Exogenous Melatonin and Abscisic Acid Expedite the Flavonoids Biosynthesis in Grape Berry of Vitis vinifera cv. Kyoho




Abstract

1. Introduction
2. Results
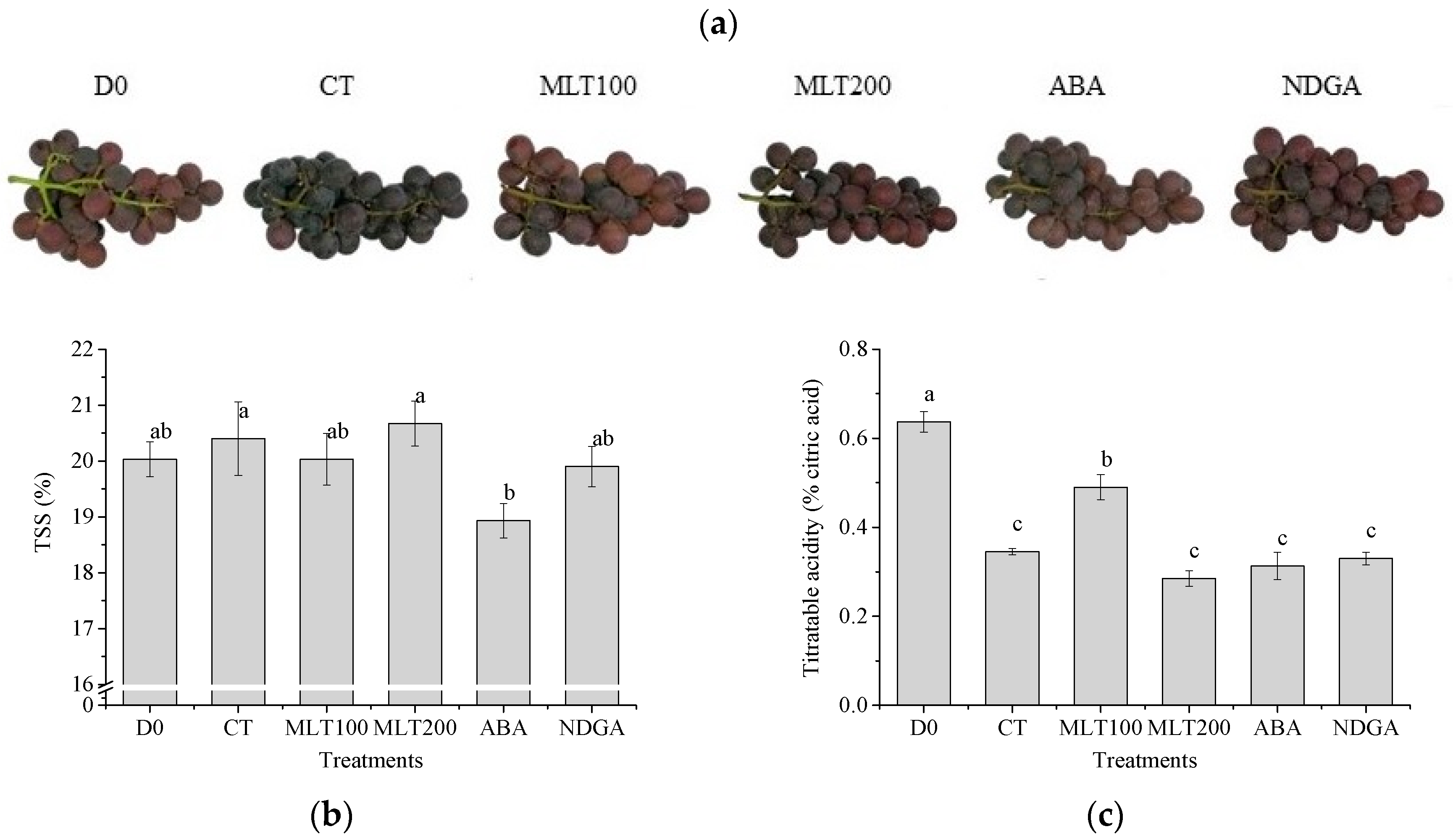
2.1. Grape Morphology and Berry TSS, TA Concentrations
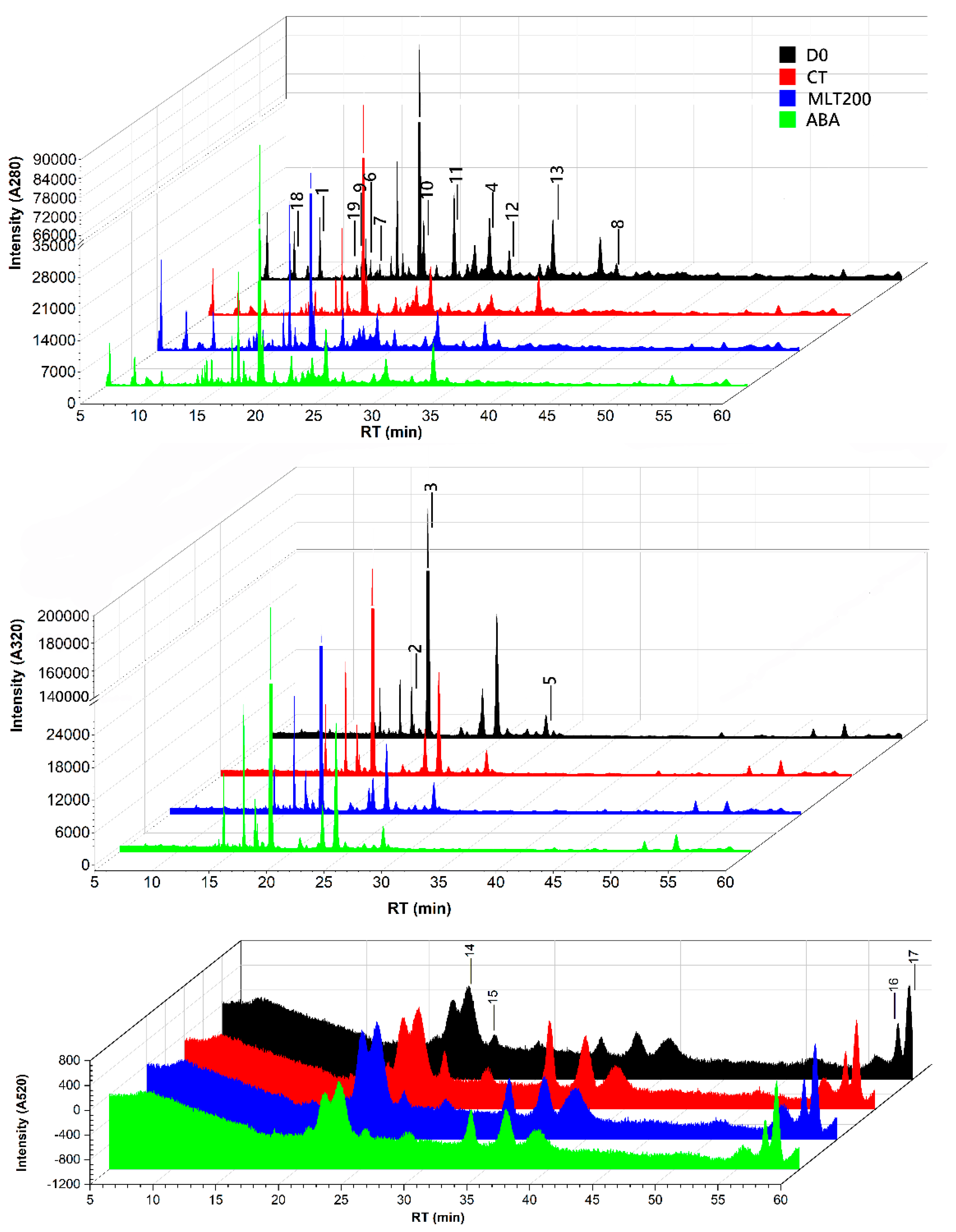
2.2. Polyphenolic Profiles
2.3. Total Phenolic Acid, Flavonol, Flavanol, and Anthocyanin Contents
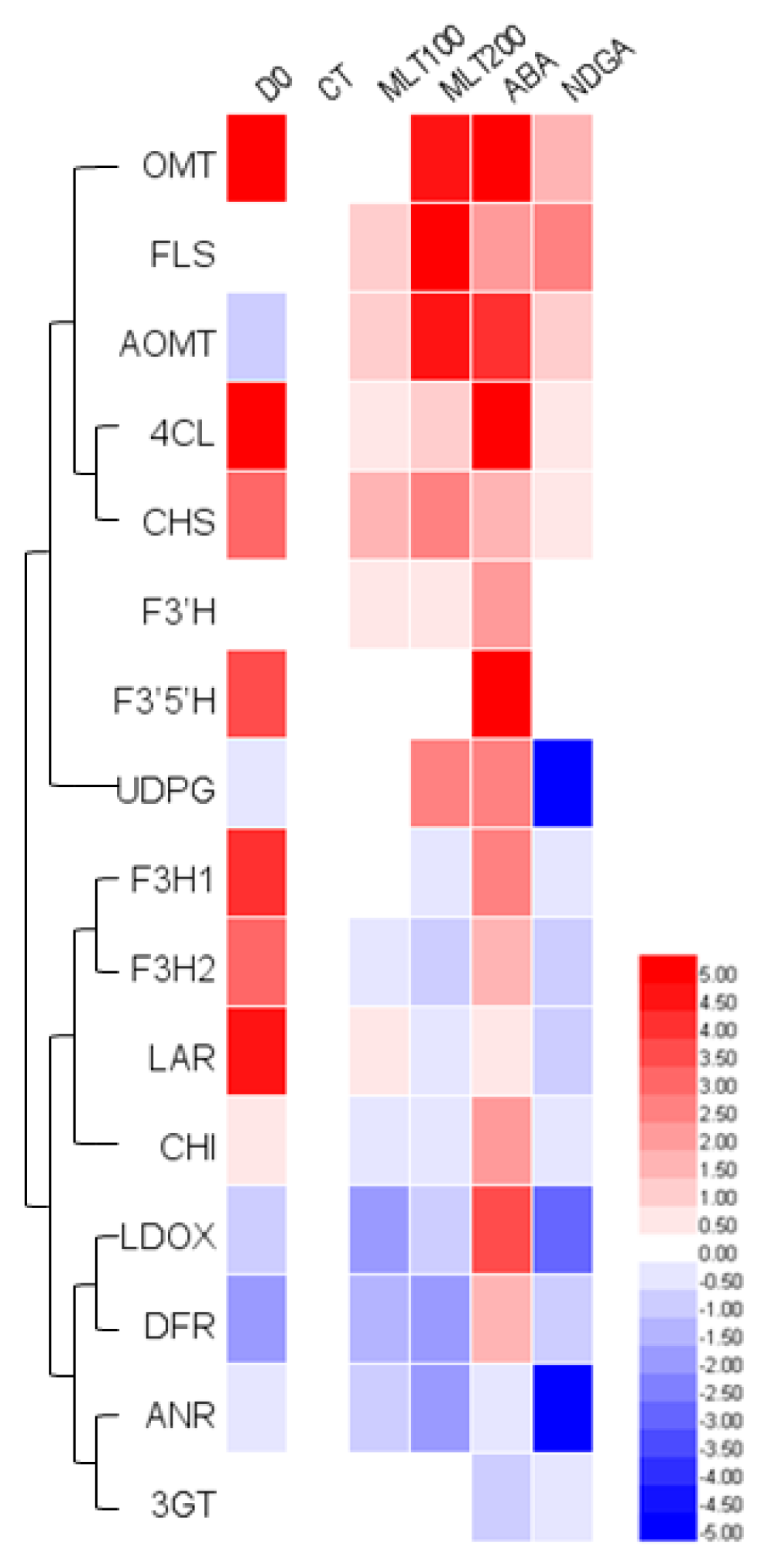
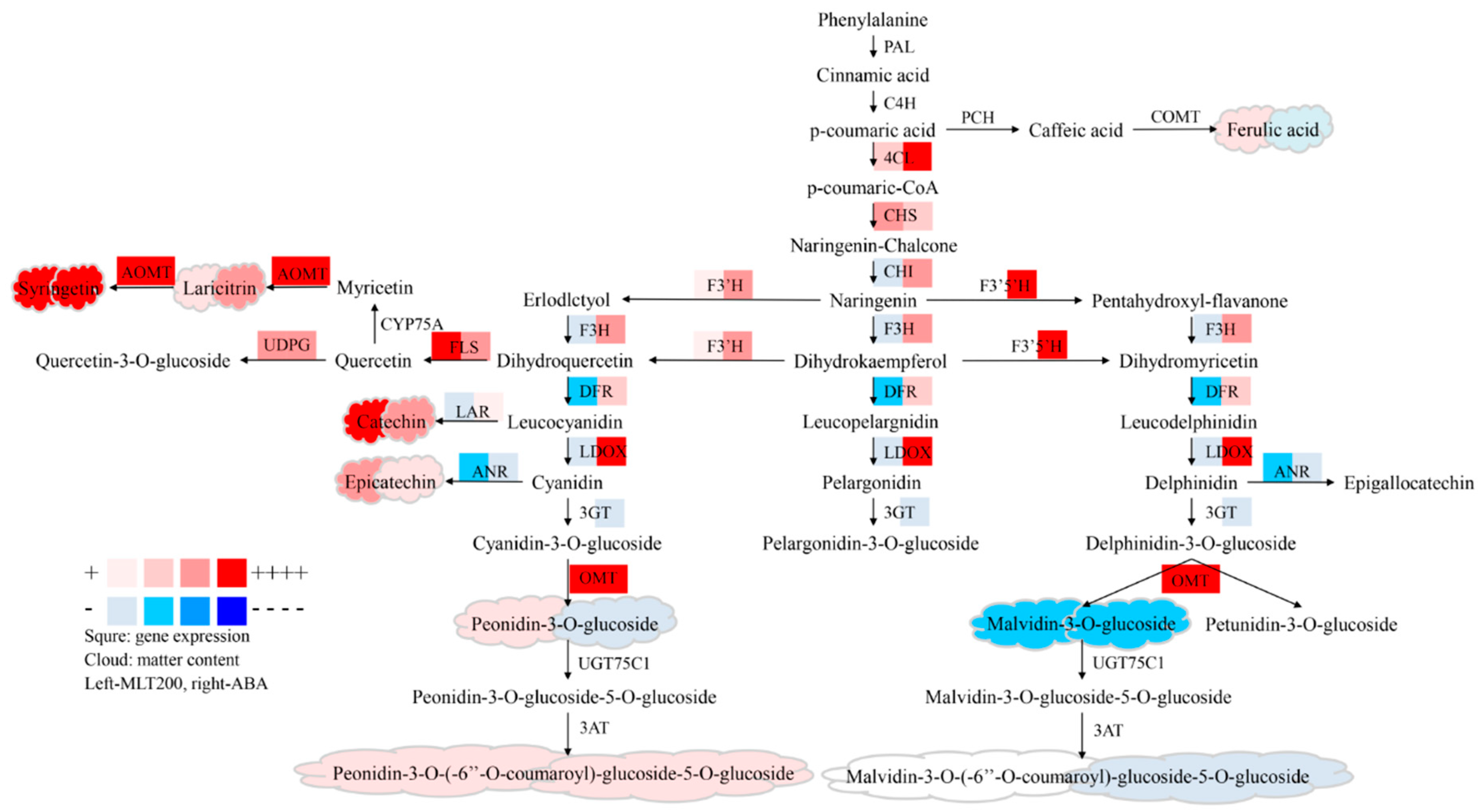
2.4. Expression of Genes Involved in Polyphenolic Biosynthesis
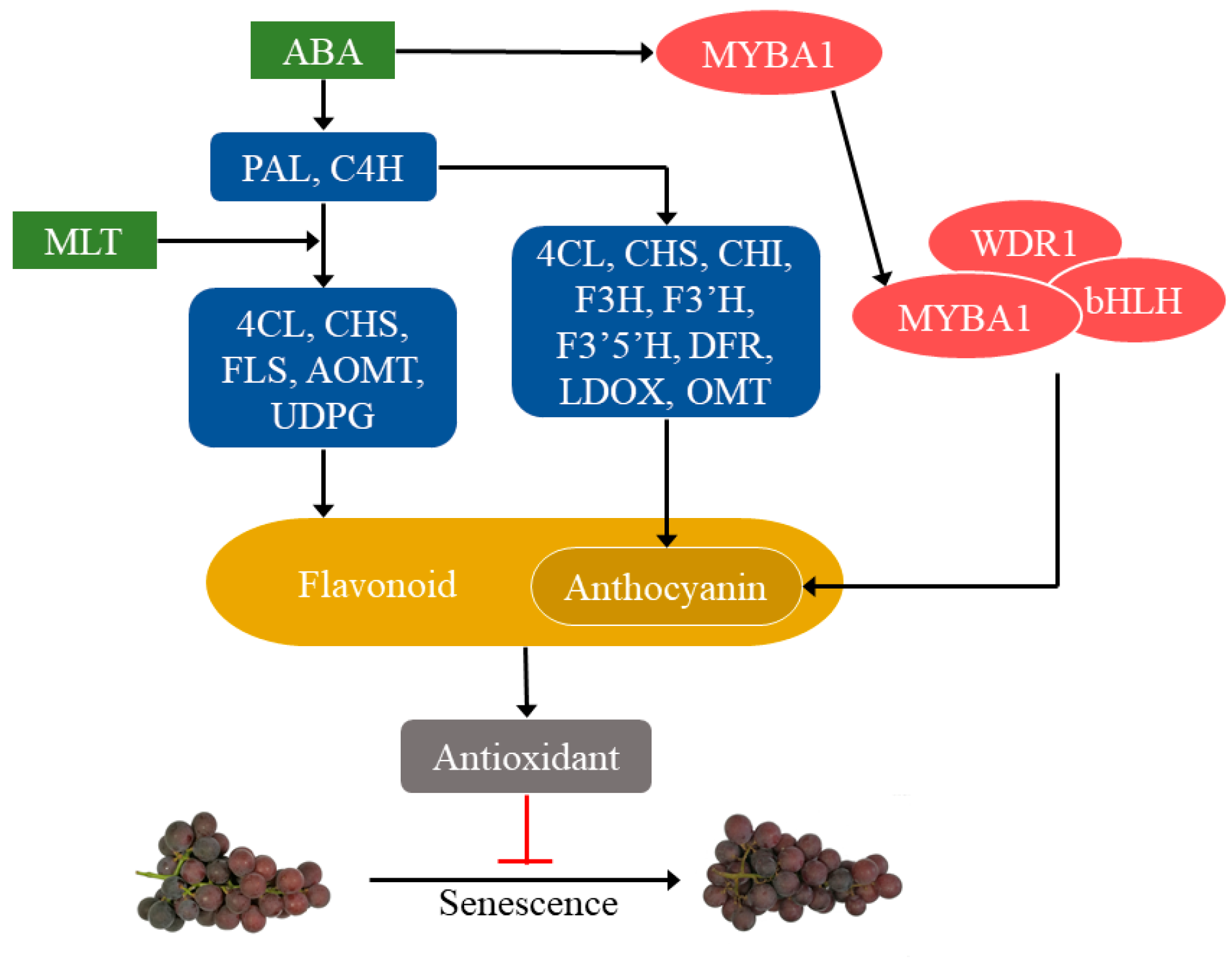
3. Discussion
4. Materials and Methods
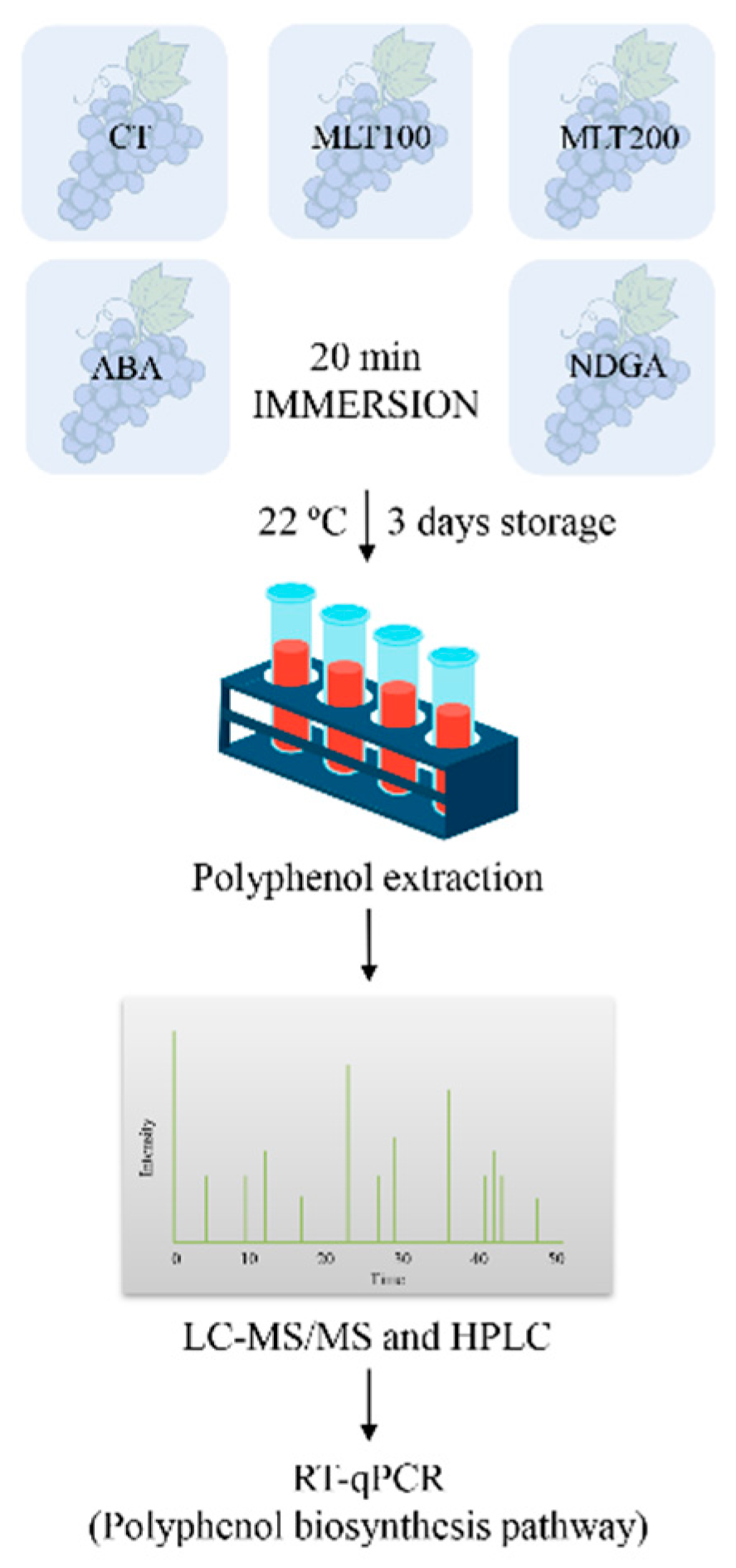
4.1. Plant Materials and Experimental Design
4.2. Total Soluble Solids (TSS) and Titratable Acidity (TA) Determination
4.3. Polyphenols Extraction and Separation
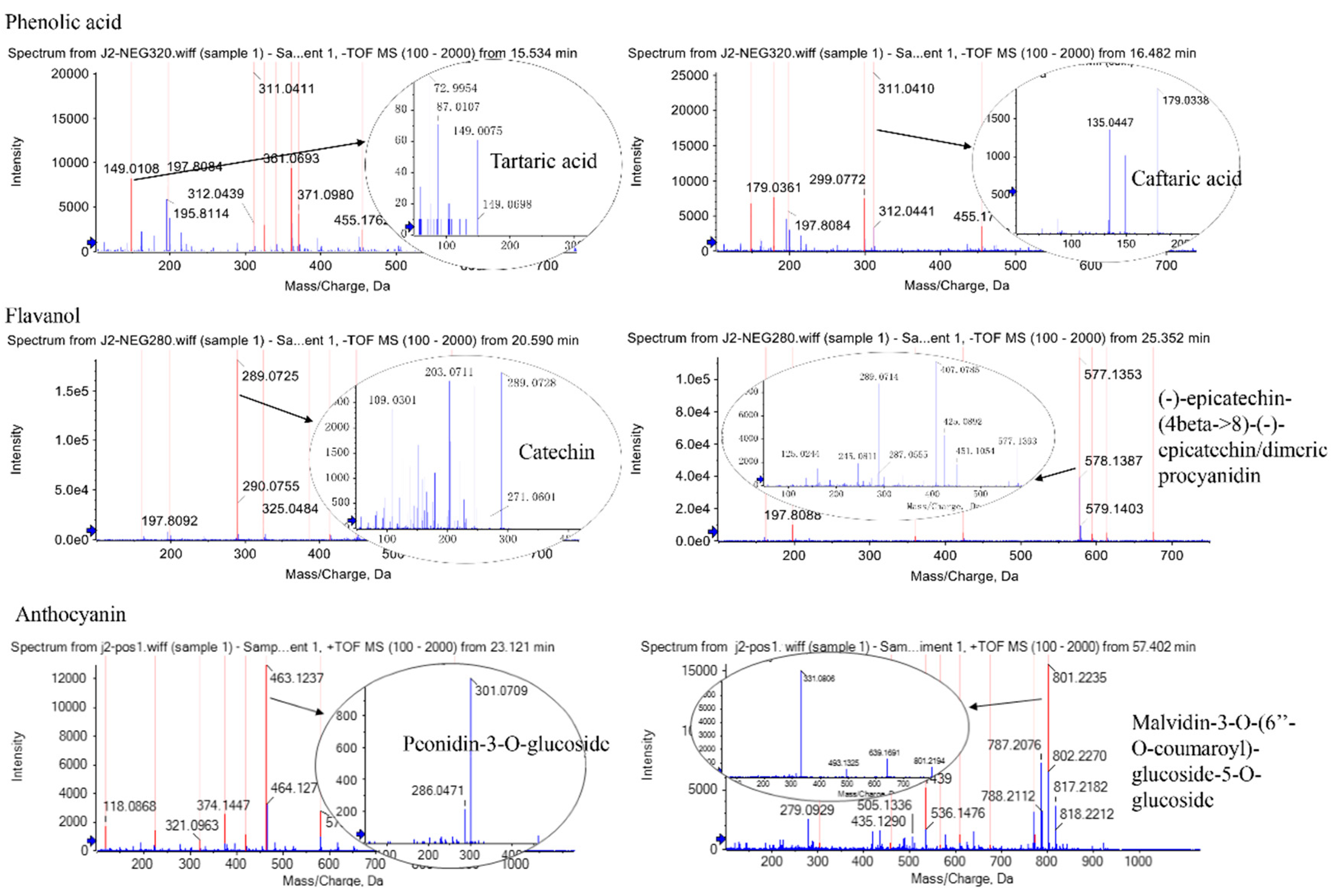
4.4. Identification and Quantification of Polyphenols by HPLC and HPLC-Q-TOF-MS
4.5. Real-Time Quantitative PCR (RT-qPCR)
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PAL | phenylalanine ammonia-lyase |
| C4H | cinnamate-4-hydroxylase |
| PCH | p-coumarate 3-hydroxylase |
| COMT | caffeic acid 3-O-methyltransferase |
| 4CL | 4-coumaroyl-CoA synthase |
| CHS | chalcone synthase |
| CHI | chalcone isomerase |
| F3′H | flavonoid 3′-hydroxylase |
| F3′5′H | flavonoid 3′5′-hydroxylase |
| F3H | flavanone 3β-hydroxylase |
| DFR | dihydroflavonol 4-reductase |
| LDOX | leucoanthocyanidin dioxygenase |
| 3GT | anthocyanidin 3-O-glucosyltransferase |
| OMT | anthocyanin 3′-methyltransferase |
| UGT75C1 | anthocyanin 5-O-glucosyltransferase |
| 3AT | anthocyanidin 3-O-glucoside 6″-O-acyltransferase |
| CYP75A | flavonoid 3′,5′-hydroxylase |
| AOMT | flavonoid 3′,5′-methyltransferase |
| FLS | flavonol synthase |
| UDPG | flavonoid-3-O-glucosyltransferase |
| LAR | leucoanthocyanidin reductase |
| ANR | anthocyanidin reductase |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
|---|---|---|
| 4CL | ACCACCTCCCTCTCCACAC | GCTCCGAGAAAGGAGAACG |
| CHS | ACCCACCTGGATTCTCTCG | GAAGGCTTCCACCAAGCTC |
| CHI | TTGTGTTGGTTCCTCTTGTTC | GCAGACGAATCTCATTCAGTATAG |
| F3H1 | CCAATCATAGCAGACTGTCC | TCAGAGGATACACGGTTGCC |
| F3H2 | CTGTGGTGAACTCCGACTGC | CAAATGTTATGGGCTCCTCC |
| F3′H | GAGATCAACGGCTACCACATC | CCTGAATTCTAGTGGCTTCTCC |
| F3′5′H | GCTGGCACTAGAATGGGAATAG | CTCAACTCCATCCGGCATTT |
| DFR | TTGTAATGGTCAATGTGCC | CATGCAGAGACCACCTTG |
| LDOX | ACCTTCATCCTCCACAACAT | AGTAGAGCCTCCTGGGTCTT |
| 3GT | TCATGAAGACCAGACCCTTA | TGTTGCTACTACGGGGTCTA |
| OMT | AGATAAAGAAGCGTACCAAA | GCTCATGGTAGTTGAGGTAG |
| LAR | CGGTTGGTAGCATTAAGAGGTTC | GGAGTTGCAGCAGATGTAGGTGT |
| ANR | TTGATGGGACAGGTCTGGTTG | AGTGTCTTGGAGGCAGGATAGC |
| FLS | CCAGCCAATCCTTCAAACTT | TTTGCCATTTGCATTGTGTA |
| UDPG | TGCTACCTAAGGCGACTG | GCTTGGATTTGAGATCATTGG |
| AOMT | AGGTGTGGAGCATAAGATCA | TTATCGTAAGCGATTATGCC |
- Polyphenols Market Projected to Reach $1.33 Billion by 2024. Available online: http://www.abnewswire.com/pressreleases/polyphenols-market-projected-to-reach-133-billion-by-2024-grand-view-research-inc_75694.html (accessed on 11 November 2019).
- Shahidi, F.; Yeo, J. Bioactivities of phenolics by focusing on suppression of chronic diseases: A review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef] [PubMed]
- Distribution of the World’s Grapevine Varieties. Available online: https://www.amorimcorkitalia.com/media/en-distribution-of-the-worlds-3f7.pdf (accessed on 11 November 2019).
- Vantassel, D.; Roberts, N.; Oenill, S. Melatonin from higher-plants-isolation and identification of N-acetyl 5-methoxytryptamine. Plant Physiol. 1995, 108, 101. [Google Scholar]
- Zhai, R.; Liu, J.; Liu, F.; Zhao, Y.; Liu, L.; Fang, C.; Wang, H.; Li, X.; Wang, Z.; Ma, F.; et al. Melatonin limited ethylene production, softening and reduced physiology disorder in pear (Pyrus communis L.) fruit during senescence. Postharvest Biol. Technol. 2018, 139, 38–46. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of melatonin treatment on the postharvest quality of strawberry fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Z.K.; Chai, H.K.; Cheng, N.; Yang, Y.; Wang, D.N.; Yang, T.; Cao, W. Melatonin treatment delays postharvest senescence and regulates reactive oxygen species metabolism in peach fruit. Postharvest Biol. Technol. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Hu, W.; Yang, H.; Tie, W.; Yan, Y.; Ding, Z.; Liu, Y.; Wu, C.; Wang, J.; Reiter, R.J.; Tan, D.X.; et al. Natural variation in banana varieties highlights the role of melatonin in postharvest ripening and quality. J. Agric. Food Chem. 2017, 65, 9987–9994. [Google Scholar] [CrossRef]
- Iriti, M.; Varoni, E.M. The good health of Bacchus: Melatonin in grapes, the unveiled myth. LWT-Food Sci. Technol. 2016, 65, 758–761. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2015, 66, 657–668. [Google Scholar] [CrossRef]
- Zhang, Y.; Huber, D.J.; Hu, M.; Jiang, G.; Gao, Z.; Xu, X.; Jiang, Y.; Zhang, Z. Delay of postharvest browning in litchi fruit by melatonin via the enhancing of antioxidative processes and oxidation repair. J. Agric. Food Chem. 2018, 66, 7475–7484. [Google Scholar] [CrossRef]
- Xu, L.; Yue, Q.; Bian, F.; Sun, H.; Zhai, H.; Yao, Y. Melatonin enhances phenolics accumulation partially via ethylene signaling and resulted in high antioxidant capacity in grape berries. Front. Plant Sci. 2017, 8, 1426. [Google Scholar] [CrossRef]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. Apple bZIP transcription factor MdbZIP44 regulates abscisic acid-promoted anthocyanin accumulation. Plant Cell Environ. 2018, 41, 2678–2692. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yin, W.; Wu, J.; Chai, L.; Yi, H. Effects of exogenous abscisic acid on the expression of citrus fruit ripening-related genes and fruit ripening. Sci. Hortic. 2016, 201, 175–183. [Google Scholar] [CrossRef]
- Villalobos-Gonzalez, L.; Pena-Neira, A.; Ibanez, F.; Pastenes, C. Long-term effects of abscisic acid (ABA) on the grape berry phenylpropanoid pathway: Gene expression and metabolite content. Plant Physiol. Biochem. 2016, 105, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Li, J.; Wang, D.; Wang, H.; Qin, Y.; Hu, G.; Zhao, J. Transcriptome profiling of Litchi chinensis pericarp in response to exogenous cytokinins and abscisic acid. Plant Growth Regul. 2017, 84, 437–450. [Google Scholar] [CrossRef]
- Li, D.; Li, L.; Luo, Z.; Mou, W.; Mao, L.; Ying, T. Comparative transcriptome analysis reveals the influence of abscisic acid on the metabolism of pigments, ascorbic acid and folic acid during strawberry fruit ripening. PLoS ONE 2015, 10, e0130037. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.; Li, D.; Luo, Z.; Mao, L.; Ying, T. Transcriptomic analysis reveals possible influences of ABA on secondary metabolism of pigments, flavonoids and antioxidants in tomato fruit during ripening. PLoS ONE 2015, 10, e0129598. [Google Scholar] [CrossRef]
- Shen, X.; Zhao, K.; Liu, L.; Zhang, K.; Yuan, H.; Liao, X.; Wang, Q.; Guo, X.; Li, F.; Li, T. A role for PacMYBA in ABA-regulated anthocyanin biosynthesis in red-colored sweet cherry cv. Hong Deng (Prunus avium L.). Plant Cell Physiol. 2014, 55, 862–880. [Google Scholar] [CrossRef]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Takahashi, A.; Masuda, T.; Noji, S. Light and abscisic acid independently regulated FaMYB10 in Fragaria x ananassa fruit. Planta 2015, 241, 953–965. [Google Scholar] [CrossRef]
- Fontana, A.R.; Antoniolli, A.; Bottini, R. Grape pomace as a sustainable source of bioactive compounds: Extraction, characterization, and biotechnological applications of phenolics. J. Agric. Food Chem. 2013, 61, 8987–9003. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, X.; Yang, Q.; Zhao, Q. Exogenous melatonin delays postharvest fruit senescence and maintains the quality of sweet cherries. Food Chem. 2019, 301, 125311. [Google Scholar] [CrossRef]
- Meng, J.F.; Xu, T.F.; Song, C.Z.; Yu, Y.; Hu, F.; Zhang, L.; Zhang, Z.W.; Xi, Z.M. Melatonin treatment of pre-veraison grape berries to increase size and synchronicity of berries and modify wine aroma components. Food Chem. 2015, 185, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, G.; Temsamani, H.; Bordenave, L.; Pedrot, E.; Chaher, N.; Cluzet, S.; Delaunay, J.C.; Ollat, N.; Delrot, S.; Merillon, J.M.; et al. Flavonol profiles in berries of wild Vitis accessions using liquid chromatography coupled to mass spectrometry and nuclear magnetic resonance spectrometry. Food Chem. 2015, 169, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.F.; Yu, Y.; Shi, T.C.; Fu, Y.S.; Zhao, T.; Zhang, Z.W. Melatonin treatment of pre-veraison grape berries modifies phenolic components and antioxidant activity of grapes and wine. Food Sci. Technol. 2019, 39, 35–42. [Google Scholar] [CrossRef]
- Debnath, B.; Hussain, M.; Li, M.; Lu, X.; Sun, Y.; Qiu, D. Exogenous melatonin improves fruit quality features, health promoting antioxidant compounds and yield traits in tomato fruits under acid rain stress. Molecules 2018, 23, 1868. [Google Scholar] [CrossRef]
- Wu, Q.; Bai, J.; Tao, X.; Mou, W.; Luo, Z.; Mao, L.; Ban, Z.; Ying, T.; Li, L. Synergistic effect of abscisic acid and ethylene on color development in tomato (Solanum lycopersicum L.) fruit. Sci. Hortic. 2018, 235, 169–180. [Google Scholar] [CrossRef]
- Olivares, D.; Contreras, C.; Munoz, V.; Rivera, S.; Gonzalez-Aguero, M.; Retamales, J.; Defilippi, B.G. Relationship among color development, anthocyanin and pigment-related gene expression in ‘Crimson Seedless’ grapes treated with abscisic acid and sucrose. Plant Physiol. Biochem. 2017, 115, 286–297. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y.; He, C.; Zhu, S. Postharvest exogenous application of abscisic acid reduces internal browning in pineapple. J. Agric. Food Chem. 2015, 63, 5313–5320. [Google Scholar] [CrossRef]
- Gao, Z.; Li, Q.; Li, J.; Chen, Y.; Luo, M.; Li, H.; Wang, J.; Wu, Y.; Duan, S.; Wang, L.; et al. Characterization of the ABA receptor VlPYL1 that regulates anthocyanin accumulation in grape berry skin. Front. Plant Sci. 2018, 9, 592. [Google Scholar] [CrossRef]
- Shin, D.H.; Cho, M.; Choi, M.G.; Das, P.K.; Lee, S.K.; Choi, S.B.; Park, Y.I. Identification of genes that may regulate the expression of the transcription factor production of anthocyanin pigment 1 (PAP1)/MYB75 involved in Arabidopsis anthocyanin biosynthesis. Plant Cell Rep. 2015, 34, 805–815. [Google Scholar] [CrossRef]
- Wei, J.; Li, D.X.; Zhang, J.R.; Shan, C.; Rengel, Z.; Song, Z.B.; Chen, Q. Phytomelatonin receptor PMTR1-mediated signaling regulates stomatal closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef]
- Xu, L.; Yue, Q.; Xiang, G.; Bian, F.; Yao, Y. Melatonin promotes ripening of grape berry via increasing the levels of ABA, H2O2, and particularly ethylene. Hortic. Res. 2018, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Lu, Z.; Yang, Y.; Wang, D.; Yang, T.; Cao, M.; Cao, W. Melatonin treatment reduces chilling injury in peach fruit through its regulation of membrane fatty acid contents and phenolic metabolism. Food Chem. 2018, 245, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Matus, J.T.; Poupin, M.J.; Canon, P.; Bordeu, E.; Alcalde, J.A.; Arce-Johnson, P. Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.). Plant Mol. Biol. 2010, 72, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HemI: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







| Peak No. | Compound | Spectrum (nm) | RT (min) | Mw (Da) | Productions | D0 | CT | MLT100 | MLT200 | ABA | NDGA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Phenolic acids | |||||||||||
| 1 | Gallic acid | 280 | 9.80 | 170 | 125 | 0.85 ± 0.02 c | 0.45 ± 0.09 e | 0.65 ± 0.03 d | 0.94 ± 0.03 b | 0.68 ± 0.03 d | 1.22 ± 0.00 a |
| 2 | Caftaric acid | 320 | 18.15 | 312 | 179,149,135 | 611.56 ± 4.70 a | 538.68 ± 17.25 b | 488.60 ± 33.41 bc | 511.59 ± 8.23 c | 602.93 ± 40.67 a | 402.73 ± 2.43 d |
| 3 | Coutaric acid | 280 | 23.86 | 296 | 163 | 3.56 ± 0.04 a | 2.83 ± 0.12 c | 2.29 ± 0.07 d | 2.96 ± 0.04 c | 3.06 ± 0.03 b | 2.08 ± 0.01 e |
| 4 | Fertaric acid | 320 | 27.99 | 326 | 193,134 | 21.28 ± 0.16 b | 20.74 ± 0.43 c | 17.71 ± 0.20 e | 24.98 ± 0.15 a | 19.67 ± 0.23 d | 20.50 ± 0.16 c |
| Total phenolic acid content | 637.25 ± 4.92 a | 562.70 ± 17.89 b | 509.25 ± 33.71 c | 540.47 ± 8.45 bc | 626.34 ± 40.96 a | 426.53 ± 2.60 d | |||||
| Flavonols | |||||||||||
| 5 | Laricitrin | 280 | 13.65 | 332 | 169,125 | 3.08 ± 0.07 c | 1.96 ± 0.32 e | 3.46 ± 0.22 b | 2.71 ± 0.01 d | 3.58 ± 0.54 bc | 4.41 ± 0.16 a |
| 6 | Syringetin | 280 | 14.88 | 346 | 183 | 1.50 ± 0.00 a | - | - | 1.16 ± 0.00 b | 0.73 ± 0.00 d | 0.88 ± 0.00 c |
| 7 | Kaempferol 3-O-glucoronide | 280 | 34.27 | 462 | 415,311,149 | 1.94 ± 0.22 a | 1.38 ± 0.04 b | 1.34 ± 0.10 b | 1.97 ± 0.12 a | 1.50 ± 0.12 b | 2.16 ± 0.10 a |
| Total flavonol content | 6.52 ± 0.29 b | 3.34 ± 1.73 e | 4.80 ± 0.32 d | 5.84 ± 0.13 c | 5.81 ± 0.66 bc | 7.45 ± 0.26 a | |||||
| Flavanols | |||||||||||
| 8 | Proanthocyanidin trimer | 280 | 13.26 | 866 | 866,865,695,575,451,407,287,243,125 | 2.43 ± 0.05 b | 1.34 ± 0.01 f | 1.38 ± 0.03 e | 2.01 ± 0.01 c | 1.45 ± 0.01 d | 2.96 ± 0.01 a |
| 9 | (-)-epicatechin-(4beta->8)-(-)-epicatechin/dimeric procyanidin | 280 | 18.42 | 578 | 425,407,289,245 | 10.94 ± 0.77 a | 6.05 ± 0.58 d | 5.42 ± 0.36 d | 8.25 ± 0.51 b | 8.32 ± 0.00 b | 7.23 ± 0.16 c |
| 10 | Catechin | 280 | 20.91 | 290 | 245 | 13.57 ± 0.29 a | 4.20 ± 0.02 f | 5.60 ± 0.30 e | 8.24 ± 0.15 c | 7.13 ± 0.28 d | 8.64 ± 0.18 b |
| 11 | (-)-epicatechin-(4beta->8)-(-)-epicatechin/dimeric procyanidin | 280 | 25.34 | 578 | 425,407,289,125 | 4.36 ± 0.09 b | 2.37 ± 0.18 e | 2.89 ± 0.29 d | 4.06 ± 0.09 c | 2.78 ± 0.12 d | 5.16 ± 0.40 a |
| 12 | Epicatechin | 280 | 29.02 | 290 | 245,203,109 | 13.36 ± 0.62 a | 6.67 ± 0.21 e | 8.15 ± 0.48 d | 10.24 ± 0.49 c | 8.18 ± 0.26 d | 11.62 ± 0.43 b |
| Total flavanol content | 44.66 ± 1.82 a | 20.63 ± 1.00 f | 23.44 ± 1.46 e | 32.80 ± 1.25 c | 28.36 ± 0.67 d | 35.61 ± 1.18 b | |||||
| Anthocyanins | |||||||||||
| 13 | Peonidin 3-O-glucoside | 520 | 23.35 | 463 | 301 | 4.27 ± 0.06 d | 5.52 ± 0.10 b | 3.19 ± 0.25 f | 6.10 ± 0.07 a | 4.57 ± 0.09 c | 3.62 ± 0.03 e |
| 14 | Malvidin 3-O-glucoside | 520 | 25.50 | 493 | 331 | 0.32 ± 0.18 c | 0.83 ± 0.11 a | 0.48 ± 0.05 bc | 0.57 ± 0.05 b | 0.57 ± 0.04 b | 0.45 ± 0.04 c |
| 15 | Malvidin 3-O-(6″-O-coumaroyl)-glucoside-5-O-glucoside | 520 | 57.32 | 801 | 639,493,331 | 0.99 ± 0.01 b | 1.00 ± 0.04 ab | 0.55 ± 0.02 d | 1.03 ± 0.02 a | 0.84 ± 0.00 c | 1.03 ± 0.06 ab |
| 16 | Peonidin 3-O-(6″-O-coumaroyl)-glucoside-5-O-glucoside | 520 | 58.19 | 771 | 609,463,301 | 2.67 ± 0.06 a | 2.13 ± 0.00 c | 1.20 ± 0.05 e | 2.35 ± 0.11 b | 2.22 ± 0.04 b | 2.01 ± 0.02 d |
| Total anthocyanin content | 8.25 ± 0.31 c | 9.48 ± 0.25 b | 5.42 ± 0.37 e | 10.05 ± 0.25 a | 8.20 ± 0.17 c | 7.11 ± 0.15 d | |||||
| Unidentified | |||||||||||
| 17 | Unknown | 280 | 7.49 | 370 | 207 | 6.50 ± 0.01 a | 4.49 ± 0.19 d | 4.58 ± 0.09 d | 6.19 ± 0.12 b | 4.82 ± 0.08 c | 6.44 ± 0.04 a |
| 18 | Unknown | 280 | 12.86 | 452 | 323,89 | 2.52 ± 0.21 a | 1.36 ± 0.05 d | 1.28 ± 0.09 d | 2.02 ± 0.07 b | 1.79 ± 0.01 c | 1.69 ± 0.18 c |
| Total polyphenol content | 705.70 ± 7.56 a | 602.00 ± 21.11 b | 548.77 ± 36.04 c | 597.37 ± 10.27 b | 675.32 ± 42.55 a | 484.83 ± 4.41 d | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Wang, L.; Belwal, T.; Zhang, X.; Lu, H.; Chen, C.; Li, L. Exogenous Melatonin and Abscisic Acid Expedite the Flavonoids Biosynthesis in Grape Berry of Vitis vinifera cv. Kyoho. Molecules 2020, 25, 12. https://doi.org/10.3390/molecules25010012
Yang M, Wang L, Belwal T, Zhang X, Lu H, Chen C, Li L. Exogenous Melatonin and Abscisic Acid Expedite the Flavonoids Biosynthesis in Grape Berry of Vitis vinifera cv. Kyoho. Molecules. 2020; 25(1):12. https://doi.org/10.3390/molecules25010012
Chicago/Turabian StyleYang, Mingyi, Lei Wang, Tarun Belwal, Xiaocheng Zhang, Hongyan Lu, Cunkun Chen, and Li Li. 2020. "Exogenous Melatonin and Abscisic Acid Expedite the Flavonoids Biosynthesis in Grape Berry of Vitis vinifera cv. Kyoho" Molecules 25, no. 1: 12. https://doi.org/10.3390/molecules25010012
APA StyleYang, M., Wang, L., Belwal, T., Zhang, X., Lu, H., Chen, C., & Li, L. (2020). Exogenous Melatonin and Abscisic Acid Expedite the Flavonoids Biosynthesis in Grape Berry of Vitis vinifera cv. Kyoho. Molecules, 25(1), 12. https://doi.org/10.3390/molecules25010012