
Potential Plasticity of the Mannoprotein Repertoire Associated to Mycobacterium tuberculosis Virulence Unveiled by Mass Spectrometry-Based Glycoproteomics


Abstract
1. Introduction
2. Results
2.1. Mtb Culture Filtrate Proteome Coverage Using SDS–PAGE Separation and MS-Based Shotgun Proteomics
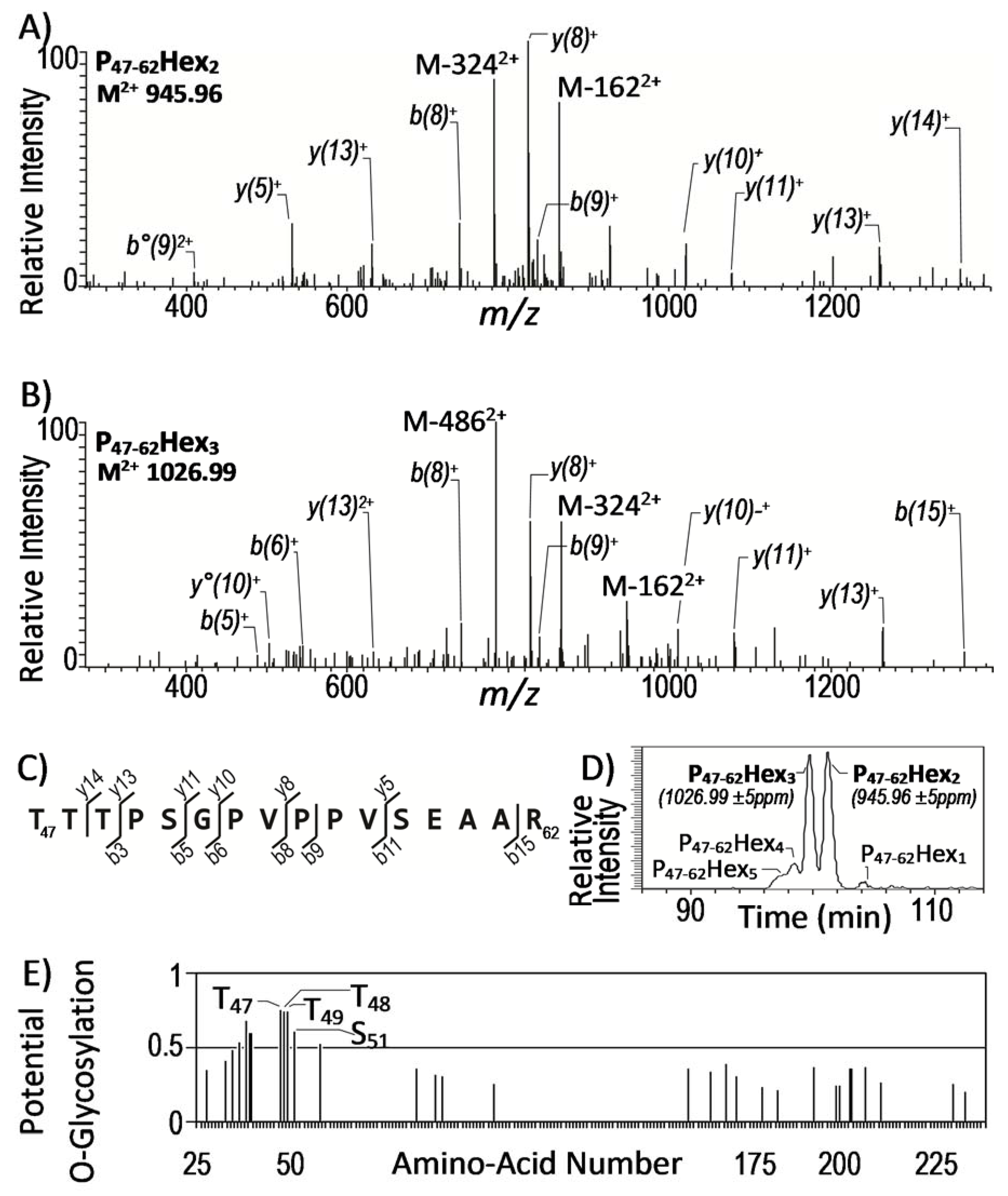
2.2. Identification of Glycopeptides Present in the Culture Filtrate of Mtb
2.3. Improvement of MS2 Data Processing for Mtb Glycoproteins Discovery
2.4. High Confidence Identification of Mtb Glycosylated Proteins
2.5. Assessment of the PMTub Associated Mannoprotein Repertoire
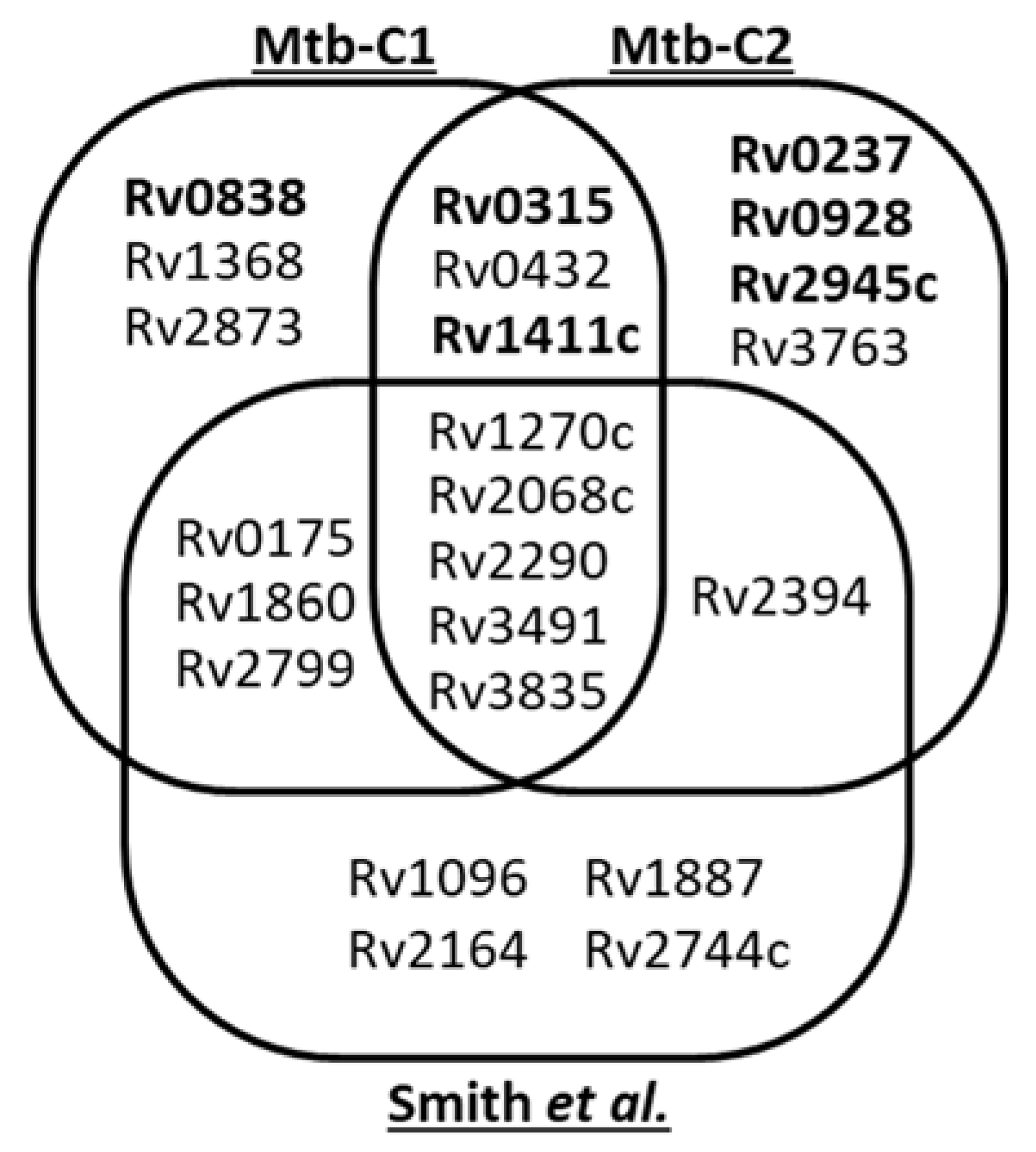
2.6. Variability of the Mtb Mannoproteome Determination
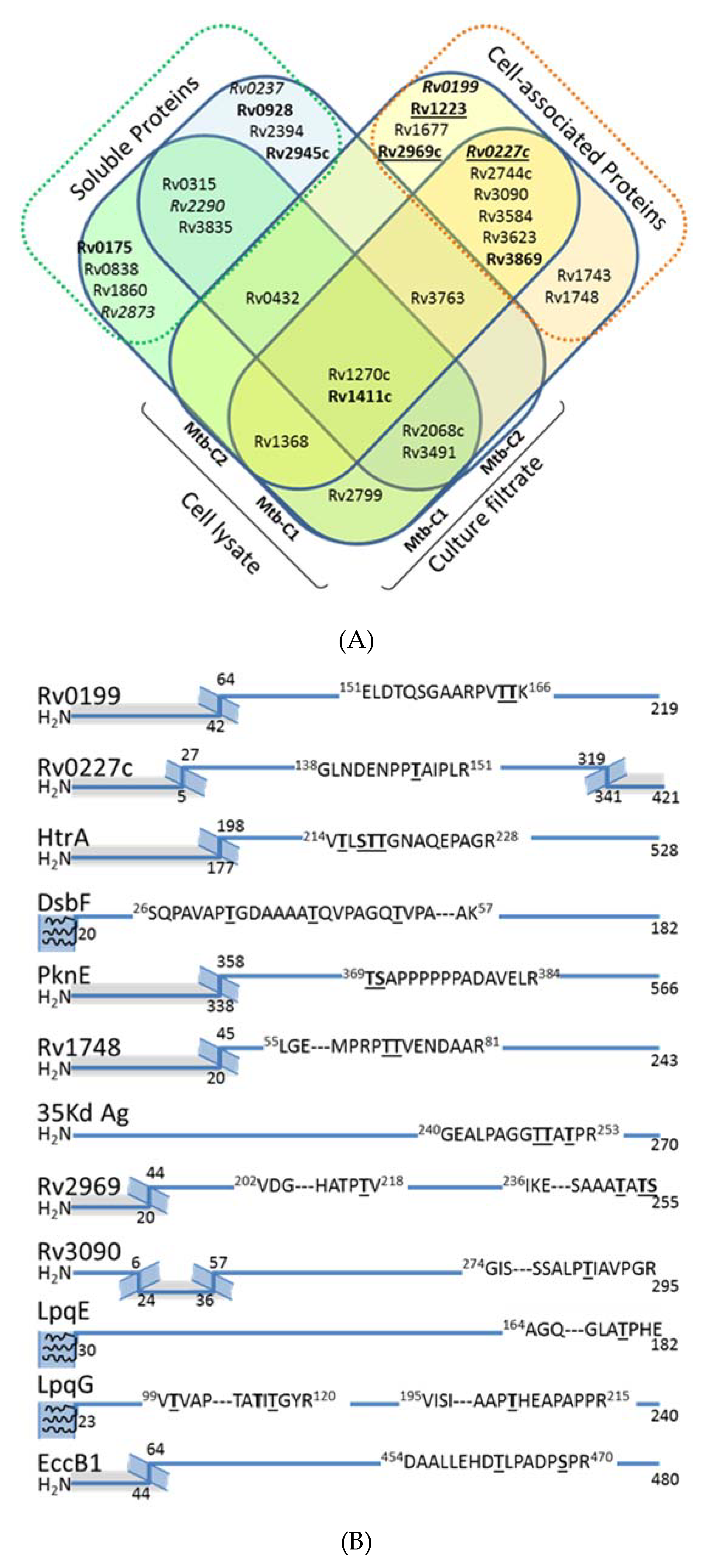
2.7. Exploration of the Mtb Cell Associated Glycoproteome
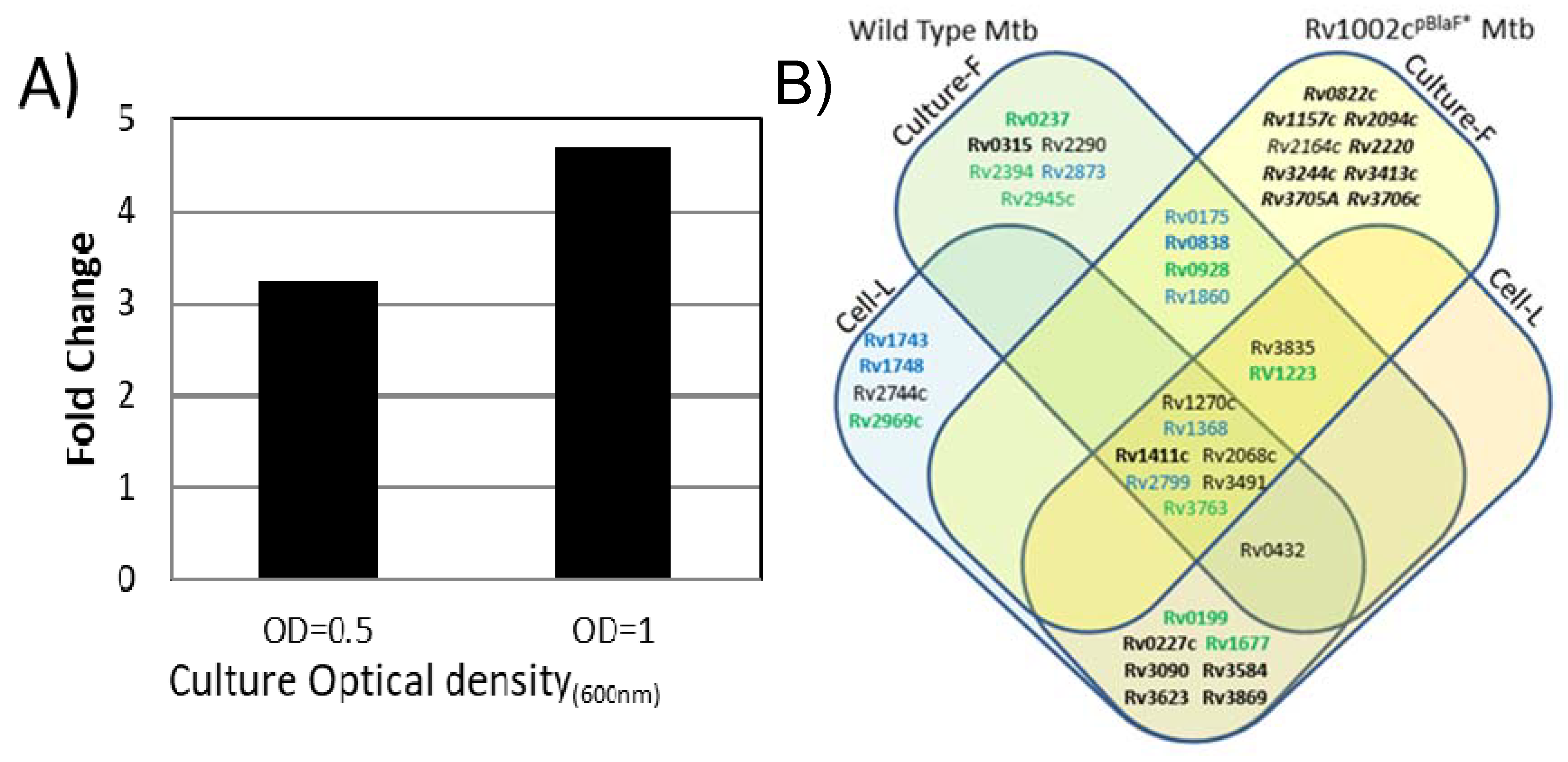
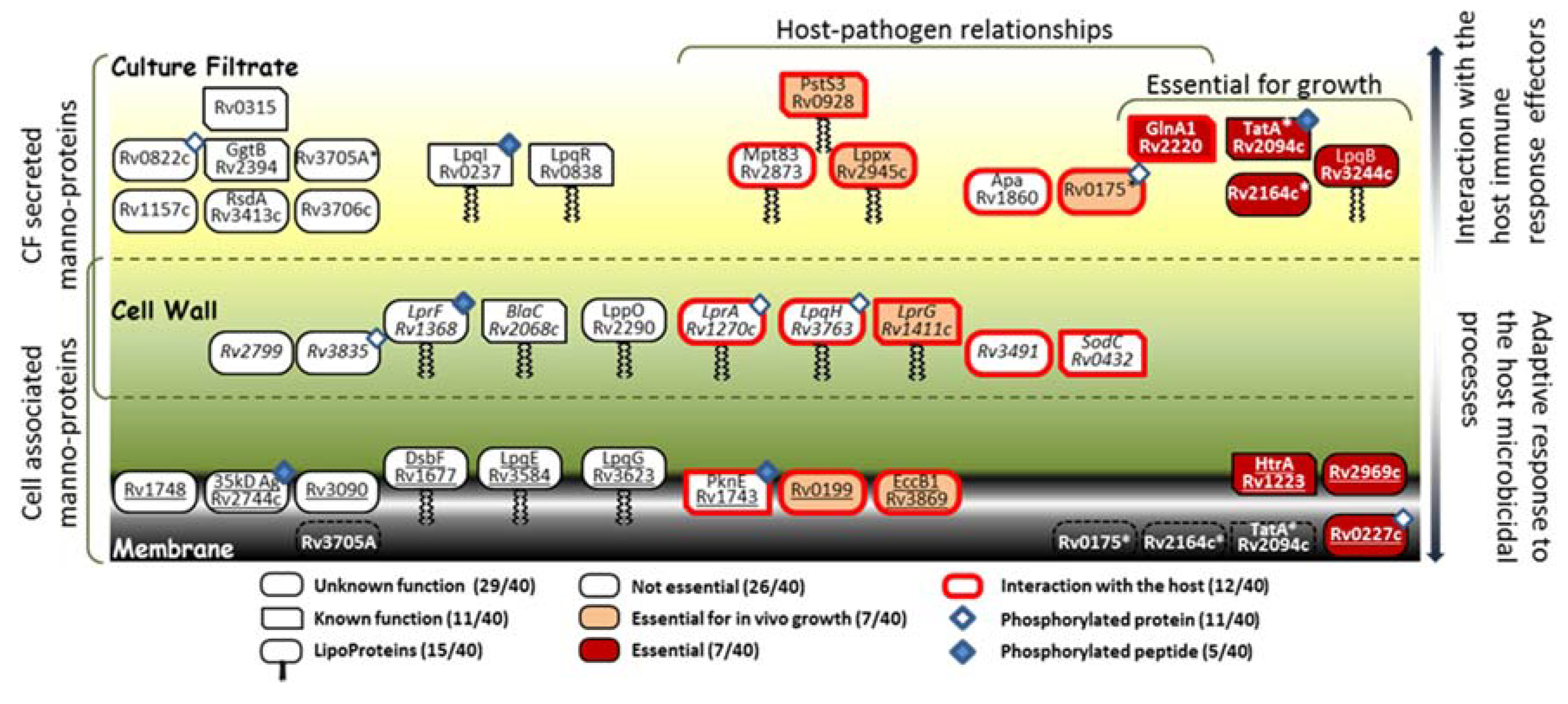
2.8. Potential Impact of the Mtb PMT Expression on the Mtb Glycoprotein Repertoire
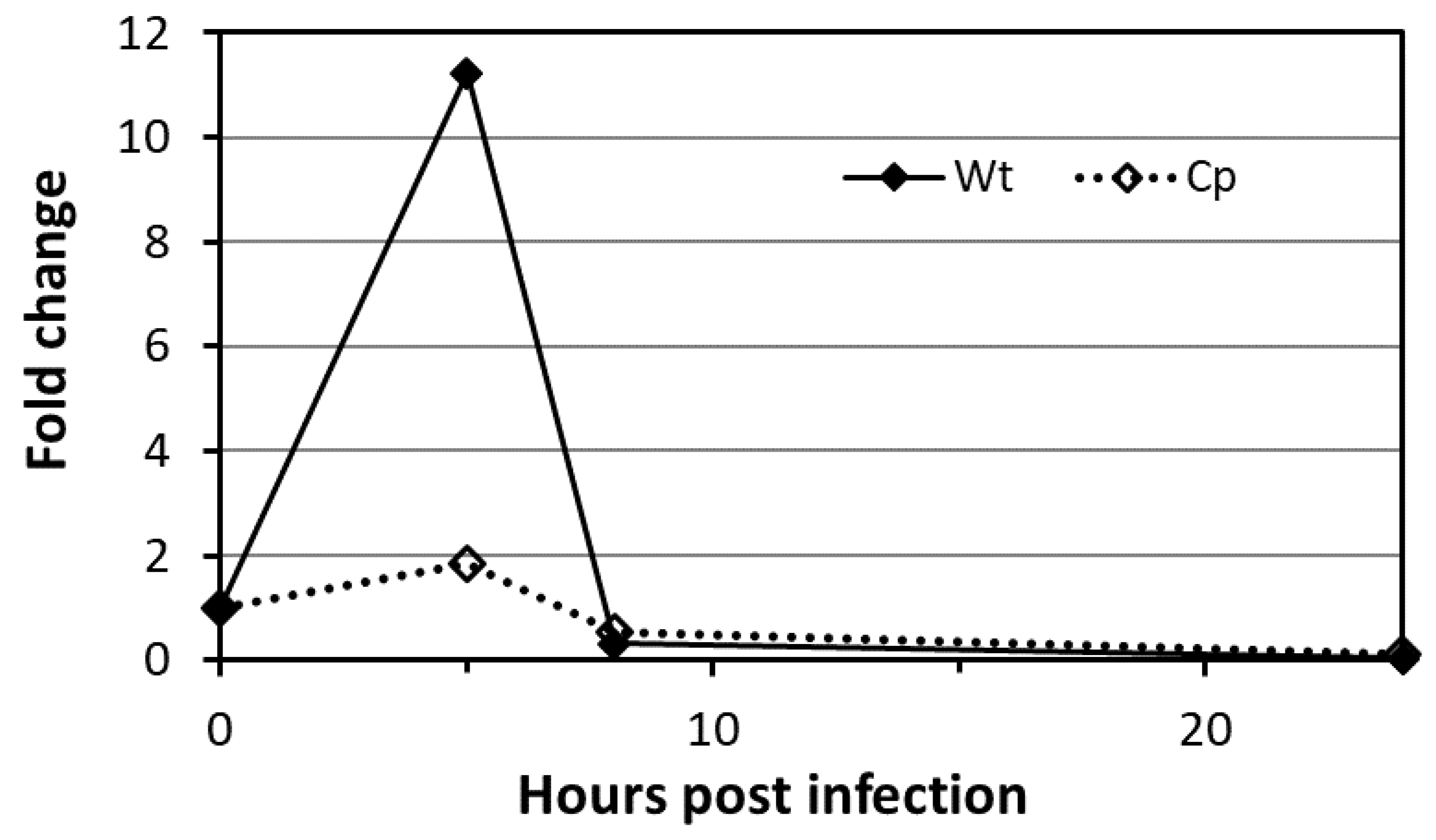
2.9. Mtb PMTub Expression Is Increased in the Macrophage
3. Materials and Methods
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Chauhan, V.; Thakur, S. State of the Globe: The Global Battle for Survival against Mycobacterium tuberculosis. J. Glob. Infect. Dis. 2017, 9, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Dickey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: Antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 2017, 16, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.F.; Tonini, L.; Malaga, W.; Beau, M.; Stella, A.; Bouyssie, D.; Jackson, M.C.; Nigou, J.; Puzo, G.; Guilhot, C.; et al. Bacterial protein-O-mannosylating enzyme is crucial for virulence of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2013, 110, 6560–6565. [Google Scholar] [CrossRef] [PubMed]
- Espitia, C.; Mancilla, R. Identification, isolation and partial characterization of Mycobacterium tuberculosis glycoprotein antigens. Clin. Exp. Immunol. 1989, 77, 378–383. [Google Scholar]
- Garbe, T.; Harris, D.; Vordermeier, M.; Lathigra, R.; Ivanyi, J.; Young, D. Expression of the Mycobacterium tuberculosis 19-kilodalton antigen in Mycobacterium smegmatis: Immunological analysis and evidence of glycosylation. Infect. Immun. 1993, 61, 260–267. [Google Scholar] [CrossRef]
- Dobos, K.M.; Swiderek, K.; Khoo, K.H.; Brennan, P.J.; Belisle, J.T. Evidence for glycosylation sites on the 45-kilodalton glycoprotein of Mycobacterium tuberculosis. Infect. Immun. 1995, 63, 2846–2853. [Google Scholar] [CrossRef]
- Michell, S.L.; Whelan, A.O.; Wheeler, P.R.; Panico, M.; Easton, R.L.; Etienne, A.T.; Haslam, S.M.; Dell, A.; Morris, H.R.; Reason, A.J.; et al. The MPB83 antigen from Mycobacterium bovis contains O-linked mannose and (1→3)-mannobiose moieties. J. Biol. Chem. 2003, 278, 16423–16432. [Google Scholar] [CrossRef]
- Sartain, M.J.; Belisle, J.T. N-Terminal clustering of the O-glycosylation sites in the Mycobacterium tuberculosis lipoprotein SodC. Glycobiology 2009, 19, 38–51. [Google Scholar] [CrossRef]
- Gonzalez-Zamorano, M.; Mendoza-Hernandez, G.; Xolalpa, W.; Parada, C.; Vallecillo, A.J.; Bigi, F.; Espitia, C. Mycobacterium tuberculosis glycoproteomics based on ConA-lectin affinity capture of mannosylated proteins. J. Proteome Res. 2009, 8, 721–733. [Google Scholar] [CrossRef]
- Brulle, J.K.; Grau, T.; Tschumi, A.; Auchli, Y.; Burri, R.; Polsfuss, S.; Keller, P.M.; Hunziker, P.; Sander, P. Cloning, expression and characterization of Mycobacterium tuberculosis lipoprotein LprF. Biochem. Biophys. Res. Commun. 2010, 391, 679–684. [Google Scholar] [CrossRef]
- Smith, G.T.; Sweredoski, M.J.; Hess, S. O-linked glycosylation sites profiling in Mycobacterium tuberculosis culture filtrate proteins. J. Proteom. 2014, 97, 296–306. [Google Scholar] [CrossRef][Green Version]
- Esparza, M.; Palomares, B.; Garcia, T.; Espinosa, P.; Zenteno, E.; Mancilla, R. PstS-1, the 38-kDa Mycobacterium tuberculosis glycoprotein, is an adhesin, which binds the macrophage mannose receptor and promotes phagocytosis. Scand. J. Immunol. 2015, 81, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Jagannathan, L.; Ganguli, G.; Padhi, A.; Roy, D.; Alaridah, N.; Saha, P.; Nongthomba, U.; Godaly, G.; Gopal, R.K.; et al. A mycobacterial phosphoribosyltransferase promotes bacillary survival by inhibiting oxidative stress and autophagy pathways in macrophages and zebrafish. J. Biol. Chem. 2015, 290, 13321–13343. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Zhang, F.; Yang, S.; Kang, J.; Sha, S.; Ma, Y. Mycobacterium tuberculosis Rv0431 expressed in Mycobacterium smegmatis, a potentially mannosylated protein, mediated the immune evasion of RAW 264.7 macrophages. Microb. Pathog. 2016, 100, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.L.; Delahay, R.; Gallagher, A.; Robertson, B.; Young, D. Analysis of post-translational modification of mycobacterial proteins using a cassette expression system. Febs Lett. 2000, 473, 358–362. [Google Scholar] [CrossRef]
- Campbell, M.P.; Aoki-Kinoshita, K.F.; Lisacek, F.; York, W.S.; Packer, N.H. Glycoinformatics. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017; pp. 667–679. [Google Scholar]
- Lisacek, F.; Mariethoz, J.; Alocci, D.; Rudd, P.M.; Abrahams, J.L.; Campbell, M.P.; Packer, N.H.; Stahle, J.; Widmalm, G.; Mullen, E.; et al. Databases and Associated Tools for Glycomics and Glycoproteomics. Methods Mol. Biol. 2017, 1503, 235–264. [Google Scholar] [CrossRef]
- Yang, Y.; Franc, V.; Heck, A.J.R. Glycoproteomics: A Balance between High-Throughput and In-Depth Analysis. Trends Biotechnol. 2017, 35, 598–609. [Google Scholar] [CrossRef]
- Bollineni, R.C.; Koehler, C.J.; Gislefoss, R.E.; Anonsen, J.H.; Thiede, B. Large-scale intact glycopeptide identification by Mascot database search. Sci. Rep. 2018, 8, 2117. [Google Scholar] [CrossRef]
- Hu, Y.; Shah, P.; Clark, D.J.; Ao, M.; Zhang, H. Reanalysis of Global Proteomic and Phosphoproteomic Data Identified a Large Number of Glycopeptides. Anal. Chem. 2018, 90, 8065–8071. [Google Scholar] [CrossRef]
- Liu, M.Q.; Zeng, W.F.; Fang, P.; Cao, W.Q.; Liu, C.; Yan, G.Q.; Zhang, Y.; Peng, C.; Wu, J.Q.; Zhang, X.J.; et al. pGlyco 2.0 enables precision N-glycoproteomics with comprehensive quality control and one-step mass spectrometry for intact glycopeptide identification. Nat. Commun. 2017, 8, 438. [Google Scholar] [CrossRef]
- Gaunitz, S.; Nagy, G.; Pohl, N.L.; Novotny, M.V. Recent Advances in the Analysis of Complex Glycoproteins. Anal. Chem. 2017, 89, 389–413. [Google Scholar] [CrossRef] [PubMed]
- Banazadeh, A.; Veillon, L.; Wooding, K.M.; Zabet-Moghaddam, M.; Mechref, Y. Recent advances in mass spectrometric analysis of glycoproteins. Electrophoresis 2017, 38, 162–189. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.L.; Chen, S.F. A Brief Review of Bioinformatics Tools for Glycosylation Analysis by Mass Spectrometry. Mass Spectrom. 2017, 6, S0064. [Google Scholar] [CrossRef] [PubMed]
- Dobos, K.M.; Khoo, K.H.; Swiderek, K.M.; Brennan, P.J.; Belisle, J.T. Definition of the full extent of glycosylation of the 45-kilodalton glycoprotein of Mycobacterium tuberculosis. J. Bacteriol. 1996, 178, 2498–2506. [Google Scholar] [CrossRef] [PubMed]
- Birhanu, A.G.; Yimer, S.A.; Kalayou, S.; Riaz, T.; Zegeye, E.D.; Holm-Hansen, C.; Norheim, G.; Aseffa, A.; Abebe, M.; Tonjum, T. Ample glycosylation in membrane and cell envelope proteins may explain the phenotypic diversity and virulence in the Mycobacterium tuberculosis complex. Sci. Rep. 2019, 9, 2927. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Nakano, M.; Hincapie, M.; Kolarich, D.; Baker, M.S.; Hancock, W.S.; Packer, N.H. The lectin riddle: Glycoproteins fractionated from complex mixtures have similar glycomic profiles. Omics A J. Integr. Biol. 2010, 14, 487–499. [Google Scholar] [CrossRef]
- Alley, W.R., Jr.; Mann, B.F.; Novotny, M.V. High-sensitivity analytical approaches for the structural characterization of glycoproteins. Chem. Rev. 2013, 113, 2668–2732. [Google Scholar] [CrossRef]
- Mehaffy, C.; Belisle, J.T.; Dobos, K.M. Mycobacteria and their sweet proteins: An overview of protein glycosylation and lipoglycosylation in M. tuberculosis. Tuberculosis 2019, 115, 1–13. [Google Scholar] [CrossRef]
- Mouton-Barbosa, E.; Roux-Dalvai, F.; Bouyssie, D.; Berger, F.; Schmidt, E.; Righetti, P.G.; Guerrier, L.; Boschetti, E.; Burlet-Schiltz, O.; Monsarrat, B.; et al. In-depth exploration of cerebrospinal fluid by combining peptide ligand library treatment and label-free protein quantification. Mol. Cell. Proteom. 2010, 9, 1006–1021. [Google Scholar] [CrossRef]
- Zaia, J. Mass spectrometry and glycomics. OMICS 2010, 14, 401–418. [Google Scholar] [CrossRef]
- Darula, Z.; Chalkley, R.J.; Lynn, A.; Baker, P.R.; Medzihradszky, K.F. Improved identification of O-linked glycopeptides from ETD data with optimized scoring for different charge states and cleavage specificities. Amino. Acids 2010. [Google Scholar] [CrossRef] [PubMed]
- Bouyssie, D.; Gonzalez de Peredo, A.; Mouton, E.; Albigot, R.; Roussel, L.; Ortega, N.; Cayrol, C.; Burlet-Schiltz, O.; Girard, J.P.; Monsarrat, B. Mascot file parsing and quantification (MFPaQ), a new software to parse, validate, and quantify proteomics data generated by ICAT and SILAC mass spectrometric analyses: Application to the proteomics study of membrane proteins from primary human endothelial cells. Mol. Cell. Proteom. 2007, 6, 1621–1637. [Google Scholar] [CrossRef]
- Kelkar, D.S.; Kumar, D.; Kumar, P.; Balakrishnan, L.; Muthusamy, B.; Yadav, A.K.; Shrivastava, P.; Marimuthu, A.; Anand, S.; Sundaram, H.; et al. Proteogenomic analysis of Mycobacterium tuberculosis by high resolution mass spectrometry. Mol. Cell. Proteom. 2011, 10, M111.011445. [Google Scholar] [CrossRef]
- Schubert, O.T.; Mouritsen, J.; Ludwig, C.; Rost, H.L.; Rosenberger, G.; Arthur, P.K.; Claassen, M.; Campbell, D.S.; Sun, Z.; Farrah, T.; et al. The Mtb proteome library: A resource of assays to quantify the complete proteome of Mycobacterium tuberculosis. Cell Host. Microb. 2013, 13, 602–612. [Google Scholar] [CrossRef]
- Albrethsen, J.; Agner, J.; Piersma, S.R.; Hojrup, P.; Pham, T.V.; Weldingh, K.; Jimenez, C.R.; Andersen, P.; Rosenkrands, I. Proteomic profiling of Mycobacterium tuberculosis identifies nutrient-starvation-responsive toxin-antitoxin systems. Mol. Cell. Proteom. 2013, 12, 1180–1191. [Google Scholar] [CrossRef]
- Roy, A.; Bhattacharya, S.; Bothra, A.K.; Sen, A. A database for Mycobacterium secretome analysis: ‘MycoSec’ to accelerate global health research. Omics A J. Integr. Biol. 2013, 17, 502–509. [Google Scholar] [CrossRef]
- Herrmann, J.L.; O’Gaora, P.; Gallagher, A.; Thole, J.E.; Young, D.B. Bacterial glycoproteins: A link between glycosylation and proteolytic cleavage of a 19 kDa antigen from Mycobacterium tuberculosis. EMBO J. 1996, 15, 3547–3554. [Google Scholar] [CrossRef]
- Alonso, H.; Parra, J.; Malaga, W.; Payros, D.; Liu, C.F.; Berrone, C.; Robert, C.; Meunier, E.; Burlet-Schiltz, O.; Riviere, M.; et al. Protein O-mannosylation deficiency increases LprG-associated lipoarabinomannan release by Mycobacterium tuberculosis and enhances the TLR2-associated inflammatory response. Sci. Rep. 2017, 7, 7913. [Google Scholar] [CrossRef]
- Parra, J.; Marcoux, J.; Poncin, I.; Canaan, S.; Herrmann, J.L.; Nigou, J.; Burlet-Schiltz, O.; Riviere, M. Scrutiny of Mycobacterium tuberculosis 19 kDa antigen proteoforms provides new insights in the lipoglycoprotein biogenesis paradigm. Sci. Rep. 2017, 7, 43682. [Google Scholar] [CrossRef]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef]
- Byun, E.H.; Kim, W.S.; Shin, A.R.; Kim, J.S.; Whang, J.; Won, C.J.; Choi, Y.; Kim, S.Y.; Koh, W.J.; Kim, H.J.; et al. Rv0315, a novel immunostimulatory antigen of Mycobacterium tuberculosis, activates dendritic cells and drives Th1 immune responses. J. Mol. Med. 2012, 90, 285–298. [Google Scholar] [CrossRef]
- Gaur, R.L.; Ren, K.; Blumenthal, A.; Bhamidi, S.; Gonzalez-Nilo, F.D.; Jackson, M.; Zare, R.N.; Ehrt, S.; Ernst, J.D.; Banaei, N. LprG-mediated surface expression of lipoarabinomannan is essential for virulence of Mycobacterium tuberculosis. PLoS Pathog. 2014, 10, e1004376. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Richardson, E.T.; Athman, J.J.; Shi, L.; Wearsch, P.A.; McDonald, D.; Banaei, N.; Boom, W.H.; Jackson, M.; Harding, C.V. Mycobacterium tuberculosis lipoprotein LprG binds lipoarabinomannan and determines its cell envelope localization to control phagolysosomal fusion. PLoS Pathog. 2014, 10, e1004471. [Google Scholar] [CrossRef] [PubMed]
- Mohiman, N.; Argentini, M.; Batt, S.M.; Cornu, D.; Masi, M.; Eggeling, L.; Besra, G.; Bayan, N. The ppm operon is essential for acylation and glycosylation of lipoproteins in Corynebacterium glutamicum. PLoS ONE 2012, 7, e46225. [Google Scholar] [CrossRef] [PubMed]
- Zech, H.; Echtermeyer, C.; Wohlbrand, L.; Blasius, B.; Rabus, R. Biological versus technical variability in 2-D DIGE experiments with environmental bacteria. Proteomics 2011, 11, 3380–3389. [Google Scholar] [CrossRef]
- Sridhar, S.; Steele-Mortimer, O. Inherent Variability of Growth Media Impacts the Ability of Salmonella Typhimurium to Interact with Host Cells. PLoS ONE 2016, 11, e0157043. [Google Scholar] [CrossRef]
- Gamboa-Suasnavart, R.A.; Valdez-Cruz, N.A.; Cordova-Davalos, L.E.; Martinez-Sotelo, J.A.; Servin-Gonzalez, L.; Espitia, C.; Trujillo-Roldan, M.A. The O-mannosylation and production of recombinant APA (45/47 KDa) protein from Mycobacterium tuberculosis in Streptomyces lividans is affected by culture conditions in shake flasks. Microb Cell Fact. 2011, 10, 110. [Google Scholar] [CrossRef]
- Rodenburg, R.N.P.; Snijder, J.; van de Waterbeemd, M.; Schouten, A.; Granneman, J.; Heck, A.J.R.; Gros, P. Stochastic palmitoylation of accessible cysteines in membrane proteins revealed by native mass spectrometry. Nat. Commun. 2017, 8, 1280. [Google Scholar] [CrossRef]
- VanderVen, B.C.; Harder, J.D.; Crick, D.C.; Belisle, J.T. Export-mediated assembly of mycobacterial glycoproteins parallels eukaryotic pathways. Science 2005, 309, 941–943. [Google Scholar] [CrossRef]
- De Souza, G.A.; Leversen, N.A.; Malen, H.; Wiker, H.G. Bacterial proteins with cleaved or uncleaved signal peptides of the general secretory pathway. J. Proteom. 2011, 75, 502–510. [Google Scholar] [CrossRef]
- Peters, J.S.; Calder, B.; Gonnelli, G.; Degroeve, S.; Rajaonarifara, E.; Mulder, N.; Soares, N.C.; Martens, L.; Blackburn, J.M. Identification of Quantitative Proteomic Differences between Mycobacterium tuberculosis Lineages with Altered Virulence. Front. Microbiol. 2016, 7, 813. [Google Scholar] [CrossRef]
- Timm, J.; Perilli, M.G.; Duez, C.; Trias, J.; Orefici, G.; Fattorini, L.; Amicosante, G.; Oratore, A.; Joris, B.; Frere, J.M.; et al. Transcription and expression analysis, using lacZ and phoA gene fusions, of Mycobacterium fortuitum beta-lactamase genes cloned from a natural isolate and a high-level beta-lactamase producer. Mol. Microbiol. 1994, 12, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.E.; Gawronski, J.D.; Dejesus, M.A.; Ioerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011, 7, e1002251. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Wolff, K.A.; Cartabuke, R.H.; Ogwang, S.; Nguyen, L. A lipoprotein modulates activity of the MtrAB two-component system to provide intrinsic multidrug resistance, cytokinetic control and cell wall homeostasis in Mycobacterium. Mol. Microbiol. 2010, 76, 348–364. [Google Scholar] [CrossRef] [PubMed]
- Talaat, A.M.; Lyons, R.; Howard, S.T.; Johnston, S.A. The temporal expression profile of Mycobacterium tuberculosis infection in mice. Proc. Natl. Acad. Sci. USA 2004, 101, 4602–4607. [Google Scholar] [CrossRef]
- Turkarslan, S.; Peterson, E.J.; Rustad, T.R.; Minch, K.J.; Reiss, D.J.; Morrison, R.; Ma, S.; Price, N.D.; Sherman, D.R.; Baliga, N.S. A comprehensive map of genome-wide gene regulation in Mycobacterium tuberculosis. Sci. Data 2015, 2, 150010. [Google Scholar] [CrossRef]
- Galagan, J.E.; Minch, K.; Peterson, M.; Lyubetskaya, A.; Azizi, E.; Sweet, L.; Gomes, A.; Rustad, T.; Dolganov, G.; Glotova, I.; et al. The Mycobacterium tuberculosis regulatory network and hypoxia. Nature 2013, 499, 178–183. [Google Scholar] [CrossRef]
- Schultz, J.C.; Takayama, K. The role of mannosylphosphorylpolyisoprenol in glycoprotein biosynthesis in Mycobacterium smegmatis. Biochim. Biophys. Acta 1975, 381, 175–184. [Google Scholar] [CrossRef]
- Chan, C.E.; Gotze, S.; Seah, G.T.; Seeberger, P.H.; Tukvadze, N.; Wenk, M.R.; Hanson, B.J.; MacAry, P.A. The diagnostic targeting of a carbohydrate virulence factor from M. Tuberculosis. Sci. Rep. 2015, 5, 10281. [Google Scholar] [CrossRef]
- Facciuolo, A.; Mutharia, L.M. Mycobacterial glycoproteins: A novel subset of vaccine candidates. Front. Cell Infect. Microbiol. 2014, 4, 133. [Google Scholar] [CrossRef]
- Van Els, C.A.; Corbiere, V.; Smits, K.; van Gaans-van den Brink, J.A.; Poelen, M.C.; Mascart, F.; Meiring, H.D.; Locht, C. Toward Understanding the Essence of Post-Translational Modifications for the Mycobacterium tuberculosis Immunoproteome. Front. Immunol. 2014, 5, 361. [Google Scholar] [CrossRef]
- Espitia, C.; Servin-Gonzalez, L.; Mancilla, R. New insights into protein O-mannosylation in actinomycetes. Mol. Biosyst. 2010, 6, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Romain, F.; Augier, J.; Pescher, P.; Marchal, G. Isolation of a proline-rich mycobacterial protein eliciting delayed-type hypersensitivity reactions only in guinea pigs immunized with living mycobacteria. Proc. Natl. Acad. Sci. USA 1993, 90, 5322–5326. [Google Scholar] [CrossRef] [PubMed]
- Espitia, C.; Espinosa, R.; Saavedra, R.; Mancilla, R.; Romain, F.; Laqueyrerie, A.; Moreno, C. Antigenic and structural similarities between Mycobacterium tuberculosis 50- to 55-kilodalton and Mycobacterium bovis BCG 45- to 47-kilodalton antigens. Infect. Immun. 1995, 63, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Bando-Campos, G.; Juarez-Lopez, D.; Roman-Gonzalez, S.A.; Castillo-Rodal, A.I.; Olvera, C.; Lopez-Vidal, Y.; Arreguin-Espinosa, R.; Espitia, C.; Trujillo-Roldan, M.A.; Valdez-Cruz, N.A. Recombinant O-mannosylated protein production (PstS-1) from Mycobacterium tuberculosis in Pichia pastoris (Komagataella phaffii) as a tool to study tuberculosis infection. Microb. Cell Fact. 2019, 18, 11. [Google Scholar] [CrossRef]
- Pitarque, S.; Herrmann, J.L.; Duteyrat, J.L.; Jackson, M.; Stewart, G.R.; Lecointe, F.; Payre, B.; Schwartz, O.; Young, D.B.; Marchal, G.; et al. Deciphering the molecular bases of Mycobacterium tuberculosis binding to the lectin DC-SIGN reveals an underestimated complexity. Biochem. J. 2005, 392, 615–624. [Google Scholar] [CrossRef]
- Ragas, A.; Roussel, L.; Puzo, G.; Riviere, M. The Mycobacterium tuberculosis cell-surface glycoprotein apa as a potential adhesin to colonize target cells via the innate immune system pulmonary C-type lectin surfactant protein A. J. Biol. Chem. 2007, 282, 5133–5142. [Google Scholar] [CrossRef]
- Horn, C.; Namane, A.; Pescher, P.; Riviere, M.; Romain, F.; Puzo, G.; Barzu, O.; Marchal, G. Decreased capacity of recombinant 45/47-kDa molecules (Apa) of Mycobacterium tuberculosis to stimulate T lymphocyte responses related to changes in their mannosylation pattern. J. Biol. Chem. 1999, 274, 32023–32030. [Google Scholar] [CrossRef]
- Romain, F.; Horn, C.; Pescher, P.; Namane, A.; Riviere, M.; Puzo, G.; Barzu, O.; Marchal, G. Deglycosylation of the 45/47-kilodalton antigen complex of Mycobacterium tuberculosis decreases its capacity to elicit in vivo or in vitro cellular immune responses. Infect. Immun. 1999, 67, 5567–5572. [Google Scholar] [CrossRef]
- Nandakumar, S.; Kannanganat, S.; Dobos, K.M.; Lucas, M.; Spencer, J.S.; Fang, S.; McDonald, M.A.; Pohl, J.; Birkness, K.; Chamcha, V.; et al. O-mannosylation of the Mycobacterium tuberculosis adhesin Apa is crucial for T cell antigenicity during infection but is expendable for protection. PLoS Pathog. 2013, 9, e1003705. [Google Scholar] [CrossRef]
- Wilkinson, K.A.; Newton, S.M.; Stewart, G.R.; Martineau, A.R.; Patel, J.; Sullivan, S.M.; Herrmann, J.L.; Neyrolles, O.; Young, D.B.; Wilkinson, R.J. Genetic determination of the effect of post-translational modification on the innate immune response to the 19 kDa lipoprotein of Mycobacterium tuberculosis. BMC Microbiol. 2009, 9, 93. [Google Scholar] [CrossRef]
- Sieling, P.A.; Hill, P.J.; Dobos, K.M.; Brookman, K.; Kuhlman, A.M.; Fabri, M.; Krutzik, S.R.; Rea, T.H.; Heaslip, D.G.; Belisle, J.T.; et al. Conserved mycobacterial lipoglycoproteins activate TLR2 but also require glycosylation for MHC class II-restricted T cell activation. J. Immunol. 2008, 180, 5833–5842. [Google Scholar] [CrossRef] [PubMed]
- Sassetti, C.M.; Rubin, E.J. Genetic requirements for mycobacterial survival during infection. Proc. Natl. Acad. Sci. USA 2003, 100, 12989–12994. [Google Scholar] [CrossRef] [PubMed]
- Sassetti, C.M.; Boyd, D.H.; Rubin, E.J. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 2003, 48, 77–84. [Google Scholar] [CrossRef] [PubMed]
- DeJesus, M.A.; Gerrick, E.R.; Xu, W.; Park, S.W.; Long, J.E.; Boutte, C.C.; Rubin, E.J.; Schnappinger, D.; Ehrt, S.; Fortune, S.M.; et al. Comprehensive Essentiality Analysis of the Mycobacterium tuberculosis Genome via Saturating Transposon Mutagenesis. mBio 2017, 8, e02133-16. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.M.; Ocampo, M.; Curtidor, H.; Vanegas, M.; Patarroyo, M.E.; Patarroyo, M.A. Mycobacterium tuberculosis surface protein Rv0227c contains high activity binding peptides which inhibit cell invasion. Peptides 2012, 38, 208–216. [Google Scholar] [CrossRef]
- White, M.J.; Savaryn, J.P.; Bretl, D.J.; He, H.; Penoske, R.M.; Terhune, S.S.; Zahrt, T.C. The HtrA-like serine protease PepD interacts with and modulates the Mycobacterium tuberculosis 35-kDa antigen outer envelope protein. PLoS ONE 2011, 6, e18175. [Google Scholar] [CrossRef]
- Majlessi, L.; Prados-Rosales, R.; Casadevall, A.; Brosch, R. Release of mycobacterial antigens. Immunol Rev. 2015, 264, 25–45. [Google Scholar] [CrossRef]
- Choi, S.; Choi, H.G.; Lee, J.; Shin, K.W.; Kim, H.J. Mycobacterium tuberculosis protein Rv2220 induces maturation and activation of dendritic cells. Cell Immunol. 2018, 328, 70–78. [Google Scholar] [CrossRef]
- Premkumar, L.; Heras, B.; Duprez, W.; Walden, P.; Halili, M.; Kurth, F.; Fairlie, D.P.; Martin, J.L. Rv2969c, essential for optimal growth in Mycobacterium tuberculosis, is a DsbA-like enzyme that interacts with VKOR-derived peptides and has atypical features of DsbA-like disulfide oxidases. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1981–1994. [Google Scholar] [CrossRef]
- Fortuin, S.; Tomazella, G.G.; Nagaraj, N.; Sampson, S.L.; Gey van Pittius, N.C.; Soares, N.C.; Wiker, H.G.; de Souza, G.A.; Warren, R.M. Phosphoproteomics analysis of a clinical Mycobacterium tuberculosis Beijing isolate: Expanding the mycobacterial phosphoproteome catalog. Front. Microbiol 2015, 6, 6. [Google Scholar] [CrossRef]
- Prisic, S.; Dankwa, S.; Schwartz, D.; Chou, M.F.; Locasale, J.W.; Kang, C.M.; Bemis, G.; Church, G.M.; Steen, H.; Husson, R.N. Extensive phosphorylation with overlapping specificity by Mycobacterium tuberculosis serine/threonine protein kinases. Proc. Natl. Acad. Sci. USA 2010, 107, 7521–7526. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Pinto, S.M.; Patil, A.H.; Advani, J.; Subba, P.; Kumar, M.; Sharma, J.; Dey, G.; Ravikumar, R.; Buggi, S.; et al. Quantitative Proteomic and Phosphoproteomic Analysis of H37Ra and H37Rv Strains of Mycobacterium tuberculosis. J. Proteome Res. 2017, 16, 1632–1645. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, L.; Liu, B.; Jin, Q. Phosphoproteomic analysis of bacillus Calmette-Guerin using gel-based and gel-free approaches. J. Proteom. 2015, 126, 189–199. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Protein Description | Exp_Mr | Mr_Delta | Start-End | Score | Pep_Seq | Var_Mod | Ref. |
|---|---|---|---|---|---|---|---|
| RV0175 MCE-associated membrane protein | 5175.19 | −0.023 | 100–144 | 16.97 | DCVAATQAPDAGAMSASMQKIIECGTGDFGAQASLYTSMLVEAYQ | 3 Hex, Ox, Pr | [12] |
| 3311.31 | 0.006 | 100–124 | 12.85 | DCVAATQAPDAGAMSASMQKIIECG | 5 Hex, 2 Ox | ||
| Rv0432 superoxide dismutase [Cu-Zn] | 3480.57 | 0.009 | 53–83 | 34.27 | SPAPSGLSGHDEESPGAQSLTSTLTAPDGTK | 3 Hex | [9] |
| 3134.43 | 0.007 | 55–83 | 15.87 | APSGLSGHDEESPGAQSLTSTLTAPDGTK | 2 Hex | ||
| Rv1270c lipoprotein lprA | 2356.01 | 0.004 | 34–51 | 17.75 | ASDTAATASNGDAAMLLK | 4 Hex | [12] |
| 2193.97 | 0.008 | 34–51 | 53.89 | ASDTAATASNGDAAMLLK | 3 Hex | ||
| 2047.91 | 0.006 | 34–51 | 82.16 | ASDTAATASNGDAAMLLK | 2 Hex, Ox | ||
| 2031.91 | 0.007 | 34–51 | 75.72 | ASDTAATASNGDAAMLLK | 2 Hex | ||
| 1885.85 | 0.006 | 34–51 | 79.85 | ASDTAATASNGDAAMLLK | 1 Hex, Ox | ||
| 1869.86 | 0.005 | 34–51 | 80.64 | ASDTAATASNGDAAMLLK | 1 Hex | ||
| RV1368 lipoprotein lprF | 3110.46 | 0.010 | 42–67 | 40.23 | KPTTASSPSPGSPSPEAQQILQDSSK | 3 Hex | [11] |
| 2948.41 | 0.007 | 42–67 | 45.9 | KPTTASSPSPGSPSPEAQQILQDSSK | 2 Hex | ||
| 2786.35 | 0.008 | 42–67 | 41.05 | KPTTASSPSPGSPSPEAQQILQDSSK | 1 Hex | ||
| 2359.11 | 0.003 | 46–67 | 11.61 | ASSPSPGSPSPEAQQILQDSSK | 1 Hex | ||
| RV1860 alanine and proline-rich secreted protein apa | 4563.39 | 0.012 | 278–321 | 22.07 | ALAESIRPLVAPPPAPAPAPAEPAPAPAPAGEVAPTPTTPTPQR | 2 Hex | [26] |
| 4401.33 | 0.010 | 278–321 | 29.05 | ALAESIRPLVAPPPAPAPAPAEPAPAPAPAGEVAPTPTTPTPQR | 1 Hex | ||
| 2263.07 | −0.012 | 309–325 | 20.43 | EVAPTPTTPTPQRTLPA | 3 Hex | ||
| RV2068c beta-lactamase | 2747.30 | −0.017 | 25–47 | 17 | ARHASGARPASTTLPAGADLADR | 3 Hex | [12] |
| 2005.98 | 0.000 | 31–47 | 27.16 | ARPASTTLPAGADLADR | 2 Hex | ||
| 1934.94 | 0.001 | 32–47 | 28.64 | RPASTTLPAGADLADR | 2 Hex | ||
| 1778.85 | 0.005 | 33–47 | 20.07 | PASTTLPAGADLADR | 2 Hex | ||
| 1772.81 | 0.004 | 35–47 | 16.3 | STTLPAGADLADR | 3 Hex | ||
| 1610.75 | 0.000 | 35–47 | 19.21 | STTLPAGADLADR | 2 Hex | ||
| 1523.72 | 0.001 | 36–47 | 20.21 | TTLPAGADLADR | 2 Hex | ||
| RV2290 hypothetical protein | 4998.26 | −0.006 | 47–85 | 13.09 | ATVMVEGHTHTISGVVECRTSPAVRTATPSESGTQTTR | 6 Hex | [12] |
| 1497.68 | 0.003 | 73–85 | 23.23 | TATPSESGTQTTR | 1 Hex | ||
| RV2799 hypothetical protein | 2096.95 | 0.003 | 68–83 | 26.33 | SPIVATTDPSPFDPCR | 2 Hex, Pr | [12] |
| 2082.94 | 0.006 | 68–83 | 61.05 | SPIVATTDPSPFDPCR | 2 Hex, Ca | ||
| 1920.88 | 0.002 | 68–83 | 37.81 | SPIVATTDPSPFDPCR | 1 Hex, Ca | ||
| RV2873 cell surface lipoprotein MPB83 | 7063.14 | −0.022 | 44–106 | 11.82 | AAPVTTAAMADPAADLIGRGCAQYAAQNPTGPGSVAGMAQDPVATAASNNPMLSTLTSALSGK | 6 Hex, 2Ox | [8] |
| 4982.32 | 0.012 | 21–62 | 10.69 | FLAGCSSTKPVSQDTSPKPATSPAAPVTTAAMADPAADLIGR | 5 Hex, Ox | ||
| 1974.89 | 0.003 | 48–62 | 14.46 | TTAAMADPAADLIGR | 3 Hex, Ox | ||
| 1958.90 | 0.011 | 48–62 | 43.02 | TTAAMADPAADLIGR | 3 Hex | ||
| RV3491 hypothetical protein | 2022.08 | 0.005 | 164–180 | 25.55 | QPFSLQLIGPPPSPVQR | 1 Hex | [12] |
| 1649.91 | 0.007 | 167–180 | 36.78 | SLQLIGPPPSPVQR | 1 Hex | ||
| RV3835 hypothetical protein | 6066.10 | −0.001 | 362–415 | 20.03 | LNLPPIPLQLPTPRPAPPAQQLPSTPPGTQHLPAQQPVVTPTRPPESHAPASAA | 3 Hex | [12] |
| 1730.00 | −0.001 | 362–375 | 10.6 | LNLPPIPLQLPTPR | 1 Hex |
| Gene / Protein Description | Exp_Mr | Mr_Delta | Start-End | Score | Sequence (Predicted Glycosylation Sites) | Var_Mod | #MS2 |
|---|---|---|---|---|---|---|---|
| RV0315 hypothetical protein | 3615.74 | 0.007 | 38–74 | 47.14 | APAGPTPAPAAPAAATGGLLFHDEFDGPAGSVPDPSK § | 1 Hex | 11 |
| 3453.68 | −0.001 | 38–74 | 46.3 | APAGPTPAPAAPAAATGGLLFHDEFDGPAGSVPDPSK § | (1) | ||
| 3544.70 | 0.005 | 39–74 | 28.62 | PAGPTPAPAAPAAATGGLLFHDEFDGPAGSVPDPSK | 1 Hex | 7 | |
| 3382.64 | −0.010 | 39–74 | 36.27 | PAGPTPAPAAPAAATGGLLFHDEFDGPAGSVPDPSK | (1) | ||
| RV0838 D-alanyl-D-alanine dipeptidase | 2051.97 | 0.007 | 47–62 | 11.36 | TTTPSGPVPPVSEAAR * | 3 Hex | 1 |
| 1889.92 | 0.006 | 47–62 | 18.28 | TTTPSGPVPPVSEAAR * | 2 Hex | 6 | |
| RV1411c lipoprotein lprG | 1115.61 | 0.001 | 228–236 | 13.65 | VQVTKPPVS § | 1 Hex | 2 |
| 953.55 | −0.001 | 228–236 | 39.14 | VQVTKPPVS § | (12) | ||
| RV1625c adenylate cyclase | 1658.73 | 0.010 | 28–37 | 16.01 | TQARAPTQHY | 3Hex, Da | 2 |
| RV1731 succinate-dehydrogenase | 1632.82 | 0.004 | 2–14 | 26.43 | PAPSAEVFDRLRN | 1 Hex | 4 |
| RV1935c enoyl-CoA hydratase | 1497.68 | −0.001 | 62–73 | 16.82 | AFELAEADDTVR | 1 Hex | 2 |
| Rv2563 ABC transporter | 1705.82 | 0.004 | 188–202 | 12.89 | GQPTISSIGIDGMPR | 1 Hex, Ox | 3 |
| Rv3209 Hypothetical protein | 3813.80 | −0.002 | 156–186 | 12.78 | LYSRLNCSIVNTGAQTVVASTNNAIIATCTR | 3 Hex, 3 Da, Pr | 2 |
| Peptide Sequence | Exper. Mr | Expec. Mr | Modification | Score | Gene: Protein Description |
|---|---|---|---|---|---|
| GSGDSHNGGVTTPPLDDLVGDDLVPYR * | 2915.33 | 2752.28 | Hex, Da | 19.64 | Rv0237: beta-glucosidase |
| APAGPTPAPAAPAAATGGLLFHDEFDGPAGSVPDPSK | 3615.74 | 3453.67 | Hex | 27.93 | Rv0315: hypothetical protein |
| SPAPSGLSGHDEESPGAQSLTSTLTAPDGTK | 3480.57 | 2994.40 | 3 Hex | 21.75 | Rv0432: superoxide dismutase |
| GNDDNVTGGGATTGQASAK * | 2205.93 | 1719.76 | 3 Hex | 47.62 | Rv0928: phosphate-binding protein pstS 3 |
| ASDTAATASNGDAAMLLK | 1885.85 | 1706.80 | Hex, Ox, Da | 87.28 | Rv1270c: lipoprotein lprA |
| TLSGDLTTNPTAATGNVK | 1922.94 | 1759.88 | Hex, Da | 12.82 | Rv1411c: lipoprotein lprG |
| ASTTLPAGADLADR | 1681.79 | 1357.67 | 2 Hex | 19.5 | Rv2068c: beta-lactamase |
| TATPSESGTQTTR | 1497.68 | 1335.62 | Hex | 36.17 | Rv2290: hypothetical protein |
| GAPSTAGPCEIVPNGTPAPK | 2082.98 | 1862.91 | Hex, Ca, Da | 30.21 | Rv2394: gamma-glutamyltransferase |
| VPVSPTASDPALLAEIR | 2059.06 | 1734.94 | 2 Hex | 35.43 | Rv2945c: lipoprotein lPpx |
| QPFSLQLIGPPPSPVQR | 2022.08 | 1860.02 | Hex | 23 | Rv3491: hypothetical protein |
| GETTTAAGTTASPGAASGPK | 2380.04 | 1731.82 | 4 Hex | 36.99 | Rv3763: lipoprotein lpqH |
| LNLPPIPLQLPTPR | 1730.00 | 1567.93 | Hex | 33.46 | Rv3835: hypothetical protein |
| Gene | Protein Description | exp_mr | mr_Delta (ppm) | Start-End | Score | Peptide Sequence | Var_Mod | Wt/Cp |
|---|---|---|---|---|---|---|---|---|
| RV0199 | hypothetical protein | 1833.94 | 4.39 | 151–166 | 28.45 | (R)ELDTQSGAARPVVTTK(L) | Hex | Wt/Cp |
| 1671.88 | 2.09 | 151–166 | 54.66 | (R)ELDTQSGAARPVVTTK(L) | Wt/Cp | |||
| RV0227c | hypothetical protein | 1667.84 | 0.02 | 138–151 | 28.6 | (R)GLNDENPPTAIPLR(H) | Hex | Wt/Cp |
| 1505.78 | 0.57 | 138–151 | 49.36 | (R)GLNDENPPTAIPLR(H) | Wt/Cp | |||
| RV0432 | superoxide dismutase [Cu-Zn] | 2966.33 | −1.00 | 57–83 | 30.15 | (P)SGLSGHDEESPGAQSLTSTLTAPDGTK(V) | 2 Hex | Wt/Cp |
| 3480.56 | 0.06 | 53–83 | 18.28 | (G)SPAPSGLSGHDEESPGAQSLTSTLTAPDGTK(V) | 3 Hex | Wt/Cp | ||
| RV1223 | htrA | 1662.80 | −2.39 | 214–228 | 50.29 | (K)VTLSTTGNAQEPAGR(F) | Hex | -/Cp |
| 1501.74 | 0.11 | 214–228 | 76.54 | (K)VTLSTTGNAQEPAGR(F) | Da | -/Cp | ||
| RV1270c | lipoprotein lprA | 2356.01 | −0.18 | 34–51 | 75.21 | (K)ASDTAATASNGDAAMLLK(Q) | 4 Hex, Da | Wt/Cp |
| 2193.96 | 1.41 | 34–51 | 66.26 | (K)ASDTAATASNGDAAMLLK(Q) | 3 Hex, Da | -/Cp | ||
| 2030.93 | 2.89 | 34–51 | 79.32 | (K)ASDTAATASNGDAAMLLK(Q) | 2 Hex | Wt/Cp | ||
| 1869.85 | −0.08 | 34–51 | 62.35 | (K)ASDTAATASNGDAAMLLK(Q) | Hex, Da | Wt/Cp | ||
| 1868.87 | 3.90 | 34–51 | 67.93 | (K)ASDTAATASNGDAAMLLK(Q) | Hex | Wt/Cp | ||
| 1707.80 | −0.75 | 34–51 | 85.51 | (K)ASDTAATASNGDAAMLLK(Q) | Da | Wt/Cp | ||
| RV1368 | lipoprotein lprF | 3272.50 | −0.54 | 42–67 | 43.73 | (K)KPTTASSPSPGSPSPEAQQILQDSSK(A) | 4 Hex | Wt/Cp |
| 3110.45 | −0.19 | 42–67 | 47.88 | (K)KPTTASSPSPGSPSPEAQQILQDSSK(A) | 3 Hex | Wt/Cp | ||
| 2948.39 | −2.28 | 42–67 | 61.82 | (K)KPTTASSPSPGSPSPEAQQILQDSSK(A) | 2 Hex | Wt/Cp | ||
| 2786.34 | −0.74 | 42–67 | 89.33 | (K)KPTTASSPSPGSPSPEAQQILQDSSK(A) | Hex | Wt/Cp | ||
| 2624.29 | −0.54 | 42–67 | 72.17 | (K)KPTTASSPSPGSPSPEAQQILQDSSK(A) | Wt/Cp | |||
| RV1411c | lipoprotein lprG | 2084.00 | −1.07 | 75–92 | 67.17 | (K)TLSGDLTTNPTAATGNVK(L) | 2 Hex | -/Cp |
| 1921.95 | 0.15 | 75–92 | 96.46 | (K)TLSGDLTTNPTAATGNVK(L) | Hex | Wt/Cp | ||
| 1759.89 | −0.41 | 75–92 | 98.98 | (K)TLSGDLTTNPTAATGNVK(L) | Wt/Cp | |||
| 1277.66 | −0.19 | 228–236 | 11.3 | (K)VQVTKPPVS(-) | 2 Hex | Wt/- | ||
| 1115.61 | −1.23 | 228–236 | 24.38 | (K)VQVTKPPVS(-) | Hex | Wt/Cp | ||
| 953.55 | −2.24 | 228–236 | 40.29 | (K)VQVTKPPVS(-) | Wt/Cp | |||
| RV1677 | hypothetical protein dsbF | 3563.73 | −1.49 | 26–57 | 11 | (K)SQPAVAPTGDAAAATQVPAGQTVPAQLQFSAK(T) | 3 Hex | Wt/Cp |
| 3401.68 | −0.35 | 26–57 | 32.98 | (K)SQPAVAPTGDAAAATQVPAGQTVPAQLQFSAK(T) | 2 Hex | Wt/Cp | ||
| 3239.63 | 0.45 | 26–57 | 49.38 | (K)SQPAVAPTGDAAAATQVPAGQTVPAQLQFSAK(T) | Hex | -/Cp | ||
| 3077.58 | 0.03 | 26–57 | 42.18 | (K)SQPAVAPTGDAAAATQVPAGQTVPAQLQFSAK(T) | Wt/Cp | |||
| RV1743 | serine/threonine-protein kinase pknE | 1937.95 | −0.11 | 369–384 | 21.41 | (R)TSAPPPPPPADAVELR(V) | 2 Hex | Wt/- |
| 1613.84 | −2.36 | 369–384 | 22.97 | (R)TSAPPPPPPADAVELR(V) | Wt/Cp | |||
| RV1748 | hypothetical protein | 2889.32 | −1.73 | 55–81 | 30.26 | (R)LGEASGDLASDSPAMPRPTTVENDAAR(W) | Hex | Wt/- |
| 2727.28 | 0.69 | 55–81 | 69.44 | (R)LGEASGDLASDSPAMPRPTTVENDAAR(W) | Wt/Cp | |||
| RV2068c | beta-lactamase | 1778.84 | 0.05 | 33–47 | 13.4 | (R)PASTTLPAGADLADR(F) | 2 Hex | -/Cp |
| 1454.74 | 0.11 | 33–47 | 61.69 | (R)PASTTLPAGADLADR(F) | -/Cp | |||
| RV2744c | phage shock protein A | 1459.71 | −1.44 | 240–253 | 52.1 | (R)GEALPAGGTTATPR(P) | Hex | Wt/Cp |
| 1297.66 | −0.89 | 240–253 | 88.98 | (R)GEALPAGGTTATPR(P) | Wt/Cp | |||
| RV2799 | hypothetical protein | 2082.93 | 2.19 | 68–83 | 43.65 | (K)SPIVATTDPSPFDPCR(D) | 2 Hex, Ca | Wt/Cp |
| RV2969c | hypothetical protein | 2076.05 | −0.45 | 236–255 | 56.32 | (K)IKEIVGDVPGIDSAAATATS(-) | Hex | Wt/- |
| 1913.99 | −0.25 | 236–255 | 64.95 | (K)IKEIVGDVPGIDSAAATATS(-) | Wt/Cp | |||
| 1851.97 | −0.65 | 202–218 | 39.2 | (K)VDGLAAAVNVHATPTVR(V) | Hex | Wt/- | ||
| 1689.91 | −1.01 | 202–218 | 82.14 | (K)VDGLAAAVNVHATPTVR(V) | Wt/- | |||
| RV3090 | hypothetical protein | 2354.19 | 0.16 | 274–295 | 44.23 | (K)GISPLGCWPGSSALPTIAVPGR(-) | Hex, Ca | Wt/Cp |
| 2192.14 | 0.77 | 274–295 | 79.41 | (K)GISPLGCWPGSSALPTIAVPGR(-) | Ca | Wt/Cp | ||
| RV3491 | hypothetical protein | 2022.08 | −0.37 | 164–180 | 38.76 | (K)QPFSLQLIGPPPSPVQR(Y) | Hex | -/Cp |
| 1649.90 | 1.07 | 167–180 | 19.71 | (F)SLQLIGPPPSPVQR(Y) | Hex | Wt/Cp | ||
| 1487.84 | −3.18 | 167–180 | 32.07 | (F)SLQLIGPPPSPVQR(Y) | Wt/- | |||
| RV3584 | lipoprotein lpqE | 1998.96 | 2.22 | 164–182 | 47.69 | (K)AGQGSVMVPISAGLATPHE(-) | Hex, Ox | -/Cp |
| 1982.96 | 0.11 | 164–182 | 56.18 | (K)AGQGSVMVPISAGLATPHE(-) | Hex | Wt/- | ||
| 1820.91 | −0.63 | 164–182 | 110.3 | (K)AGQGSVMVPISAGLATPHE(-) | Wt/Cp | |||
| RV3623 | hypothetical protein | 2219.11 | −0.33 | 195–215 | 64.2 | (K)VISISEASGAAPTHEAPAPPR(G) | Hex | Wt/Cp |
| 2057.06 | 3.52 | 195–215 | 91.34 | (K)VISISEASGAAPTHEAPAPPR(G) | Wt/Cp | |||
| 2454.19 | 0.84 | 99–120 | 28.12 | (R)VTVAPQYSNPEPAGTATITGYR(A) | Hex | Wt/- | ||
| 2292.14 | −0.79 | 99–120 | 93.23 | (R)VTVAPQYSNPEPAGTATITGYR(A) | Wt/- | |||
| RV3763 | lipoprotein lpqH | 3623.49 | 1.18 | 27–51 | 17.12 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 9 Hex | Wt/Cp |
| 3461.43 | 0.06 | 27–51 | 32.84 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 8 Hex | Wt/Cp | ||
| 3299.38 | 1.35 | 27–51 | 38.84 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 7 Hex | Wt/Cp | ||
| 3137.33 | −0.11 | 27–51 | 19.27 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 6 Hex | Wt/Cp | ||
| 2813.22 | 0.64 | 27–51 | 31.75 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 4 Hex | Wt/Cp | ||
| 2651.17 | 1.27 | 27–51 | 51.82 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 3 Hex | Wt/Cp | ||
| 2489.12 | 1.19 | 27–51 | 69.7 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | 2 Hex | Wt/Cp | ||
| 2327.06 | 1.42 | 27–51 | 72.66 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | Hex | -/Cp | ||
| 2165.00 | −1.59 | 27–51 | 111.1 | (K)STTGSGETTTAAGTTASPGAASGPK(V) | Wt/Cp | |||
| RV3835 | hypothetical protein | 2030.98 | 0.87 | 72–87 | 13.38 | (R)LAGYIASNPVPSTGAK(I) | 3 Hex | -/Cp |
| 1706.87 | 0.35 | 72–87 | 26.07 | (R)LAGYIASNPVPSTGAK(I) | Hex | -/Cp | ||
| RV3869 | ESX-1 secretion system protein eccB1 | 1978.95 | 0.12 | 454–470 | 43.68 | (K)DAALLEHDTLPADPSPR(K) | Hex | Wt/Cp |
| 1816.89 | −1.10 | 454–470 | 60.59 | (K)DAALLEHDTLPADPSPR(K) | Wt/Cp |
| Gene | prot_desc | pep_exp_mr | start-pep_end | pep_score | pep_seq | pep_var_mod |
|---|---|---|---|---|---|---|
| RV0822c | hypothetical protein | 1637.79 | 522-534 | 16.01 | LGNTPSTPPTTTK | 2 Hex |
| 1475.74 | 522-534 | 35.88 | LGNTPSTPPTTTK | Hex | ||
| 1313.68 | 522-534 | 32.55 | LGNTPSTPPTTTK | |||
| RV1157c | hypothetical protein | 1294.64 | 211-224 | 23.21 | AAAPAPASAAPAPA | Hex |
| 1132.59 | 211-224 | 22.01 | AAAPAPASAAPAPA | |||
| 1365.68 | 211-225 | 10.82 | AAAPAPASAAPAPAA | Hex | ||
| RV2094c | TatA/E family twin arginine-targeting protein translocase | 1503.74 | 55-66 | 22.44 | SIETPTPVQSQR | Hex |
| 1341.69 | 55-66 | 52.68 | SIETPTPVQSQR | |||
| RV2220 | glutamine synthetase 1 | 1340.63 | 2-11 | 21.43 | TEKTPDDVFK | Hex |
| 1178.58 | 2-11 | 55.91 | TEKTPDDVFK | |||
| RV3244c | lipoprotein lpqB | 3156.58 | 29-53 | 30.03 | GTVERPVPSNLPKPSPGMDPDVLLR | 3 Hex |
| 2994.52 | 29-53 | 22.99 | GTVERPVPSNLPKPSPGMDPDVLLR | 2 Hex | ||
| RV3413c | hypothetical protein | 1646.78 | 285-299 | 30.05 | PAGQPAPETPVSPTH | Hex |
| 1484.73 | 285-299 | 50.33 | PAGQPAPETPVSPTH | |||
| RV3705A | hypothetical protein | 1860.89 | 115-129 | 15.18 | VGPTGPGPTTAPARP | 3 Hex |
| 1698.83 | 115-129 | 30.6 | VGPTGPGPTTAPARP | 2 Hex | ||
| RV3706c | hypothetical protein | 2981.43 | 81-106 | 8.08 | AVRPGPGPGGPGQVPSSVSPPATPAP | 4 Hex |
| 2819.37 | 81-106 | 25.94 | AVRPGPGPGGPGQVPSSVSPPATPAP | 3 Hex | ||
| 2657.32 | 81-106 | 26.69 | AVRPGPGPGGPGQVPSSVSPPATPAP | 2 Hex | ||
| 2910.39 | 82-106 | 16.99 | VRPGPGPGGPGQVPSSVSPPATPAP | 4 Hex | ||
| 2748.34 | 82-106 | 20.4 | VRPGPGPGGPGQVPSSVSPPATPAP | 3 Hex | ||
| 2586.28 | 82-106 | 25.37 | VRPGPGPGGPGQVPSSVSPPATPAP | 2 Hex |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonini, L.; Sadet, B.; Stella, A.; Bouyssié, D.; Nigou, J.; Burlet-Schiltz, O.; Rivière, M. Potential Plasticity of the Mannoprotein Repertoire Associated to Mycobacterium tuberculosis Virulence Unveiled by Mass Spectrometry-Based Glycoproteomics. Molecules 2020, 25, 2348. https://doi.org/10.3390/molecules25102348
Tonini L, Sadet B, Stella A, Bouyssié D, Nigou J, Burlet-Schiltz O, Rivière M. Potential Plasticity of the Mannoprotein Repertoire Associated to Mycobacterium tuberculosis Virulence Unveiled by Mass Spectrometry-Based Glycoproteomics. Molecules. 2020; 25(10):2348. https://doi.org/10.3390/molecules25102348
Chicago/Turabian StyleTonini, Laure, Bashir Sadet, Alexandre Stella, David Bouyssié, Jérôme Nigou, Odile Burlet-Schiltz, and Michel Rivière. 2020. "Potential Plasticity of the Mannoprotein Repertoire Associated to Mycobacterium tuberculosis Virulence Unveiled by Mass Spectrometry-Based Glycoproteomics" Molecules 25, no. 10: 2348. https://doi.org/10.3390/molecules25102348
APA StyleTonini, L., Sadet, B., Stella, A., Bouyssié, D., Nigou, J., Burlet-Schiltz, O., & Rivière, M. (2020). Potential Plasticity of the Mannoprotein Repertoire Associated to Mycobacterium tuberculosis Virulence Unveiled by Mass Spectrometry-Based Glycoproteomics. Molecules, 25(10), 2348. https://doi.org/10.3390/molecules25102348