Introduction
2. Results
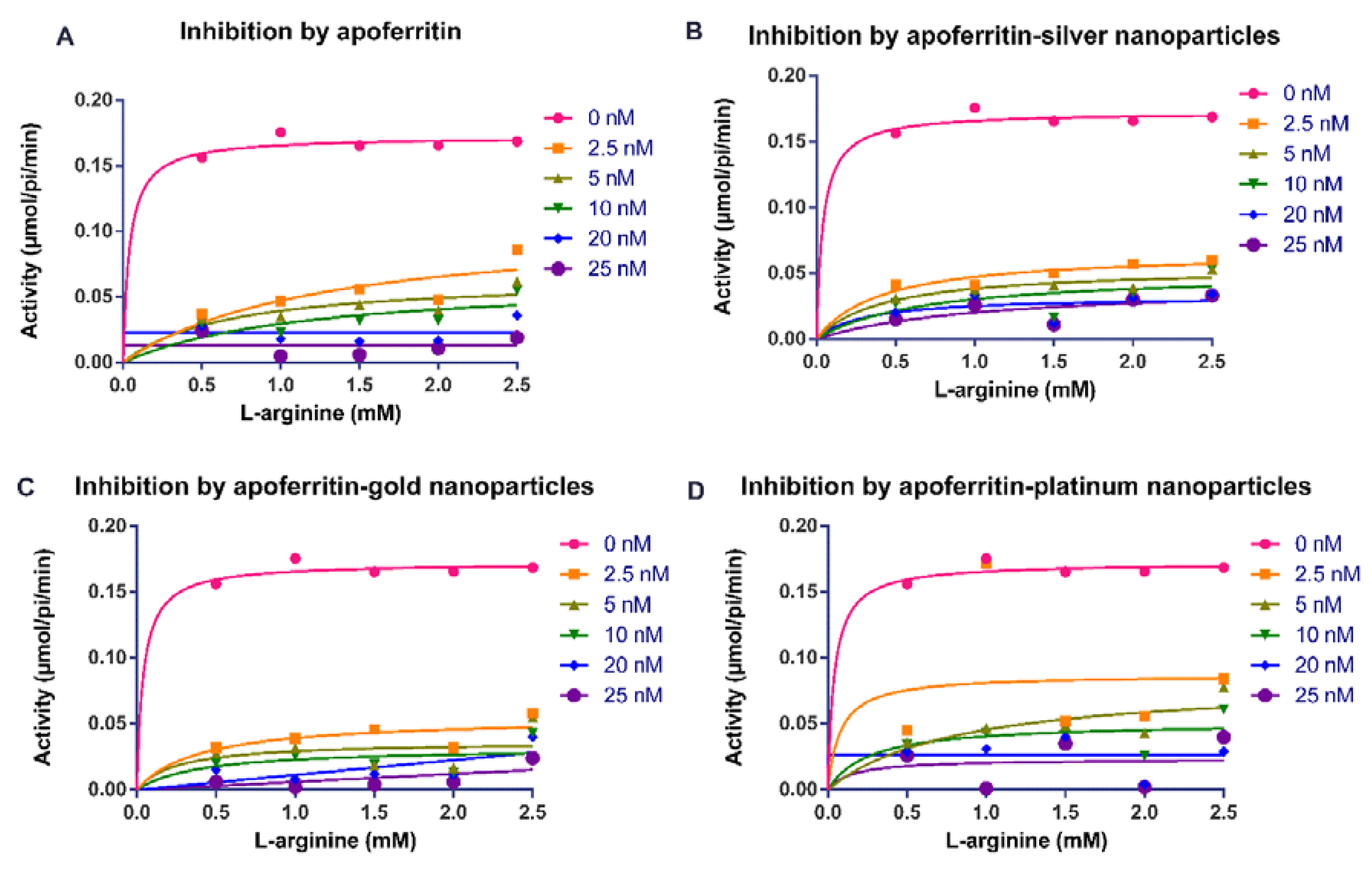
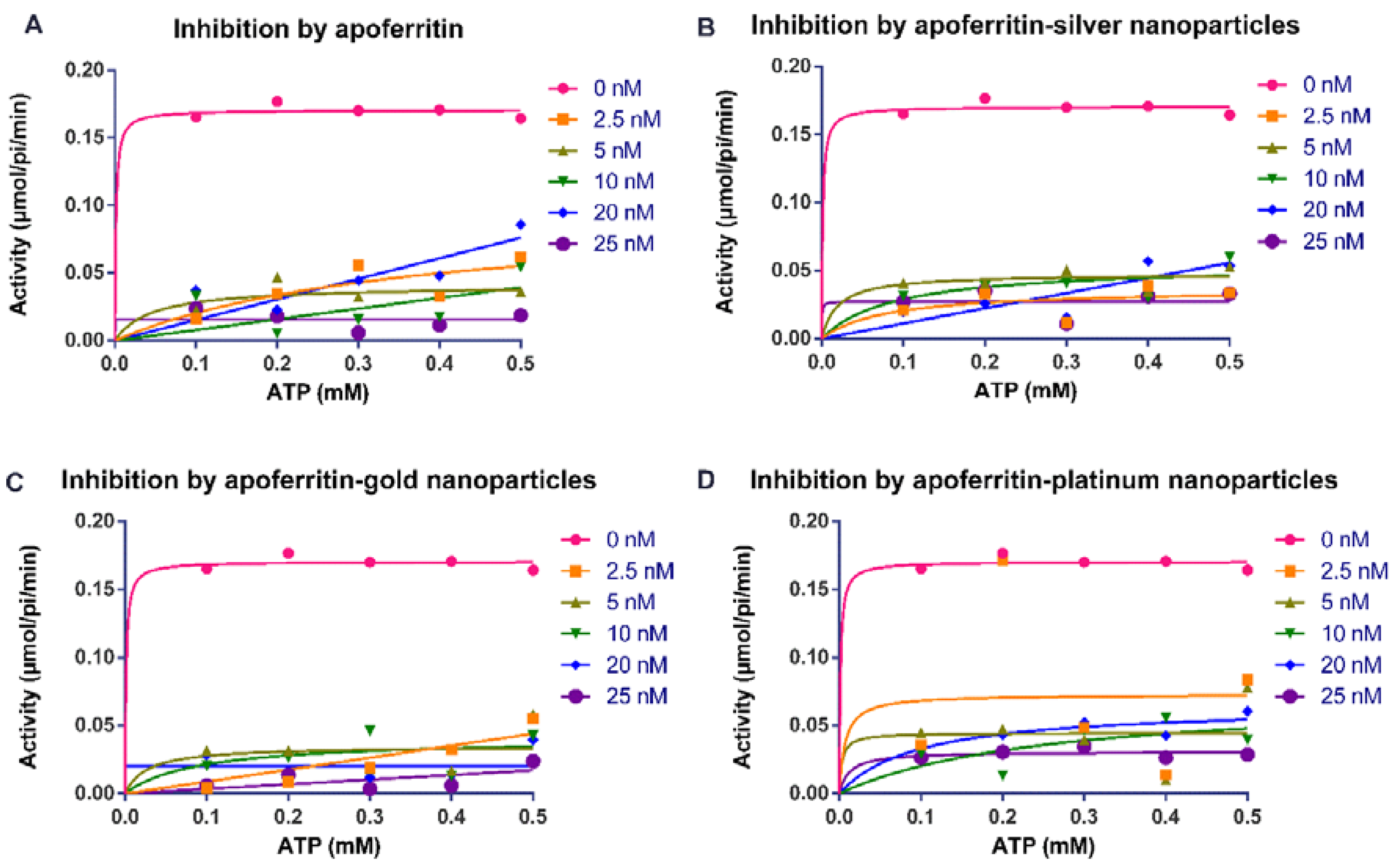
2.1. Apoferritin and Apoferritin-Capped Nanoparticles Reduced the Activity of Arginine Kinase of T. brucei and Modulated Kinetic Parameters
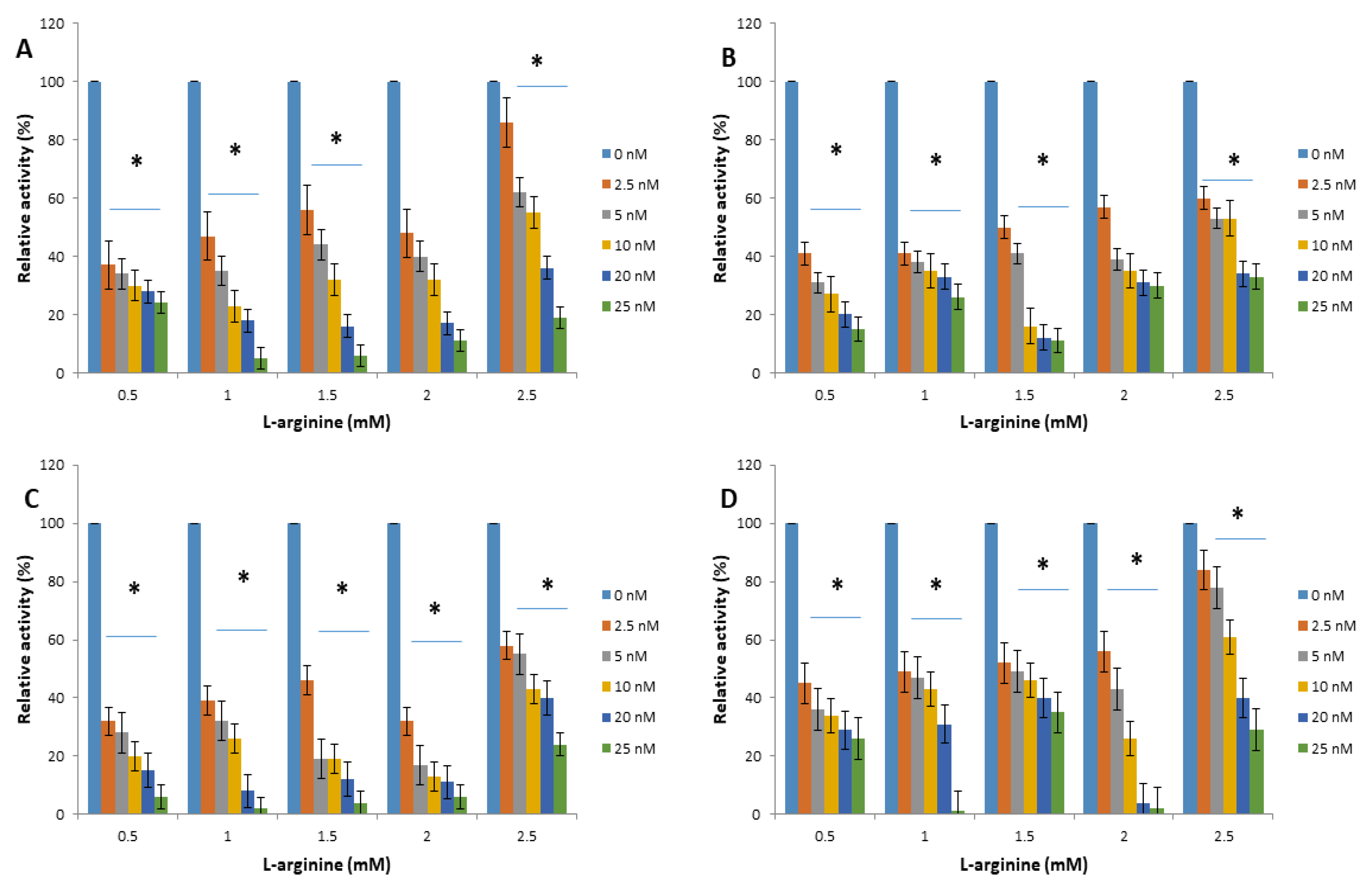
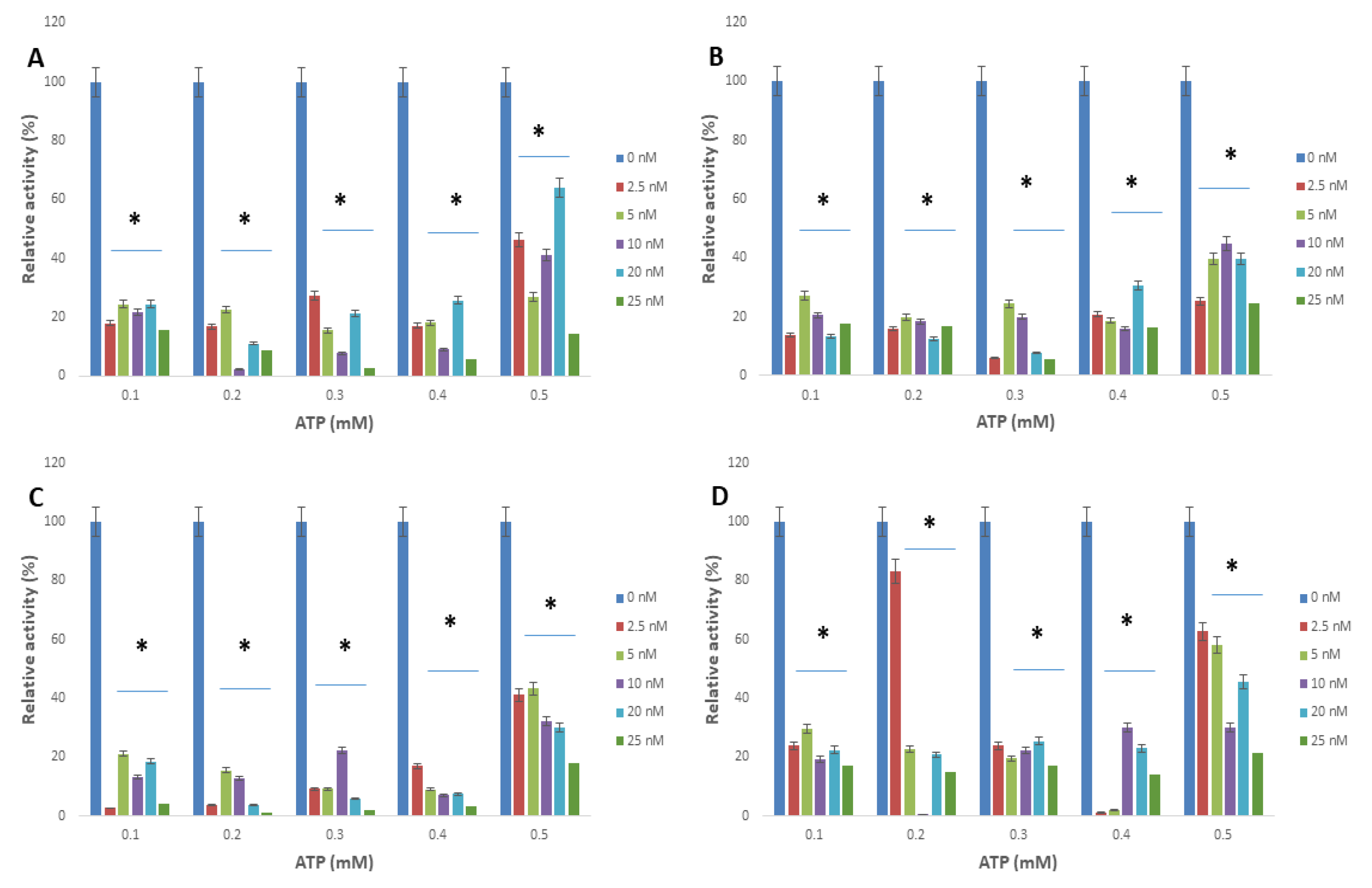
2.2. Apoferritin and Apoferritin-Capped Nanoparticles Caused Significant Decreases in the Relative Activity of Arginine Kinase of T. brucei
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Determination of Protein Concentration and Activity of the Arginine Kinase of T. brucei
4.3. Assay to Determine Arginine Kinase Activity in the Presence of Apoferritin or Apoferritin-Nanoparticles
4.4. Data Presentation and Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Trypanosomiasis, Human African (Sleeping Sickness). 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 14 May 2020).
- Jarilla, B.R.; Agatsuma, T. Phosphagen Kinases of Parasites: Unexplored Chemotherapeutic Targets. Korean J. Parasitol. 2010, 48, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Voncken, F.G.J.; Gao, F.; Wadforth, C.; Harley, M.; Colasante, C. The Phosphoarginine Energy-Buffering System of Trypanosoma brucei Involves Multiple Arginine Kinase Isoforms with Different Subcellular Locations. PLoS ONE 2013, 8, e65908. [Google Scholar] [CrossRef]
- Pereira, C.A.; Bouvier, L.A.; Cámara, M.D.L.M.; Miranda, M.R. Singular Features of Trypanosomatids’ Phosphotransferases Involved in Cell Energy Management. Enzym. Res. 2011, 2011, 1–12. [Google Scholar] [CrossRef]
- Bachmaier, S.; Santos, Y.V.; Kramer, S.; Githure, G.B.; Klöckner, T.; Pepperl, J.; Baums, C.; Schenk, R.; Schwede, F.; Genieser, H.-G.; et al. Nucleoside analogue activators of cyclic AMP-independent protein kinase A of Trypanosoma. Nat. Commun. 2019, 10, 1421. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Shah, N.; Waghela, B.; Pathak, C.; Pappachan, A. Leishmania donovani adenylate kinase 2a prevents ATP-mediated cell cytolysis in macrophages. Parasitol. Int. 2019, 72, 101929. [Google Scholar] [CrossRef]
- Thomas, S.M.; Purmal, A.; Pollastri, M.; Mensa-Wilmot, K. Discovery of a Carbazole-Derived Lead Drug for Human African Trypanosomiasis. Sci. Rep. 2016, 6, 32083. [Google Scholar] [CrossRef] [PubMed]
- Holzmuller, P.; Geiger, A.; Nzoumbou-Boko, R.; Pissarra, J.; Hamrouni, S.; Rodrigues, V.; Dauchy, F.-A.; Lemesre, J.-L.; Vincendeau, P.; Bras-Gonçalves, R. Trypanosomatid Infections: How Do Parasites and Their Excreted–Secreted Factors Modulate the Inducible Metabolism of l-Arginine in Macrophages? Front. Immunol. 2018, 9, 778. [Google Scholar] [CrossRef]
- Ooi, C.-P.; Rotureau, B.; Gribaldo, S.; Georgikou, C.; Julkowska, D.; Blisnick, T.; Perrot, S.; Subota, I.; Bastin, P. The Flagellar Arginine Kinase in Trypanosoma brucei Is Important for Infection in Tsetse Flies. PLoS ONE 2015, 10, e0133676. [Google Scholar] [CrossRef]
- Pereira, C.A. Arginine Kinase: A Potential Pharmacological Target in Trypanosomiasis. Infect. Disord. Drug Targets 2014, 14, 30–36. [Google Scholar] [CrossRef]
- Ellington, W.R. Evolution and Physiological Roles of Phosphagen Systems. Annu. Rev. Physiol. 2001, 63, 289–325. [Google Scholar] [CrossRef]
- Chen, X.; Yao, P.; Chu, X.; Hao, L.; Guo, X.; Xu, B. Isolation of arginine kinase from Apis cerana cerana and its possible involvement in response to adverse stress. Cell Stress Chap. 2014, 20, 169–183. [Google Scholar] [CrossRef]
- Fraga, D.; Aryal, M.; Hall, J.E.; Rae, E.; Snider, M.J. Characterization of the arginine kinase isoforms in Caenorhabditis elegans. Comp. Biochem. Physiol. Part. B Biochem. Mol. Boil. 2015, 187, 85–101. [Google Scholar] [CrossRef] [PubMed]
- El Kouni, M.H. Pyrimidine metabolism in schistosomes: A comparison with other parasites and the search for potential chemotherapeutic targets. Comp. Biochem. Physiol. Part. B Biochem. Mol. Boil. 2017, 213, 55–80. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.A.; Alonso, G.D.; Ivaldi, S.; Silber, A.M.; Alves, M.J.M.; Torres, H.N.; Flawiá, M.M. Arginine kinase overexpression improves Trypanosoma cruzi survival capability. FEBS Lett. 2003, 554, 201–205. [Google Scholar] [CrossRef]
- Miranda, M.R.; Canepa, G.; Bouvier, L.A.; Pereira, C.A. Trypanosoma cruzi: Oxidative stress induces arginine kinase expression. Exp. Parasitol. 2006, 114, 341–344. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Whiteley, C. Interaction of nanoparticles with arginine kinase from Trypanosoma brucei: Kinetic and mechanistic evaluation. Int. J. Boil. Macromol. 2013, 62, 450–456. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Whiteley, C. Interaction of metal nanoparticles with recombinant arginine kinase from Trypanosoma brucei: Thermodynamic and spectrofluorimetric evaluation. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 701–706. [Google Scholar] [CrossRef][Green Version]
- Adeyemi, O.S.; Sulaiman, A.F.; Iniaghe, O.M. Interaction between Gallotannin and a Recombinant Form of Arginine Kinase of Trypanosoma brucei: Thermodynamic and Spectrofluorimetric Evaluation. J. Biophys. 2014, 2014, 675905. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Molefe, N.I.; Awakan, O.J.; Nwonuma, C.O.; Alejolowo, O.; Olaolu, T.; Maimako, R.F.; Suganuma, K.; Han, Y.; Kato, K. Metal nanoparticles show potential to restrict Trypanosoma growth. Artif. Cells Nanomed. Biotechnol. 2018, 46, S86–S94. [Google Scholar] [CrossRef]
- Benelli, G. Gold nanoparticles against parasites and insect vectors. Acta Trop. 2018, 178, 73–80. [Google Scholar] [CrossRef]
- Narayanaswamy, R.; Kanagesan, S.; Pandurangan, A.; Padmanabhan, P. Basics to different imaging techniques, different nanobiomaterials for image enhancement. Nanobiomat. Med. Imag. 2016, 8, 101–129. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, H.; Zhang, Y.; Liu, G.; Niu, G.; Chen, X. Functional ferritin nanoparticles for biomedical applications. Front. Chem. Sci. Eng. 2017, 11, 633–646. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pereira, C.A.; Alonso, G.D.; Paveto, M.C.; Iribarren, A.; Cabanas, M.L.; Torres, H.N.; Flawia, M.M. Trypanosoma cruzi arginine kinase characterization and cloning. A novel energetic pathway in protozoan parasites. J. Boil. Chem. 2000, 275, 1495–1501. [Google Scholar] [CrossRef]
- Pereira, C.A.; Alonso, G.D.; Torres, H.N.; Flawia, M.M. Arginine Kinase: A Common Feature for Management of Energy Reserves in African and American Flagellated Trypanosomatids. J. Eukaryot. Microbiol. 2002, 49, 82–85. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Sulaiman, F.A. Evaluation of metal nanoparticles for drug delivery systems. J. Biomed. Res. 2015, 29, 145–149. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Han, Y.; Kato, K. Nanoparticles show potential to retard bradyzoites in vitro formation of Toxoplasma gondii. Folia Parasitol. 2019, 66, 1–6. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Han, Y.; Kato, K. Exploring Amino Acid-Capped Nanoparticles for Selective Anti-Parasitic Action and Improved Host Biocompatibility. J. Biomed. Nanotechnol. 2018, 14, 847–867. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Kato, K. Inorganic nanoparticles kill Toxoplasma gondii via changes in redox status and mitochondrial membrane potential. Int. J. Nanomed. 2017, 12, 1647–1661. [Google Scholar] [CrossRef]
- Jiao, Z.-H.; Li, M.; Feng, Y.-X.; Shi, J.-C.; Zhang, J.; Shao, B. Hormesis Effects of Silver Nanoparticles at Non-Cytotoxic Doses to Human Hepatoma Cells. PLoS ONE 2014, 9, e102564. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Faniyan, T.O. Antioxidant status of rats administered silver nanoparticles orally. J. Taibah Univ. Sci. 2014, 9, 182–186. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Adewumi, I.; Faniyan, T.O. Silver nanoparticles influenced rat serum metabolites and tissue morphology. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Olajide, I.O.; Adeyanju, A.A.; Awakan, O.J.; Otohinoyi, D.A. Modulation of rat plasma kynurenine level by platinum nanoparticles and likely association with oxidative stress. Biointerface Res. Appl. Chem. 2018, 8, 3364–3367. [Google Scholar]
- Adeyemi, O.S.; Sulaiman, F.A.; Akanji, M.A.; Oloyede, H.O.B.; Olatunde, A.; Salman, S.T.; Aransiola, A.R.; Ajayi, A.G.; Ekundayo, M.M.; Abubakar, F.A.; et al. Biochemical and morphological changes in rats exposed to platinum nanoparticles. Comp. Haematol. Int. 2016, 25, 855–864. [Google Scholar] [CrossRef]
- Sennuga, A.; Van Marwijk, J.; Whiteley, C.G. Ferroxidase activity of apoferritin is increased in the presence of platinum nanoparticles. Nanotechnol. 2011, 23, 035102. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors upon request. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| * Vmax (µmol/sec) | ** Vmax (µmol/sec) | Km (L-arginine) (mM) | Km (ATP) (mM) | * Mean Ki (nM) | ** Mean Ki (nM) | |
|---|---|---|---|---|---|---|
| Control Assay | 0.169 ± 0.004 | 0.170 ± 0.010 | 0.021 ± 0.01 | 0.005 ± 0.00 | - | - |
| Apoferritin | 0.012 ± 0.001 | 0.016 ± 0.001 | 0.041 ± 0.00 | 0.015 ± 0.00 | 0.062 ± 0.01 | 0.001 ± 0.00 |
| Apoferritin-AgNPs | 0.044 ± 0.002 | 0.034 ± 0.000 | 0.041 ± 0.01 | 0.011 ± 0.00 | 0.168 ± 0.03 | 0.057 ± 0.00 |
| Apoferritin-AuNPs | 0.067 ± 0.006 | 0.016 ± 0.001 | 0.044 ± 0.00 | 0.015 ± 0.00 | 0.160 ± 0.00 | 0.003 ± 0.00 |
| Apoferritin-PtNPs | 0.024 ± 0.007 | 0.030 ± 0.001 | 0.043 ± 0.00 | 0.001 ± 0.00 | 0.245 ± 0.03 | 0.002 ± 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeyemi, O.S.; Arowolo, A.T.; Hetta, H.F.; Al-Rejaie, S.; Rotimi, D.; Batiha, G.E.-S. Apoferritin and Apoferritin-Capped Metal Nanoparticles Inhibit Arginine Kinase of Trypanosoma brucei. Molecules 2020, 25, 3432. https://doi.org/10.3390/molecules25153432
Adeyemi OS, Arowolo AT, Hetta HF, Al-Rejaie S, Rotimi D, Batiha GE-S. Apoferritin and Apoferritin-Capped Metal Nanoparticles Inhibit Arginine Kinase of Trypanosoma brucei. Molecules. 2020; 25(15):3432. https://doi.org/10.3390/molecules25153432
Chicago/Turabian StyleAdeyemi, Oluyomi Stephen, Afolake T. Arowolo, Helal F. Hetta, Salim Al-Rejaie, Damilare Rotimi, and Gaber El-Saber Batiha. 2020. "Apoferritin and Apoferritin-Capped Metal Nanoparticles Inhibit Arginine Kinase of Trypanosoma brucei" Molecules 25, no. 15: 3432. https://doi.org/10.3390/molecules25153432
APA StyleAdeyemi, O. S., Arowolo, A. T., Hetta, H. F., Al-Rejaie, S., Rotimi, D., & Batiha, G. E.-S. (2020). Apoferritin and Apoferritin-Capped Metal Nanoparticles Inhibit Arginine Kinase of Trypanosoma brucei. Molecules, 25(15), 3432. https://doi.org/10.3390/molecules25153432