
Alvaxanthone, a Thymidylate Synthase Inhibitor with Nematocidal and Tumoricidal Activities

 ,
,  ,
,  , , , ,
, , , , 
 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Chemical Library and Virtual Screening
2.2. Predicted Properties of Compounds From the In-House Library
2.3. TS Inhibition
2.4. Toxicity to C. Elegans
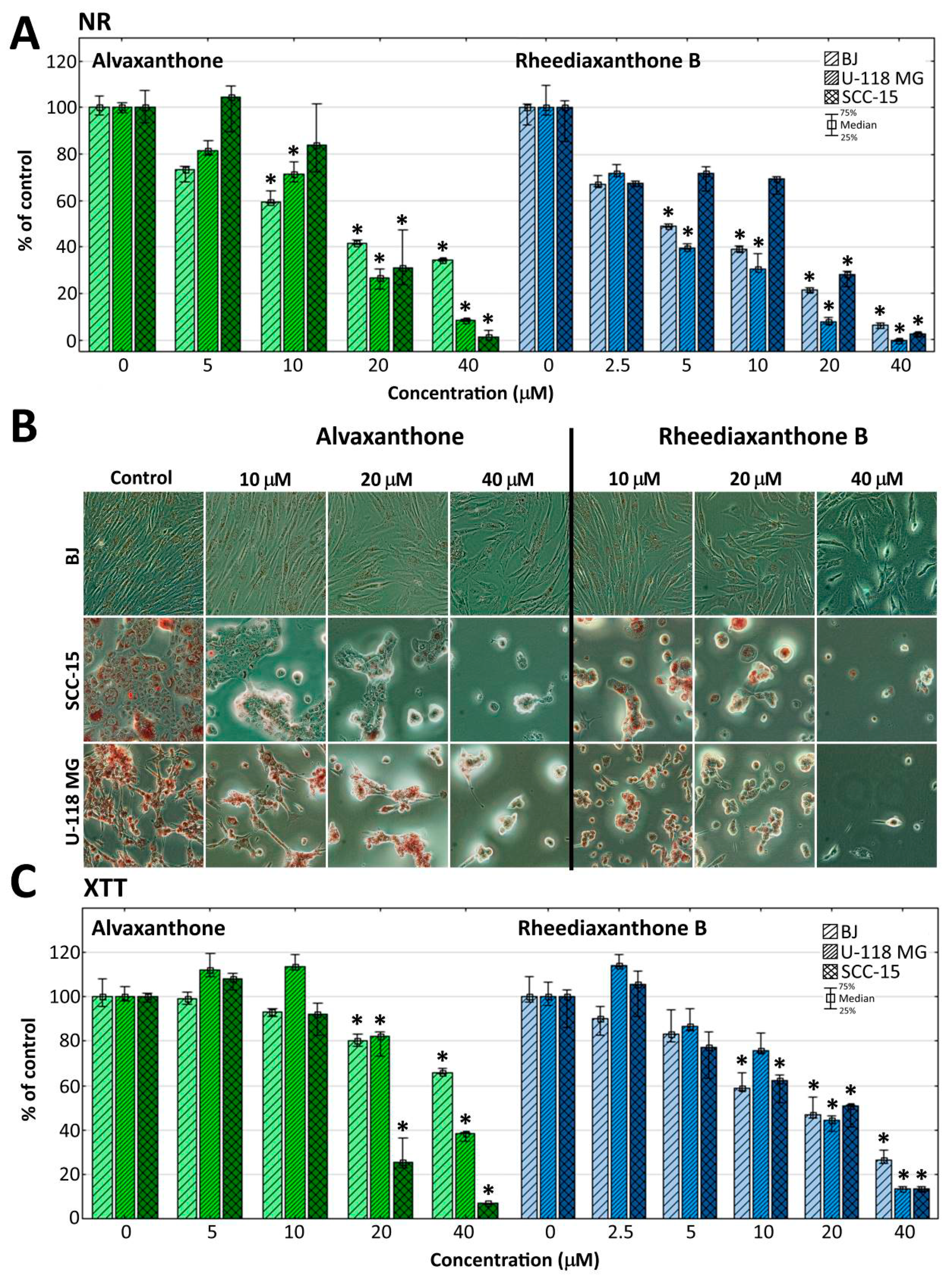
2.5. Cytotoxicity
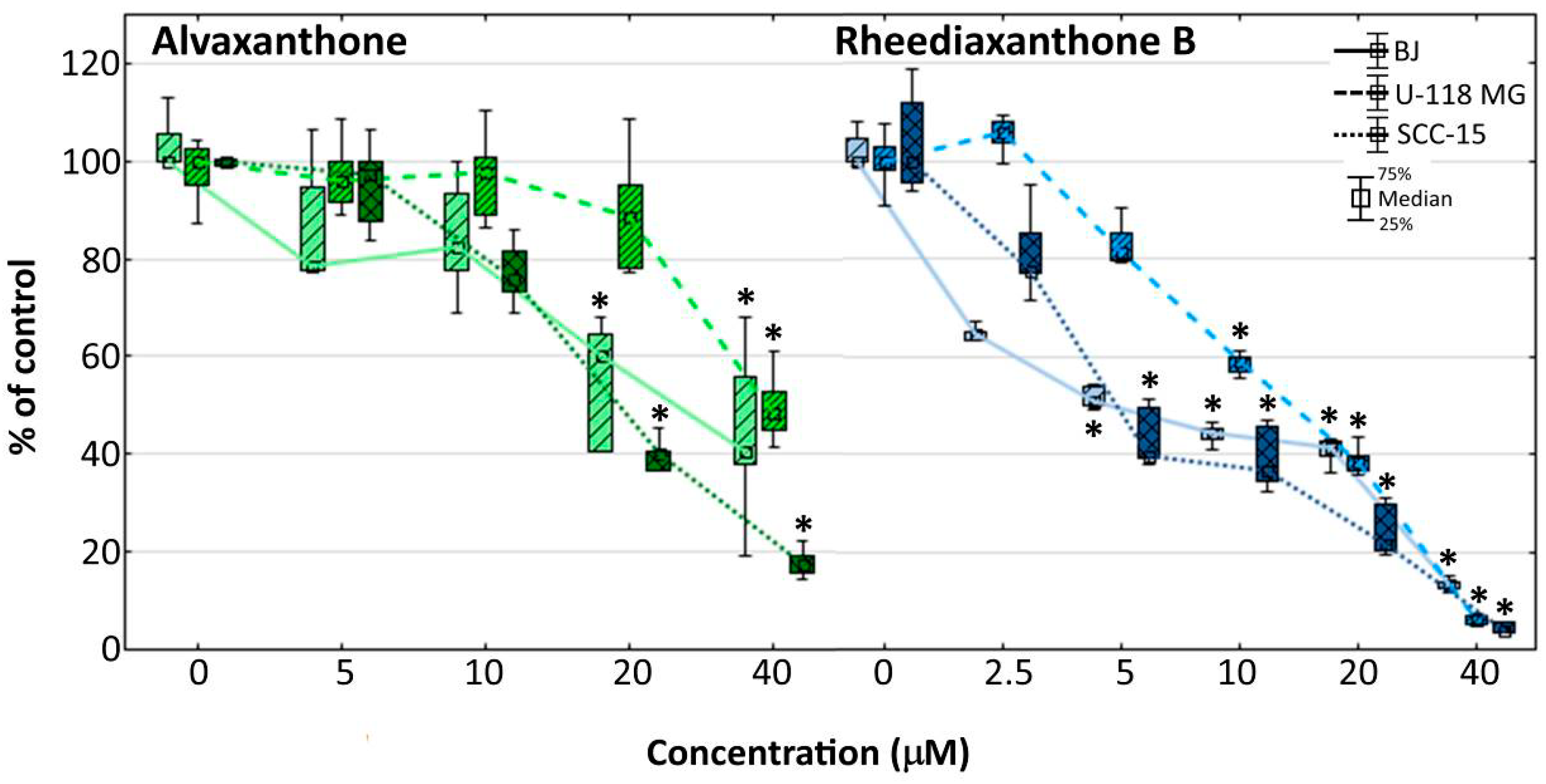
2.6. Proliferation
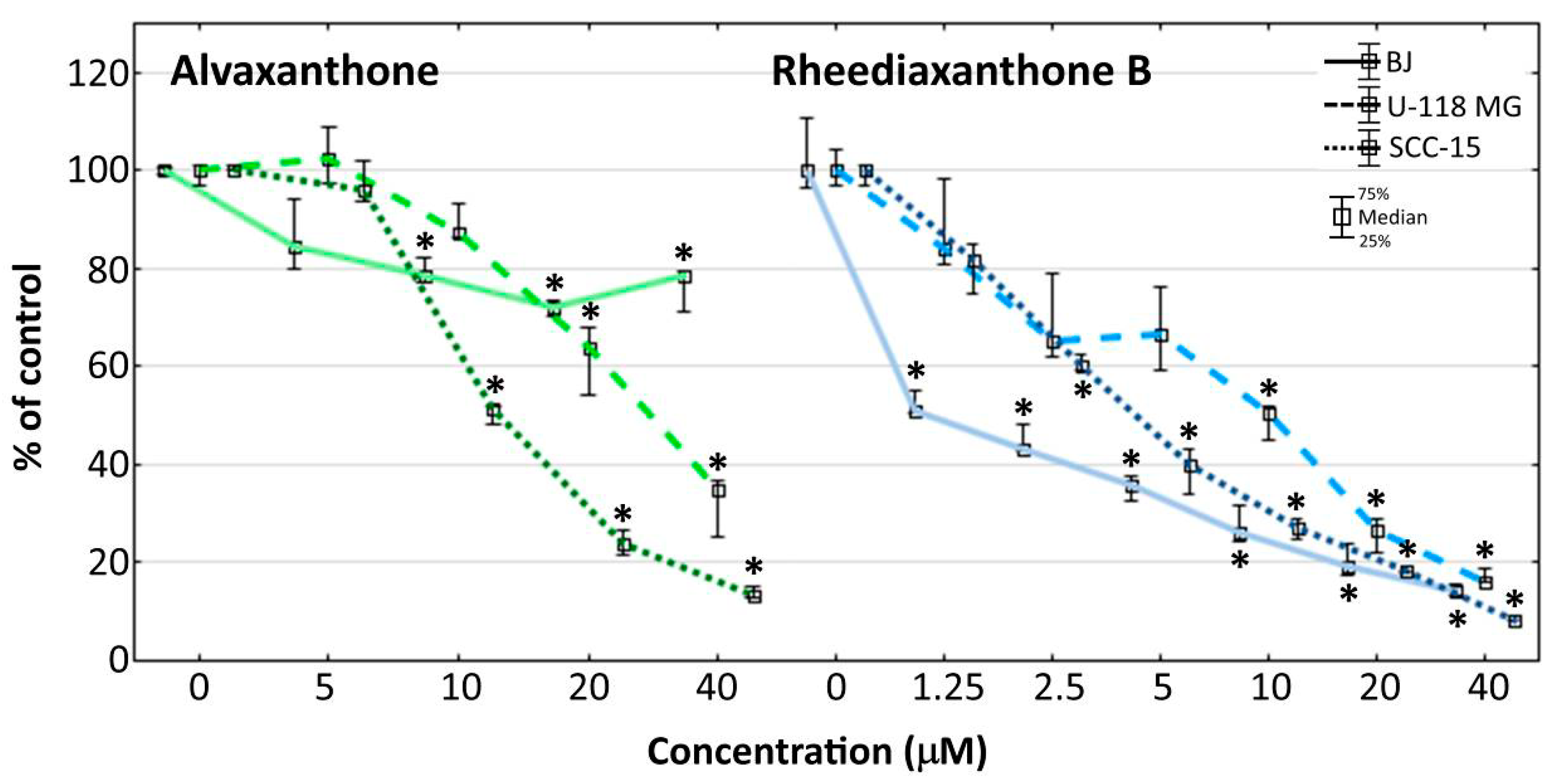
2.7. Adhesion
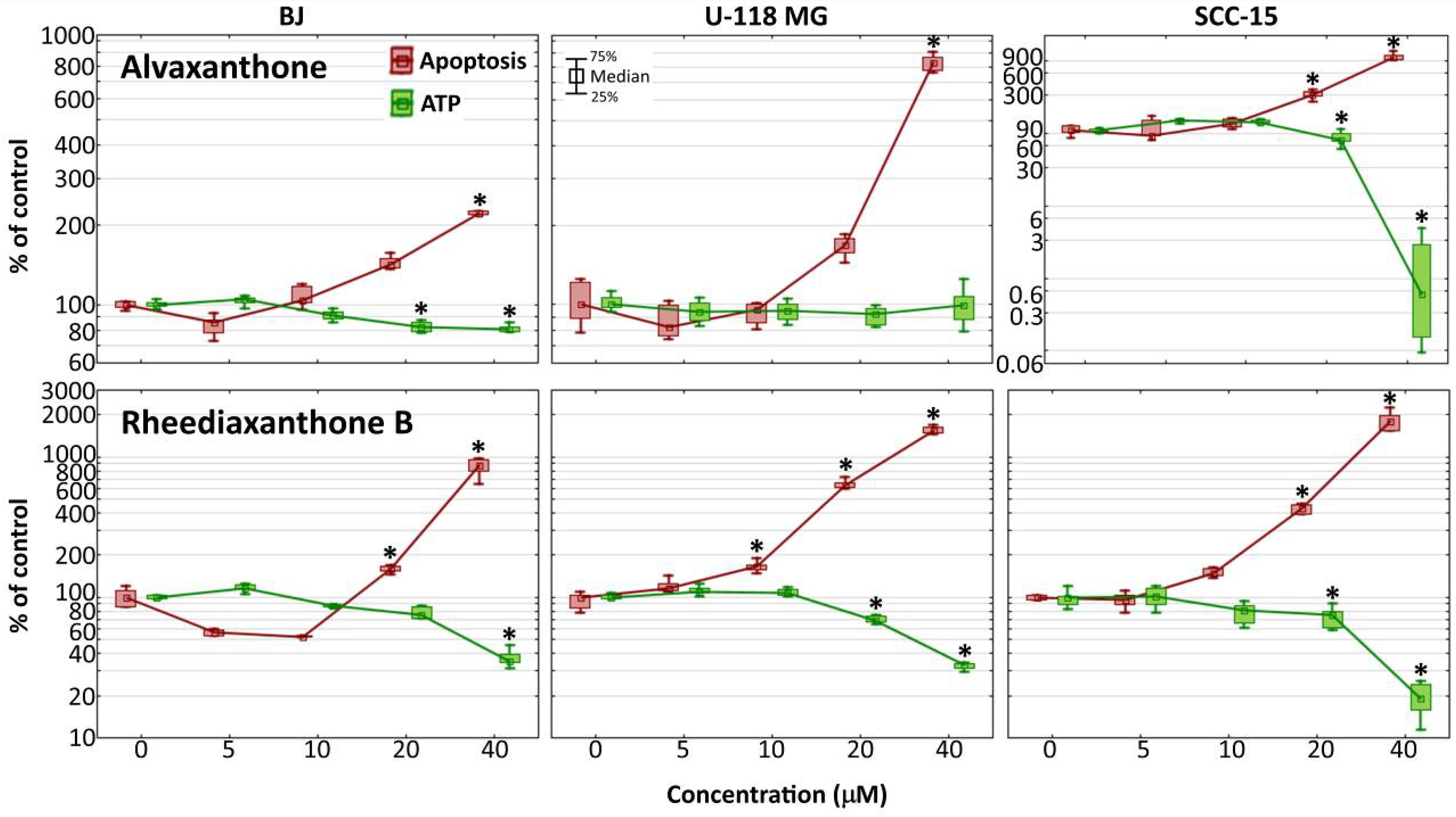
2.8. Caspase-3/7 and ATP Level
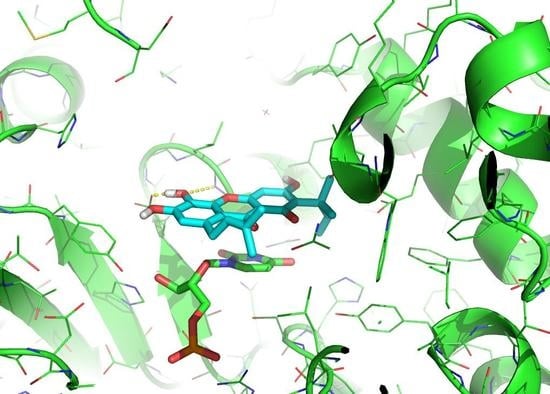
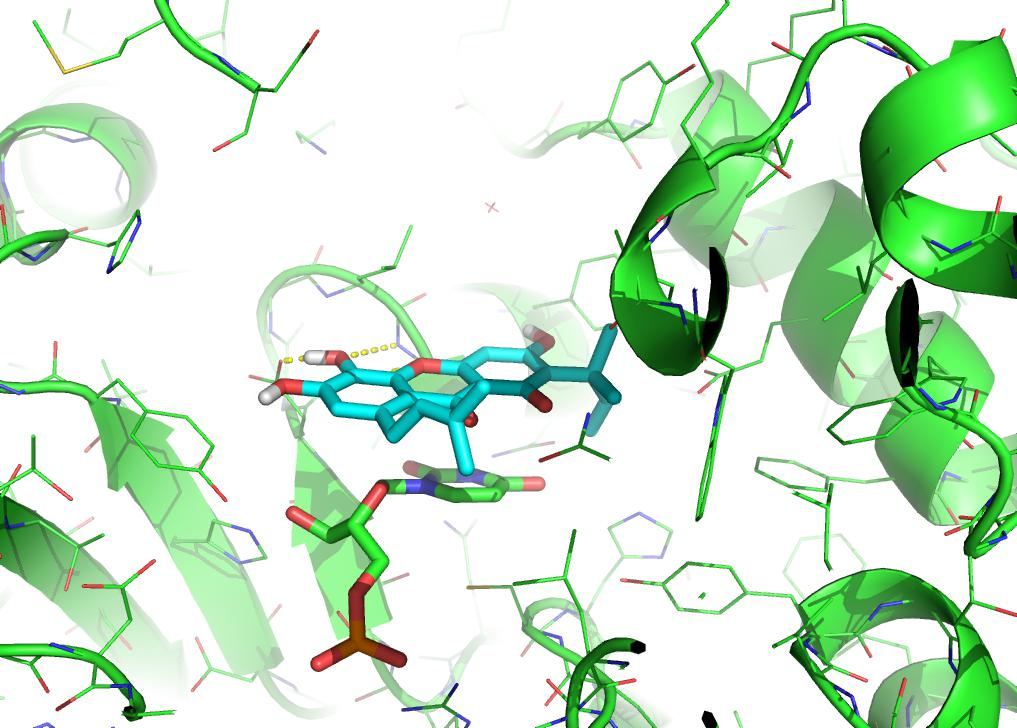
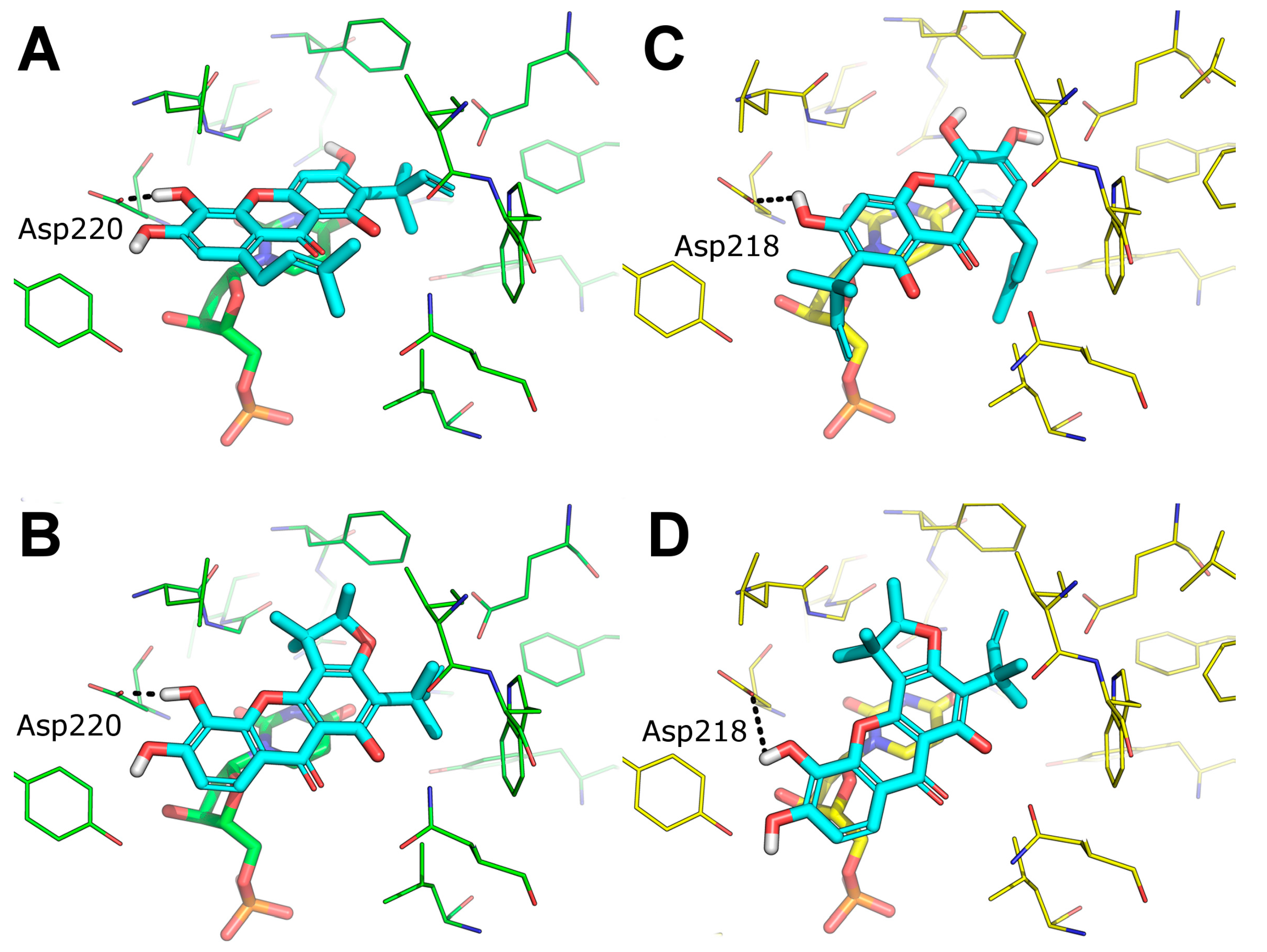
2.9. Predicted Binding Mode of Alvaxanthone and Rheediaxanthone B
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Computational Methods
4.3. Chemistry
4.4. TS preparation
4.5. Determination of Toxicity to C. elegans, a Nematode Parasite Model
4.6. Cell Culture Studies
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rode, W. Specificity of Thymidylate Synthase Inactivation by 4,5-Bisubstituted DUMP Analogues. Acta Biochim. Pol. 1993, 40, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Georgopapadakou, N.H.; Walsh, T.J. Antifungal Agents: Chemotherapeutic Targets and Immunologic Strategies. Antimicrob. Agents Chemother. 1996, 40, 279–291. [Google Scholar] [CrossRef]
- Rathod, P.K. Antimalarial Agents Directed at Thymidylate Synthase. J. Pharm. Pharmacol. 1997, 49, 65–69. [Google Scholar] [CrossRef]
- Takemura, Y.; Jackman, A.L. Folate-Based Thymidylate Synthase Inhibitors in Cancer Chemotherapy. Anti-Cancer Drugs 1997, 8, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Costi, M.P.; Ferrari, S.; Venturelli, A.; Calò, S.; Tondi, D.; Barlocco, D. Thymidylate Synthase Structure, Function and Implication in Drug Discovery. Curr. Med. Chem. 2005, 12, 2241–2258. [Google Scholar] [CrossRef]
- Gmeiner, W.H. Novel Chemical Strategies for Thymidylate Synthase Inhibition. Curr. Med. Chem. 2005, 12, 191–202. [Google Scholar] [CrossRef]
- Singh, P.; Bhardwaj, A. Mechanism of Action of Key Enzymes Associated with Cancer Propagation and Their Inhibition by Various Chemotherapeutic Agents. Mini-Rev. Med. Chem. 2008, 8, 388–398. [Google Scholar] [CrossRef]
- Jarmuła, A.; Frączyk, T.; Cieplak, P.; Rode, W. Mechanism of Influence of Phosphorylation on Serine 124 on a Decrease of Catalytic Activity of Human Thymidylate Synthase. Bioorg. Med. Chem. 2010, 18, 3361–3370. [Google Scholar] [CrossRef][Green Version]
- Taddia, L.; D’Arca, D.; Ferrari, S.; Marraccini, C.; Severi, L.; Ponterini, G.; Assaraf, Y.G.; Marverti, G.; Costi, M.P. Inside the Biochemical Pathways of Thymidylate Synthase Perturbed by Anticancer Drugs: Novel Strategies to Overcome Cancer Chemoresistance. Drug Resist. Updat. 2015, 23, 20–54. [Google Scholar] [CrossRef]
- Pacifico, P. Nematodes: Worms of the World. MLO: Med. Lab. Obs. 2001, 33, 24–31. [Google Scholar]
- Hotez, P.J.; Brindley, P.J.; Bethony, J.M.; King, C.H.; Pearce, E.J.; Jacobson, J. Helminth Infections: The Great Neglected Tropical Diseases. J. Clin. Investig. 2008, 118, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- L’Ollivier, C.; Piarroux, R. Diagnosis of Human Nematode Infections. Expert Rev. Anti-Infect. Ther. 2013, 11, 1363–1376. [Google Scholar] [CrossRef]
- Murrell, K.D.; Pozio, E. Trichinellosis: The Zoonosis That Won’t Go Quietly. Int. J. Parasitol. 2000, 30, 1339–1349. [Google Scholar] [CrossRef]
- Bürglin, T.R.; Lobos, E.; Blaxter, M.L. Caenorhabditis Elegans as a Model for Parasitic Nematodes. Int. J. Parasitol. 1998, 28, 395–411. [Google Scholar] [CrossRef]
- Burns, A.R.; Luciani, G.M.; Musso, G.; Bagg, R.; Yeo, M.; Zhang, Y.; Rajendran, L.; Glavin, J.; Hunter, R.; Redman, E.; et al. Caenorhabditis Elegans Is a Useful Model for Anthelmintic Discovery. Nat. Commun. 2015, 6, 7485. [Google Scholar] [CrossRef]
- Chaweeborisuit, P.; Suriyonplengsaeng, C.; Suphamungmee, W.; Sobhon, P.; Meemon, K. Nematicidal Effect of Plumbagin on Caenorhabditis elegans: A Model for Testing a Nematicidal Drug. Z. Naturforsch. C J. Biosci. 2016, 71, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Wińska, P.; Gołos, B.; Cieśla, J.; Zieliński, Z.; Fraczyk, T.; Wałajtys-Rode, E.; Rode, W. Developmental Arrest in Caenorhabditis elegans Dauer Larvae Causes High Expression of Enzymes Involved in Thymidylate Biosynthesis, Similar to That Found in Trichinella Muscle Larvae. Parasitology 2005, 131, 247–254. [Google Scholar] [CrossRef]
- Rode, W.; Dabrowska, M.; Zieliński, Z.; Gołos, B.; Wranicz, M.; Felczak, K.; Kulikowski, T. Trichinella spiralis and Trichinella pseudospiralis: Developmental Patterns of Enzymes Involved in Thymidylate Biosynthesis and Pyrimidine Salvage. Parasitology 2000, 120, 593–600. [Google Scholar] [CrossRef]
- Parsels, L.; Chu, E. The Role of Translational Control of the Cell Cycle. Cancer J. Sci. Am. 1998, 4, 287–295. [Google Scholar]
- Frączyk, T.; Ruman, T.; Wilk, P.; Palmowski, P.; Rogowska-Wrzesinska, A.; Cieśla, J.; Zieliński, Z.; Nizioł, J.; Jarmuła, A.; Maj, P.; et al. Properties of Phosphorylated Thymidylate Synthase. Biochim. Biophys. Acta 2015, 1854, 1922–1934. [Google Scholar] [CrossRef]
- Rahman, L.; Voeller, D.; Rahman, M.; Lipkowitz, S.; Allegra, C.; Barrett, J.C.; Kaye, F.J.; Zajac-Kaye, M. Thymidylate Synthase as an Oncogene: A Novel Role for an Essential DNA Synthesis Enzyme. Cancer Cell 2004, 5, 341–351. [Google Scholar] [CrossRef]
- Casciaro, B.; Calcaterra, A.; Cappiello, F.; Mori, M.; Loffredo, M.R.; Ghirga, F.; Mangoni, M.L.; Botta, B.; Quaglio, D. Nigritanine as a new potential antimicrobial alkaloid for the treatment of Staphylococcus aureus-induced infections. Toxins 2019, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Tottone, L.; Quaglio, D.; Zhdanovskaya, N.; Ingallina, C.; Fusto, M.; Ghirga, F.; Peruzzi, G.; Crestoni, M.E.; Simeoni, F.; et al. Identification of a novel chalcone derivative that inhibits Notch signaling in T-cell acute lymphoblastic leukemia. Sci. Rep. 2017, 7, 2213. [Google Scholar] [CrossRef] [PubMed]
- Infante, P.; Alfonsi, R.; Ingallina, C.; Quaglio, D.; Ghirga, F.; D’Acquarica, I.; Bernardi, F.; Di Magno, L.; Canettieri, G.; Screpanti, I.; et al. Inhibition of Hedgehog-dependent tumors and cancer stem cells by a newly identified naturally occurring chemotype. Cell Death Dis. 2016, 7, e2376. [Google Scholar] [CrossRef] [PubMed]
- Infante, P.; Mori, M.; Alfonsi, R.; Ghirga, F.; Aiello, F.; Toscano, S.; Ingallina, C.; Siler, M.; Cucchi, D.; Po, A.; et al. Gli1/DNA interaction is a druggable target for Hedgehog-dependent tumors. EMBO J. 2015, 34, 200–217. [Google Scholar] [CrossRef]
- Cevatemre, B.; Erkisa, M.; Aztopal, N.; Karakas, D.; Alper, P.; Tsimplouli, C.; Sereti, E.; Dimas, K.; Armutak, E.I.I.; Gurevin, E.G.G.; et al. A promising natural product, pristimerin, results in cytotoxicity against breast cancer stem cells in vitro and xenografts in vivo through apoptosis and an incomplete autopaghy in breast cancer. Pharmacol. Res. 2018, 129, 500–514. [Google Scholar] [CrossRef]
- Cevatemre, B.; Botta, B.; Mori, M.; Berardozzi, S.; Ingallina, C.; Ulukaya, E. The plant-derived triterpenoid tingenin B is a potent anticancer agent due to its cytotoxic activity on cancer stem cells of breast cancer in vitro. Chem. Biol. Interact. 2016, 260, 248–255. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular Properties That Influence the Oral Bioavailability of Drug Candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Lobell, M.; Hendrix, M.; Hinzen, B.; Keldenich, J.; Meier, H.; Schmeck, C.; Schohe-Loop, R.; Wunberg, T.; Hillisch, A. In Silico ADMET Traffic Lights as a Tool for the Prioritization of HTS Hits. ChemMedChem 2006, 1, 1229–1236. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A BOILED-Egg to Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Egan, W.J.; Merz, K.M.; Baldwin, J.J. Prediction of Drug Absorption Using Multivariate Statistics. J. Med. Chem. 2000, 43, 3867–3877. [Google Scholar] [CrossRef] [PubMed]
- Muegge, I.; Heald, S.L.; Brittelli, D. Simple Selection Criteria for Drug-like Chemical Matter. J. Med. Chem. 2001, 44, 1841–1846. [Google Scholar] [CrossRef]
- Baell, J.B.; Holloway, G.A. New Substructure Filters for Removal of Pan Assay Interference Compounds (PAINS) from Screening Libraries and for Their Exclusion in Bioassays. J. Med. Chem. 2010, 53, 2719–2740. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Duan, D.; Torosyan, H.; Doak, A.K.; Ziebart, K.T.; Sterling, T.; Tumanian, G.; Shoichet, B.K. An Aggregation Advisor for Ligand Discovery. J. Med. Chem. 2015, 58, 7076–7087. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
- Shoichet, B.K. Screening in a Spirit Haunted World. Drug Discov. Today 2006, 11, 607–615. [Google Scholar] [CrossRef]
- Morrison, J.F. The slow-binding and slow, tight-binding inhibition of enzyme-catalysed reactions. Trends Biochem. Sci. 1982, 7, 102–105. [Google Scholar] [CrossRef]
- Markowicz, J.; Uram, Ł.; Sobich, J.; Mangiardi, L.; Maj, P.; Rode, W. Antitumor and Anti-Nematode Activities of α-Mangostin. Eur. J. Pharmacol. 2019, 863, 172678. [Google Scholar] [CrossRef]
- Rozanov, D.; Cheltsov, A.; Nilsen, A.; Boniface, C.; Forquer, I.; Korkola, J.; Gray, J.; Tyner, J.; Tognon, C.E.; Mills, G.B.; et al. Targeting Mitochondria in Cancer Therapy Could Provide a Basis for the Selective Anti-Cancer Activity. PLoS ONE 2019, 14, e0205623. [Google Scholar] [CrossRef]
- Hou, A.-J.; Fukai, T.; Shimazaki, M.; Sakagami, H.; Sun, H.-D.; Nomura, T. Benzophenones and Xanthones with Isoprenoid Groups from Cudrania Cochinchinensis. J. Nat. Prod. 2001, 64, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Romar, G.A.; Kupper, T.S.; Divito, S.J. Research Techniques Made Simple: Techniques to Assess Cell Proliferation. J. Investig. Dermatol. 2016, 136, e1–e7. [Google Scholar] [CrossRef]
- Abduljauwad, S.N.; Ahmed, H.-R. Enhancing Cancer Cell Adhesion with Clay Nanoparticles for Countering Metastasis. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, N.J.; Goldstein, J.C.; von Ahsen, O.; Schuler, M.; Newmeyer, D.D.; Green, D.R. Cytochrome c Maintains Mitochondrial Transmembrane Potential and ATP Generation after Outer Mitochondrial Membrane Permeabilization during the Apoptotic Process. J. Cell Biol. 2001, 153, 319–328. [Google Scholar] [CrossRef]
- Pradelli, L.A.; Villa, E.; Zunino, B.; Marchetti, S.; Ricci, J.-E. Glucose Metabolism Is Inhibited by Caspases upon the Induction of Apoptosis. Cell Death Dis. 2014, 5, e1406. [Google Scholar] [CrossRef]
- Oropesa, M.; de la Mata, M.; Maraver, J.G.; Cordero, M.D.; Cotán, D.; Rodríguez-Hernández, A.; Domínguez-Moñino, I.; de Miguel, M.; Navas, P.; Sánchez-Alcázar, J.A. Apoptotic Microtubule Network Organization and Maintenance Depend on High Cellular ATP Levels and Energized Mitochondria. Apoptosis 2011, 16, 404–424. [Google Scholar] [CrossRef]
- Jarmuła, A.; Wilk, P.; Maj, P.; Ludwiczak, J.; Dowierciał, A.; Banaszak, K.; Rypniewski, W.; Cieśla, J.; Dąbrowska, M.; Frączyk, T.; et al. Crystal Structures of Nematode (Parasitic T. spiralis and Free Living C. elegans), Compared to Mammalian, Thymidylate Synthases (TS). Molecular Docking and Molecular Dynamics Simulations in Search for Nematode-Specific Inhibitors of TS. J. Mol. Graph. Model. 2017, 77, 33–50. [Google Scholar] [CrossRef]
- McKellar, Q.A.; Scott, E.W. The Benzimidazole Anthelmintic Agents-a Review. J. Vet. Pharmacol. Ther. 1990, 13, 223–247. [Google Scholar] [CrossRef]
- Simpkin, K.G.; Coles, G.C. The Use of Caenorhabditis elegans for Anthelmintic Screening. J. Chem. Technol. Biotechnol. 1981, 31, 66–69. [Google Scholar] [CrossRef]
- Fukai, T.; Oku, Y.; Hou, A.J.; Yonekawa, M.; Terada, S. Antimicrobial activity of hydrophobic xanthones from Cudrania cochinchinensis against Bacillus subtilis and methicillin-resistant Staphylococcus aureus. Chem. Biodivers. 2004, 1, 1385–1390. [Google Scholar] [CrossRef] [PubMed]
- Fukai, T.; Oku, Y.; Hou, A.J.; Yonekawa, M.; Terada, S. Antimicrobial Activity of Isoprenoid-Substituted Xanthones from Cudrania Cochinchinensis against Vancomycin-Resistant Enterococci. Phytomedicine 2005, 12, 510–513. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Hou, A.-J.; Zhu, G.-F.; Chen, D.-F.; Sun, H.-D. Cytotoxic and Antifungal Isoprenylated Xanthones and Flavonoids from Cudrania Fruticosa. Planta Med. 2005, 71, 273–274. [Google Scholar] [CrossRef] [PubMed]
- McGann, M. FRED Pose Prediction and Virtual Screening Accuracy. J. Chem. Inf. Model. 2011, 51, 578–596. [Google Scholar] [CrossRef] [PubMed]
- FRED 3.0.1 OpenEye Scientific Software, Santa Fe, NM. Available online: http://www.eyesopen.com (accessed on 21 June 2020).
- OMEGA 3.0.1.2: OpenEye Scientific Software, Santa Fe, NM. Available online: http://www.eyesopen.com (accessed on 21 June 2020).
- Hawkins, P.C.D.; Skillman, A.G.; Warren, G.L.; Ellingson, B.A.; Stahl, M.T. Conformer Generation with OMEGA: Algorithm and Validation Using High Quality Structures from the Protein Databank and Cambridge Structural Database. J. Chem. Inf. Model. 2010, 50, 572–584. [Google Scholar] [CrossRef]
- Chen, D.; Jansson, A.; Sim, D.; Larsson, A.; Nordlund, P. Structural analyses of human thymidylate synthase reveal a site that may control conformational switching between active and inactive states. J. Biol. Chem. 2017, 292, 13449–13458. [Google Scholar] [CrossRef]
- Lagorce, D.; Bouslama, L.; Becot, J.; Miteva, M.A.; Villoutreix, B.O. FAF-Drugs4: Free ADME-Tox Filtering Computations for Chemical Biology and Early Stages Drug Discovery. Bioinformatics 2017, 33, 3658–3660. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Kim, C.-K.; Ebizuka, Y.; Sankawa, U. Two new isoflavonoids from Echinosophora koreensis Nakai and the structure revision of sophoronol. Chem. Pharm. Bull. 1989, 37, 2879–2881. [Google Scholar] [CrossRef]
- Delle Monache, F.; Delle Monache, G.; Marini-Bettolo, G.B.; Albuquerque, M.F.D.; Mello, F.D.; Lima, O.G.D. Flavonoids of Sophora tomentosa (Leguminosae). II. Isosophoranone, a new diprenylated isoflavanone. Gazz. Chim. Ital. 1977, 107, 189–192. [Google Scholar] [CrossRef]
- Delle Monache, F.; Delle Monache, G.; Marini-Bettolo, G.B.; Albuquerque, M.F.D.; Mello, F.D.; Lima, O.G.D. Flavonoids of Sophora tomentosa (Leguminosae). I. sophoronol, a new 3-hydroxyflavanone. Gazz. Chim. Ital. 1976, 106, 935–945. [Google Scholar]
- Khan, I.U.; Ansari, W.H. flavonol glycosides from Callitris Glauca. Phytochemistry 1987, 26, 1221–1222. [Google Scholar] [CrossRef]
- Brum, R.L.; Cavalheiro, A.J.; Delle Monache, F.; Vencato, I. Jatrowediol, a Lathyrane Diterpene from Jatropha weddelliana. J. Braz. Chem. Soc. 2001, 12, 259–262. [Google Scholar] [CrossRef]
- Delle Monache, F.; De Lima, O.G.; De Mello, J.F.; Delle Monache, G.; Marini Bettolo, G.B. Chroman- and cromene-chalcones from Cordoin and Isocordoin. Gazz. Chim. Ital. 1973, 103, 779–784. [Google Scholar]
- Asakawa, Y. Chemical Constituents of Alnus sieboldiana (BETULACEAE) II. The Isolation and Structure of Flavonoids and Stilbenes. Bull. Chem. Soc. Jpn. 1971, 44, 2761–2766. [Google Scholar] [CrossRef]
- Akcos, Y.; Ezer, N.; Çalis, I.; Demirdamar, R.; Tel, B.C. Polyphenolic Compounds of Sideritis lycia and Their Anti-Inflammatory Activity. Pharm. Biol. 1999, 37, 118–122. [Google Scholar] [CrossRef]
- Fukai, T.; Wang, Q.H.; Inami, R.; Nomura, T. Structures of Prenylated Dihydrochalcone, Gancaonin J and Homoisoflavonone, Gancaonin K from Glycyrrhiza pallidiflora. Heterocycles 1990, 31, 643–650. [Google Scholar] [CrossRef]
- Ágnes, K. Természetes eredetű, potenciálisan biológiailag aktív O- és Cprenilezett flavanonok szintézise. Ph.D. Thesis, University of Debrecen, Debrecen, Hungary, Debrecen, 2007. Available online: https://adoc.tips/termeszetes-eredet-potencialisan-biologiailag-aktiv-o-es-c-p.html (accessed on 21 June 2020).
- Wolfrom, M.L.; Komitsky, F.; Mundell, P.M. Osage Orange Pigments. XVI. The Structure of Alvaxanthone. J. Org. Chem. 1965, 30, 1088–1091. [Google Scholar] [CrossRef]
- Morales Moisela, M.M.; Brel, O.; Bourdy, G.; de La Cruz, M.G.; Castro Dionicio, I.Y.; Carraz, M.; Jullian, V. Xanthones from Hypericum laricifolium Juss., and their antiproliferative activity against HEP3B cells. Rev. Soc. Quím. Perú 2018, 84, 428–435. [Google Scholar]
- Delle Monache, F.; Marquina, M.; Delle Monache, G.; Marini Bettolo, G.B.; Alves De lima, R. Xanthones, xanthonolignoids and other constituents of the roots of Vismia guaramirangae. Phytochemistry 1983, 22, 227–232. [Google Scholar] [CrossRef]
- Delle Monache, F.; Botta, B.; Nicoletti, M.; de Barros Coêlho, J.S.; de Andrade Lyra, F.D. Three new xanthones and macluraxanthone from Rheedia benthamiana PI. Triana (Guttiferae). J. Chem. Soc. Perkin Trans. 1981, 1, 484–488. [Google Scholar] [CrossRef]
- Rui, D.; Chen, X.; Li, Z.; Tang, L. Chemical Constituents of Hypericum petiolulatum. Chem. Nat. Compd. 2017, 53, 457–462. [Google Scholar] [CrossRef]
- Rode, W.; Kulikowski, T.; Kedzierska, B.; Shugar, D. Studies on the Interaction with Thymidylate Synthase of Analogues of 2′-Deoxyuridine-5′-Phosphate and 5-Fluoro-2′-Deoxyuridine-5′-Phosphate with Modified Phosphate Groups. Biochem. Pharmacol. 1987, 36, 203–210. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds might be available upon request. the compounds …… are available from. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound (No.) | Structure | IC50 [µM] Assessment (% Remaining Activity) a | |||
|---|---|---|---|---|---|
| mTS | hTS | CeTS | TsTS | ||
| Curcumin (1) |  | >50(83) <500(37) | >50(86) <500(20) | >50(84) <500(30) | >500(73) |
| Aloin (2) |  | >1000(95) | >100(94) <1000(49) | >100(83) <1000(36) | >1000(93) |
| Chlorogenic acid (3) |  | >1000(83) | >1000(76) | >1000(82) | >1000(94) |
| Phloretin (4) |  | >100(78) <1000(45) | >100(86) <1000(31) | >100(82) <1000(48) | >1000(61) |
| Sophoronol (5) |  | >100(75) <1000(10) | >100(75) <1000(8) | >100(77) <1000(8) | >100(74) <1000(35) |
| Bixin (6) |  | >667(73) | (667)NI b | >67(93) <667(46) | >67(76) <667(47) |
| Myricetin-4′-OAc (7) |  | >100(61) <1000(49) | >100(56) >1000(31) | >100(69) <1000(28) | >1000(66) |
| Agnuside (8) |  | >1000(72) | >100(90) <1000(47) | >100(100) <1000(36) | >1000(63) |
| Clusiacitran A (9) |  | >1000(62) | >100(89) <1000(47) | >100(88) <1000(44) | >1000(95) |
| Jatrowediol (10) |  | >1000(80) | >100(100) <1000(43) | >1000(77) | >1000(95) |
| 6-Chromane-O-7-flavanone (11) |  | >100(91) <1000(10) | >10(100) <100(37) | >10(100) <100(24) | >1000(85) |
| Alnusin (12) |  | (1000)NI b | >100(85) <1000(30) | >1000(80) | (1000)NI b |
| Leucosceptoside A (13) |  | >1000(77) | >1000(66) | >1000(83) | (1000)NI b |
| (±)-7-Hydro-xy-8-C-prenyl flavanone (Ovaliflavanone B) (14) |  | >100(71) <1000(2) | >100(74) <1000(7) | >100(65) <1000(3) | >100(90) <1000(25) |
| Alvaxanthone (15) |  | >10(79) <100(40) | <10(39) | >10(85) <100(18) | >10(98) <100(47) >10 †(97) <100 †(14) |
| 1,3-OH-2-OMe-Xanthone (16) |  | >100(67) <1000(32) | >100(54) <1000(27) | >1000(65) | >100(92) <1000(23) >100 †(82) <1000 †(32) |
| Rheediaxanthone B (17) |  | >10 †(56) <100 †(2) | >10 †(59 <100 †(5) | >10 †(59) <100 †(14) | >10 †(68) <100 †(40) |
| Rheediaxanthone C (18) |  | >10 †(77) <100 †(39) | >100 †(57) | >100 †(71) | >100 †(73) |
| 1-OH-7-OMe-Xanthone (19) |  | >1000 † (94) | >1000 † (100) | >1000 † (81) | >1000 † (89) |
| 2-OMe-Xanthone (20) |  | >1000 † (84) | >1000 † (88) | >1000 † (95) | >1000 † (92) |
| α-Mangostin (21) |  | >10 †(75) <100 †(5) | >10 †(89) <100 †(19) | >10 †(79) <100 †(27) | >100 †(85) |
| Compound (No.) | IC50 (μM) a | ||
|---|---|---|---|
| BJ | U-118 MG | SCC-15 | |
| Alvaxanthone (15) | 15.76 | 13.07 | 17.66 |
| 1,3-OH-2-OMe-xanthone (16) | >40 | >40 | >40 |
| Rheediaxanthone B (17) | 5.5 | 4.75 | 9.2 |
| Rheediaxanthone C (18) | 5.95 | 18.27 | 11.04 |
| 1-OH-7-OMe-Xanthone (19) | >40 | 40 | >40 |
| 2-OMe-Xanthone (20) | >40 | >40 | >40 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maj, P.; Mori, M.; Sobich, J.; Markowicz, J.; Uram, Ł.; Zieliński, Z.; Quaglio, D.; Calcaterra, A.; Cau, Y.; Botta, B.; et al. Alvaxanthone, a Thymidylate Synthase Inhibitor with Nematocidal and Tumoricidal Activities. Molecules 2020, 25, 2894. https://doi.org/10.3390/molecules25122894
Maj P, Mori M, Sobich J, Markowicz J, Uram Ł, Zieliński Z, Quaglio D, Calcaterra A, Cau Y, Botta B, et al. Alvaxanthone, a Thymidylate Synthase Inhibitor with Nematocidal and Tumoricidal Activities. Molecules. 2020; 25(12):2894. https://doi.org/10.3390/molecules25122894
Chicago/Turabian StyleMaj, Piotr, Mattia Mori, Justyna Sobich, Joanna Markowicz, Łukasz Uram, Zbigniew Zieliński, Deborah Quaglio, Andrea Calcaterra, Ylenia Cau, Bruno Botta, and et al. 2020. "Alvaxanthone, a Thymidylate Synthase Inhibitor with Nematocidal and Tumoricidal Activities" Molecules 25, no. 12: 2894. https://doi.org/10.3390/molecules25122894
APA StyleMaj, P., Mori, M., Sobich, J., Markowicz, J., Uram, Ł., Zieliński, Z., Quaglio, D., Calcaterra, A., Cau, Y., Botta, B., & Rode, W. (2020). Alvaxanthone, a Thymidylate Synthase Inhibitor with Nematocidal and Tumoricidal Activities. Molecules, 25(12), 2894. https://doi.org/10.3390/molecules25122894