
Discerning between Two Tuscany (Italy) Ancient Apple cultivars, ‘Rotella’ and ‘Casciana’, through Polyphenolic Fingerprint and Molecular Markers


 , , ,
, , , 
Abstract
1. Introduction
2. Results and Discussion
2.1. Pomological and Organoleptic Characteristics
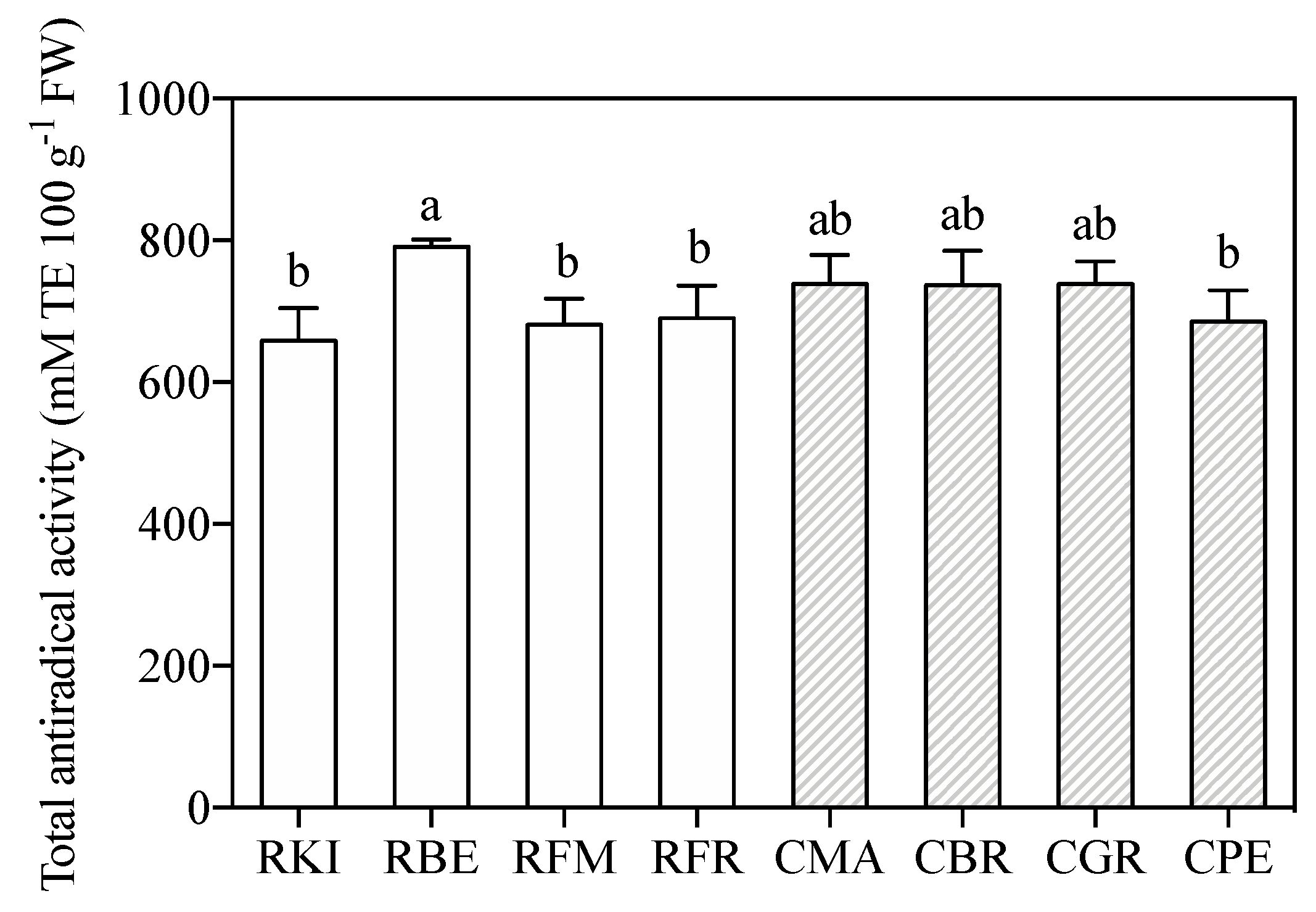
2.2. Polyphenolic Profile, Total Antiradical Activity and Nutraceutical Attributes
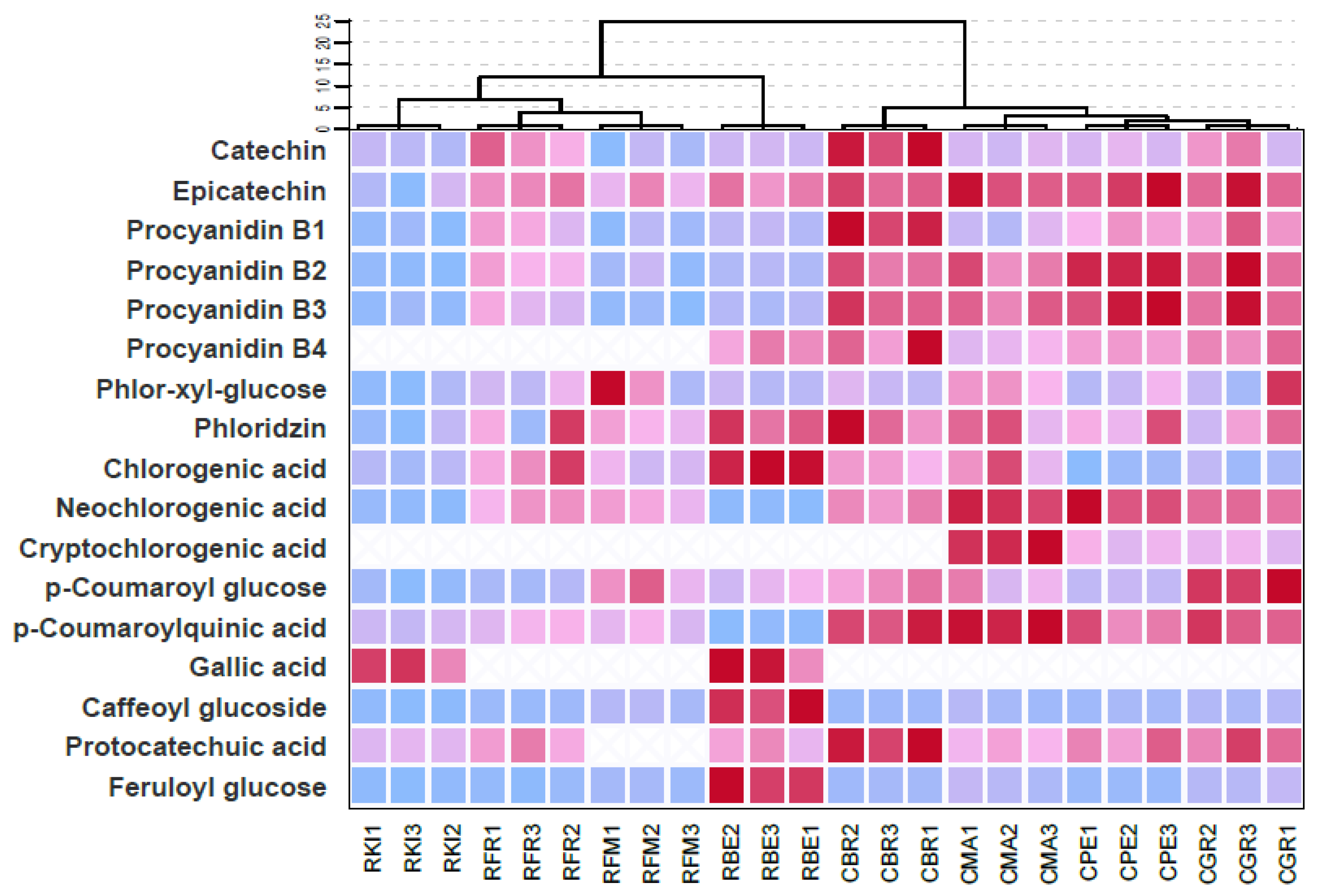
2.3. Discriminatory Analysis polyphenolic fingerprint
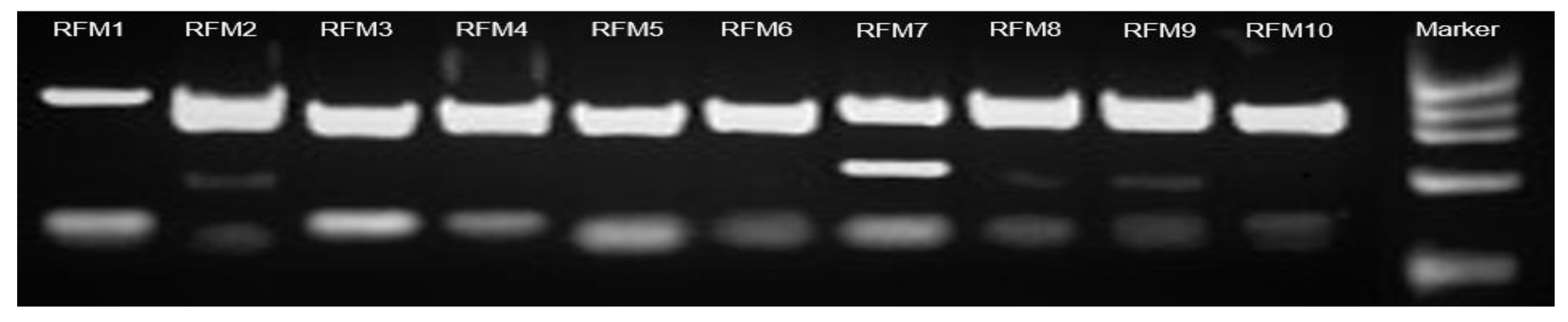
2.4. Polymorfism of ITS1 and ITS2
2.5. Phylogenetic Analysis Based on ITS1 and ITS2 Polymorfisms
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material, Pomological and Organoleptic Properties
3.3. Polyphenol Extraction
3.4. UPLC–MS Analysis
3.5. Total Antiradical Activity
3.6. DNA Extraction
3.7. PCR Amplification
3.8. Phylogenetic Analyses
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bartolini, S.; Viti, R.; Ducci, E. Local fruit varieties for sustainable cultivations: Pomological, nutraceutical and sensory characterization. Agrochimica 2015, 59, 281–284. [Google Scholar] [CrossRef]
- Wojdylo, A.; Oszmianski, J.; Laskowski, P. Polyphenolic compounds and antioxidant activity of new and old apple varieties. J. Agric. Food Chem. 2008, 56, 6520–6530. [Google Scholar] [CrossRef] [PubMed]
- Masi, E.; Taiti, C.; Vignolini, P.; Petrucci, A.W.; Giordani, E.; Heimler, D.; Romani, A.; Mancuso, S. Polyphenols and aromatic volatile compounds in biodynamic and conventional “Golden Delicious” apples (Malus domestica Bork.). Eur. Food Res. Technol. 2017, 243, 1519–1531. [Google Scholar] [CrossRef]
- Escarpa, A.; González, M.C. High-performance liquid chromatography with diode-array detection for the determination of phenolic compounds in peel and pulp from different apple varieties. J. Chromatogr. A 1998, 823, 331–337. [Google Scholar] [CrossRef]
- Iacopini, P.; Camangi, F.; Stefani, A.; Sebastiani, L. Antiradical potential of ancient Italian apple varieties of Malus×domestica Borkh. in a peroxynitrite-induced oxidative process. J. Food Compos. Anal. 2010, 23, 518–524. [Google Scholar] [CrossRef]
- Jakobek, L.; Barron, A.R. Ancient apple varieties from Croatia as a source of bioactive polyphenolic compounds. J. Food Compos. Anal. 2016, 45, 9–15. [Google Scholar] [CrossRef]
- Hyson, D.A. A Comprehensive Review of Apples and Apple Components and Their Relationship to Human Health. Adv. Nutr. 2012, 2, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, N.P.; Bondonno, C.P.; Ward, N.C.; Hogdson, J.M.; Croft, K.D. The cardiovascular health benefits of apples: Whole fruit vs. isolated compounds. Trends Food Sci. Technol. 2017, 69, 243–256. [Google Scholar] [CrossRef]
- Alcázar, E.J. Protecting crop genetic diversity for food security: Political, ethical and technical challenges. Nat. Rev. Genet. 2005, 6, 946–953. [Google Scholar] [CrossRef]
- Horrigan, L.; Lawrence, R.S.; Walker, P. How sustainable agriculture can address the environmental and human health harms of industrial agriculture. Environ. Health Perspect. 2002, 110, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Lindenmayer, D.B.; Manning, A.D. Biodiversity, ecosystem function, and resilience: Ten guiding principles for commodity production landscapes. Front. Ecol. Environ. 2006, 4, 80–86. [Google Scholar] [CrossRef]
- Di Falco, S.; Chavas, J.P. Rainfall shocks, resilience, and the effects of crop biodiversity on agroecosystem productivity. Land Econ. 2008, 84, 83–96. [Google Scholar] [CrossRef]
- Tartarini, S.; Gennari, F.; Pratesi, D.; Palazzetti, C.; Sansavini, S.; Parisi, L.; Fouillet, A.; Durel, C.E. Characterisation and genetic mapping of a major scab resistance gene from the old Italian apple cultivar “Durello di Forlì”. Acta Hortic. 2004, 663, 129–134. [Google Scholar] [CrossRef]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Torello Marinoni, D.; Cerutti, A.K.; Canterino, S.; Bounous, G. Application of sensory, nutraceutical and genetic techniques to create a quality profile of ancient apple cultivars. J. Food Qual. 2012, 35, 169–181. [Google Scholar] [CrossRef]
- Maragò, E.; Michelozzi, M.; Calamai, L.; Camangi, F.; Sebastiani, L. Antioxidant properties, sensory characteristics and volatile compounds profile of apple juices from ancient Tuscany (Italy) apple varieties. Eur. J. Hortic. Sci. 2016, 81, 255–263. [Google Scholar] [CrossRef]
- Tuna, M.; Khadka, D.K.; Shrestha, M.K.; Arumuganathan, K.; Golan-Goldhirsh, A. Characterization of natural orchardgrass (Dactylis glomerata L.) populations of Thrace region of Turkey base on ploidy and DNA polymorphisms. Euphytica 2004, 135, 39–46. [Google Scholar] [CrossRef]
- Meudt, H.M.; Clarke, A.C. Almost forgotten or latest practice? AFLP applications, analyses and advances. Trends Plant Sci. 2007, 12, 106–117. [Google Scholar] [CrossRef]
- Mondini, L.; Noorani, A.; Pagnotta, M.A. Assessing plant genetic diversity by molecular tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef]
- Poczai, P.; Hyvönen, J. Nuclear ribosomal spacer regions in plant phylogenetics: Problems and prospects. Mol. Biol. Rep. 2010, 37, 1897–1912. [Google Scholar] [CrossRef]
- Thormann, C.E.; Fereira, M.E.; Camargo, L.E.A.; Tivang, J.G.; Osborn, T.C. Comparison of RFLP and RAPD markers to estimating genetic relationships within and among cruciferous species. Theor. Appl. Genet. 1994, 88, 973–980. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Ahmad Haji, R.F.; Tiwari, S.; Gandhi, S.G.; Kumar, A.; Brindavanam, N.B.; Verma, V. Genetic diversity analysis among accessions of Desmodium gangeticum (L) DL with Simple Sequence Repeat (SSR) and Internal Transcribed Spacer (ITS) Regions for species conservation. J. Biodivers., Bioprospect. Dev. 2016, 3, 2–5. [Google Scholar]
- Le Cam, B.; Devaux, M.; Parisi, L. Specific polymerase chain reaction identification of Venturia nashicola using internally transcribed spacer region in the ribosomal DNA. Phytopathology 2001, 91, 900–904. [Google Scholar] [CrossRef]
- Bernardi, R.; Manzo, M.; Durante, M.; Petrucelli, R.; Bartolini, G. Molecular markers for cultivar characterisation in olea Olea europaea L. Acta Hortic. 2002, 586, 97–100. [Google Scholar] [CrossRef]
- Drogoudi, P.D.; Michailidis, Z.; Pantelidis, G. Peel and flesh antioxidant content and harvest quality characteristics of seven apple cultivars. Sci. Hortic. 2008, 115, 149–153. [Google Scholar] [CrossRef]
- Aguilar-Rosas, S.F.; Ballinas-Casarrubias, M.L.; Nevarez-Moorillon, G.V.; Martin-Belloso, O.; Ortega-Rivas, E. Thermal and pulsed electric fields pasteurization of apple juice: Effects on physicochemical properties and flavour compounds. J. Food Eng. 2007, 83, 41–46. [Google Scholar] [CrossRef]
- Vangdal, E. Quality criteria for fresh consumption. Acta Agric. Scand. 1985, 35, 41–47. [Google Scholar] [CrossRef]
- Skendrović, B.M.; Kresimir, I.; Druzic, J.; Kovac, A.; Voca, S. Chemical and sensory characteristics of three apple cultivars (Malus x domestica Borkh.). Agric. Conspec. Sci. 2007, 72, 317–322. [Google Scholar]
- Rodriguez-Casado, A. The health potential of fruits and vegetables phytochemicals: Notable examples. Crit. Rev. Food Sci. Nutr. 2016, 56, 1097–1107. [Google Scholar] [CrossRef]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, 2647–2659. [Google Scholar] [CrossRef]
- Verhoeyen, M.E.; Bovy, A.; Collins, G.; Muir, S.; Robinson, S.; de Vos, C.H.R.; Colliver, S. Increasing antioxidant levels in tomatoes through modification of the flavonoid biosynthetic pathway. J. Exp. Bot. 2002, 53, 2099–2106. [Google Scholar] [CrossRef]
- Toh, J.Y.; Tan, V.M.; Lim, P.C.; Lim, S.T.; Chong, M.F. Flavonoids from fruit and vegetables: A focus on cardiovascular risk factors. Curr. Atheroscler. Rep. 2013, 15, 368–375. [Google Scholar] [CrossRef]
- Arts, I.C.W.; Sesink, A.L.A.; Faassen-Peters, M.; Hollman, P.C.H. The type of sugar moiety is a major determinant of the small intestinal uptake and subsequent biliary excretion of dietary quercetin glycosides. Br. J. Nutr. 2004, 91, 841–847. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Dupont, N.; Guillermin, P. Concentrations and characteristics of procyanidins and other phenolics in apples during fruit growth. Phytochemistry 2007, 68, 1128–1138. [Google Scholar] [CrossRef]
- Zanotti, I.; Dall’Asta, M.; Mena, P.; Mele, L.; Bruni, R.; Ray, S.; Del Rio, D. Atheroprotective effects of (poly)phenols: A focus on cell cholesterol metabolism. Food Funct. 2015, 6, 13–31. [Google Scholar] [CrossRef]
- Marks, S.C.; Mullen, W.; Borges, G.; Crozier, A. Absorption, metabolism, and excretion of cider dihydrochalcones in healthy humans and subjects with an ileostomy. J. Agric. Food Chem. 2009, 57, 2009–2015. [Google Scholar] [CrossRef]
- Walle, T.; Walle, U.K. The ß-D-glucoside and sodium-dependent glucose transporter 1 (SGLT1)-inhibitor phloridzin is transported by both SGLT1 and multidrug resistance-associated proteins 1/2. Drug Metab. Dispos. 2003, 31, 1288–1291. [Google Scholar] [CrossRef]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef]
- Chinnici, F.; Bendini, A.; Gaiani, A.; Riponi, C. Radical scavenging activities of peels and pulps from cv. Golden Delicious apples as related to their phenolic composition. J. Agric. Food Chem. 2004, 52, 4684–4689. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef]
- Reber, J.D.; Eggett, D.L.; Parker, T.L. Antioxidant capacity interactions and a chemical/structural model of phenolic compounds found in strawberries. Int. J. Food Sci. Nutr. 2011, 65, 445–452. [Google Scholar] [CrossRef]
- Bolling, B.W.; Chen, Y.Y.; Chen, C.Y.O. Contributions of phenolics and added vitamin C to the antioxidant capacity of pomegranate and grape juices: Synergism and antagonism among constituents. Int. J. Food Sci. Technol. 2013, 48, 2650–2658. [Google Scholar] [CrossRef]
- Treutter, D. Biosynthesis of phenolic compounds and its regulation in apple. Plant Growth Regul. 2001, 34, 71–89. [Google Scholar] [CrossRef]
- Awad, M.A.; Wagenmakers, P.S.; De Jager, A. Effects of light on flavonoid and chlorogenic acid levels in the skin of “Jonagold” apples. Sci. Hortic. 2001, 88, 289–298. [Google Scholar] [CrossRef]
- Volz, R.K.; McGhie, T.K. Genetic variability in apple fruit polyphenol composition in Malus × domestica and Malus sieversii germplasm grown in New Zealand. J. Agric. Food Chem. 2011, 59, 11509–11521. [Google Scholar] [CrossRef] [PubMed]
- Łata, B.; Przeradzka, M.; Bínkowska, M. Great differences in antioxidant properties exist between 56 apple cultivars and vegetation seasons. J. Agric. Food Chem. 2005, 53, 8970–8978. [Google Scholar] [CrossRef]
- Łata, B.; Trampczynska, A.; Paczesna, J. Cultivar variation in apple peel and whole fruit phenolic composition. Sci. Hortic. 2009, 121, 176–181. [Google Scholar] [CrossRef]
- Landi, M.; Massai, R.; Remorini. Effect of rootstock and manual floral bud thinning on organoleptical and nutraceutical properties of sweet cherry (Prunus avium L.) cv ‘Lapins’. Agrochimica 2014, 58, 335–351. [Google Scholar] [CrossRef]
- Assumpção, C.F.; Hermes, V.S.; Pagno, C.; Castagna, A.; Mannucci, A.; Sgherri, C.; Pinzino, C.; Ranieri, A.; Flôres, S.H.; Rios, A.O. Phenolic enrichment in apple skin following post-harvest fruit UV-B treatment. Postharvest Biol. Tech. 2018, 138, 37–45. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. Lebenson Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Gawel, N.J.; Jarret, R.L. A modified CTAB DNA extraction procedure for Musa and Ipomoea. Plant Mol. Bio. Rep. 1991, 9, 262–266. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Apple Code | |||||||
|---|---|---|---|---|---|---|---|---|
| RKI | RBE | RFM | RFR | CMA | CBR | CGR | CPE | |
| Weight (g) | 132.81 ± 15.79 a | 46.00 ± 3.42 f | 110.64 ± 6.54 c | 124.43 ± 6.04 b | 69.05 ± 10.58 e | 94.73 ± 5.16 d | 69.33 ± 5.24 e | 89.36 ± 5.26 d |
| Width min (mm) | 70.02 ± 3.48 a | 47.48 ± 3.14 d | 66.07 ± 2.14 ab | 66.67 ± 2.10 ab | 53.80 ± 2.47 c | 60.86 ± 2.95 b | 57.66 ± 3.16 bc | 60.01 ± 4.12 b |
| Width max (mm) | 74.93 ± 5.11 a | 55.08 ± 3.33 c | 71.26 ± 8.50 a | 71.11 ± 3.31 a | 57.50 ± 1.34 c | 64.46 ± 2.85 b | 55.31 ± 2.29 c | 68.22 ± 2.82 ab |
| SSC (°Brix) | 15.60 ± 0.68 | 17.20 ± 1.40 | 15.40 ± 0.58 | 17.70 ± 0.98 | 16.80 ± 1.56 | 16.60 ± 1.10 | 14.80 ± 0.67 | 17.70 ± 1.90 |
| Titratable acidity (mg malic acid g−1 FW) | 4.28 ± 0.27 c | 2.71 ± 0.32 d | 3.83 ±0.82 c | 4.50 ± 0.23 c | 6.86 ± 0.29 a | 5.79 ± 0.59 b | 3.85 ± 0.39 c | 3.92 ± 0.39 c |
| Polyphenols | Apple Code | |||||||
|---|---|---|---|---|---|---|---|---|
| RKI | RBE | RFM | RFR | CMA | CBR | CGR | CPE | |
| Flavonols | ||||||||
| Q-galactoside | 0.03 ± 0.005 c | 0.07 ± 0.01 b | 0.04 ± 0.005 c | 0.05 ± 0.01 c | 0.05 ± 0.005 c | 0.08 ± 0.004 b | 0.14 ± 0.02 a | 0.04 ± 0.01 c |
| Q-glucoside | 0.17 ± 0.01 b | 0.35 ± 0.08 ab | 0.27 ± 0.04 b | 0.44 ± 0.04 a | 0.29 ± 0.01 b | 0.36 ± 0.15 ab | 0.43 ± 0.07 a | 0.26 ± 0.05 b |
| Q-arabinopyranoside | 0.07 ±0.006 c | 0.14 ± 0.03 b | 0.14 ± 0.02 b | 0.20 ± 0.04 a | 0.11 ± 0.002 bc | 0.12 ± 0.03 bc | 0.19 ± 0.01 ab | 0.14 ± 0.06 b |
| Q-arabinofuranoside | 0.08 ± 0.008 c | 0.35 ± 0.01 a | 0.20 ± 0.02 b | 0.29 ± 0.03 ab | 0.16 ± 0.01 bc | 0.24 ± 0.12 b | 0.31 ± 0.02 ab | 0.17 ± 0.07 b |
| Q-rhamnoside | 0.49 ± 0.054 d | 1.94 ± 0.12 a | 0.87 ± 0.07 c | 1.40 ± 0.35 b | 0.85 ± 0.04 c | 0.88 ± 0.20 c | 1.10 ± 0.18 c | 1.26 ± 0.11 bc |
| Total | 0.84 ± 0.06 e | 2.85 ± 0.22 a | 1.52 ± 0.15 cd | 2.38 ± 0.41 b | 1.46 ± 0.05 d | 1.68 ± 0.49 cd | 2.17 ± 0.27 bc | 1.87 ± 0.30 c |
| Flavanols | ||||||||
| Catechin | 37.07 ± 2.62 cd | 42.42 ± 0.89 c | 31.92 ± 7.57 d | 61.53 ± 6.93 b | 44.63 ± 2.27 c | 79.36 ± 6.42 a | 55.82 ± 11.06 bc | 46.09 ± 2.43 c |
| Epicatechin | 191.08 ± 39.88 c | 307.65 ± 13.09 b | 272.16 ± 29.17 b | 305.63 ± 10.26 b | 354.08 ± 28.82 ab | 336.45 ± 14.31 ab | 345.65 ± 35.30 ab | 361.40 ± 29.53 a |
| Procyanidin B1 | 5.47 ± 0.40 e | 6.99 ± 0.34 de | 6.01 ± 0.92 e | 9.39 ± 0.95 c | 7.55 ± 0.87 d | 13.11 ± 0.79 a | 10.72 ± 0.96 b | 9.91 ± 0.46 bc |
| Procyanidin B2 | 4.62 ± 0.17 e | 5.97 ± 0.21 d | 5.68 ± 1.12 de | 8.94 ± 0.40 c | 10.51 ± 0.92 b | 10.75 ± 0.64 b | 11.43 ± 1.50 b | 12.52 ± 0.18 a |
| Procyanidin B3 | 43.53 ± 3.53 e | 55.06 ± 2.41 d | 42.35 ± 3.70 e | 78.08 ± 9.45 c | 105.19 ± 6.36 b | 111.87 ± 7.04 b | 112.99 ± 14.93 ab | 123.93 ± 10.51 a |
| Procyanidin B4 | - | 1.03 ± 0.11 b | - | - | 0.72 ± 0.08 c | 1.30 ± 0.37 a | 1.11 ± 0.10 ab | 0.97 ± 0.01 b |
| Total | 281.77 ± 35.03 d | 419.12 ± 13.96 b | 358.12 ± 41.02 c | 463.57 ± 9.26 b | 522.68 ± 31.97 a | 552,84 ± 26.42 a | 537.72 ± 60.36 a | 554.82 ± 39.53 a |
| Dihydrochalcones | ||||||||
| Phlor-xyl-glucose | 29.78 ± 7.35 | 40.34 ± 3.05 | 63.05 ± 26.89 | 46.69 ± 6.86 | 61.50 ± 3.56 | 44.64 ± 4.80 | 52.30 ± 25.58 | 45.59 ± 9.06 |
| Phloridzin | 7.35 ± 2.67 c | 21.35 ± 1.96 a | 15.58 ± 1.42 ab | 15.39 ± 8.11 b | 18.64 ± 4.25 ab | 21.31 ± 4.37 a | 16.17 ± 4.47 ab | 17.49 ± 3.94 ab |
| Total | 37.13 ± 8.44 | 61.69 ± 4.98 | 78.63 ± 28.31 | 62.08 ± 14.91 | 80.14 ± 7.81 | 65.95 ± 9.17 | 68.47 ± 28.72 | 63.08 ± 12.67 |
| Phenolic acids | ||||||||
| Chlorogenic acid | 195.97 ± 13.84 d | 403.31 ± 10.80 a | 246.08 ± 20.94 c | 324.66 ± 42.79 b | 309.55 ± 51.13 b | 293.39 ± 12.96 b | 191.61 ± 20.97 d | 166.47 ± 13.33 d |
| Neochlorogenic acid | 7.20 ± 0.56 e | 6.78 ± 0.17 e | 17.96 ± 1.62 d | 19.05 ± 1.49 cd | 26.09 ± 1.21 a | 20.42 ± 0.93 c | 22.27 ± 0.38 b | 25.63 ± 2.77 a |
| Cryptochlorogenic acid | - | - | - | - | 0.71 ± 0.05 a | - | 0.32 ± 0.03 b | 0.34 ± 0.05 b |
| p -Coumaroyl glucose | 1.05 ± 0.14 e | 2.00 ± 0.24 c | 2.51 ± 0.44 b | 1.31 ± 0.10 de | 2.23 ± 0.44 bc | 2.58 ± 0.19 b | 3.35 ± 0.23 a | 1.60 ± 0.05 d |
| p -Coumaroylquinic acid | 64.96 ± 5.10 e | 27.52 ± 2.12 f | 80.82 ± 8.98 d | 86.21 ± 9.01 d | 154.50 ± 5.51 a | 138.28 ± 11.86 b | 131.00 ± 9.13 b | 118.79 ± 13.18 c |
| Gallic acid | 0.03 ± 0.005 | 0.03 ± 0.01 | - | - | - | - | - | - |
| Caffeoyl glucoside | 0.10 ± 0.01 c | 0.93 ± 0.10 a | 0.25 ± 0.04 b | 0.16 ± 0.01 c | 0.22 ± 0.05 bc | 0.17 ± 0.01 c | 0.25 ± 0.02 b | 0.20 ± 0.02 bc |
| Protocatechuic acid | 0.04 ± 0.002 d | 0.06 ± 0.01 c | - | 0.06 ± 0.01 c | 0.05 ± 0.005 c | 0.10 ± 0.01 a | 0.08 ± 0.01 b | 0.07 ± 0.01 bc |
| Feruloyl glucose | 8.64 ± 0.90 d | 75.47 ± 6.61 a | 15.56 ± 1.91 c | 10.47 ± 2.18 d | 22.91 ± 3.88 b | 15.60 ± 1.11 c | 23.68 ± 2.46 b | 12.63 ± 0.21 cd |
| Total | 277.97 ± 19.55 e | 516.10 ± 6.55 a | 363,18 ± 22.63 cd | 441.92 ± 53.28 b | 516.26 ± 48.26 a | 470.54 ± 3.72 b | 372.56 ± 28.43 c | 325,73 ± 6,64 d |
| Total polyphenols | 597.71 ± 61.94 d | 999.76 ± 11.81 b | 801.45 ± 59.47 c | 969.95 ± 60.13 b | 1120.54 ± 63.53 a | 1091.01 ± 37.78 a | 980.92 ± 23.74 b | 945.50 ± 53.04 b |
| Phenol | Correlation |
|---|---|
| Caffeoyl glucoside | 0.61 ** |
| Chlorogenic acid | 0.58 ** |
| Epicatechin | 0.51 * |
| Feruloyl glucose | 0.66 *** |
| Q-arabinofuranoside | 0.68 *** |
| Q-rhamnoside | 0.58 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Piccolo, E.; Viviani, A.; Guidi, L.; Remorini, D.; Massai, R.; Bernardi, R.; Landi, M. Discerning between Two Tuscany (Italy) Ancient Apple cultivars, ‘Rotella’ and ‘Casciana’, through Polyphenolic Fingerprint and Molecular Markers. Molecules 2019, 24, 1758. https://doi.org/10.3390/molecules24091758
Lo Piccolo E, Viviani A, Guidi L, Remorini D, Massai R, Bernardi R, Landi M. Discerning between Two Tuscany (Italy) Ancient Apple cultivars, ‘Rotella’ and ‘Casciana’, through Polyphenolic Fingerprint and Molecular Markers. Molecules. 2019; 24(9):1758. https://doi.org/10.3390/molecules24091758
Chicago/Turabian StyleLo Piccolo, Ermes, Ambra Viviani, Lucia Guidi, Damiano Remorini, Rossano Massai, Rodolfo Bernardi, and Marco Landi. 2019. "Discerning between Two Tuscany (Italy) Ancient Apple cultivars, ‘Rotella’ and ‘Casciana’, through Polyphenolic Fingerprint and Molecular Markers" Molecules 24, no. 9: 1758. https://doi.org/10.3390/molecules24091758
APA StyleLo Piccolo, E., Viviani, A., Guidi, L., Remorini, D., Massai, R., Bernardi, R., & Landi, M. (2019). Discerning between Two Tuscany (Italy) Ancient Apple cultivars, ‘Rotella’ and ‘Casciana’, through Polyphenolic Fingerprint and Molecular Markers. Molecules, 24(9), 1758. https://doi.org/10.3390/molecules24091758