
The Extracts of Artemisia absinthium L. Suppress the Growth of Hepatocellular Carcinoma Cells through Induction of Apoptosis via Endoplasmic Reticulum Stress and Mitochondrial-Dependent Pathway


Abstract
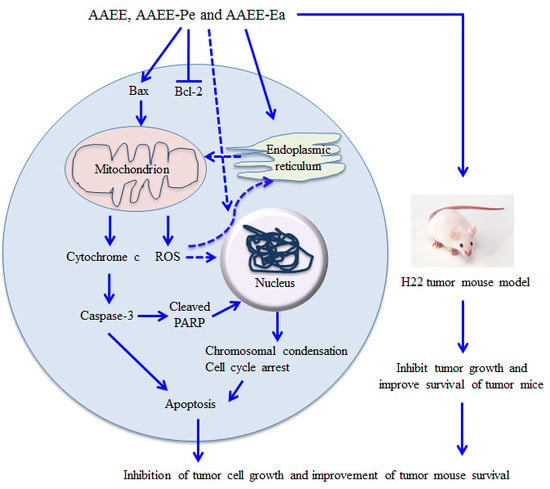
:
1. Introduction
2. Results
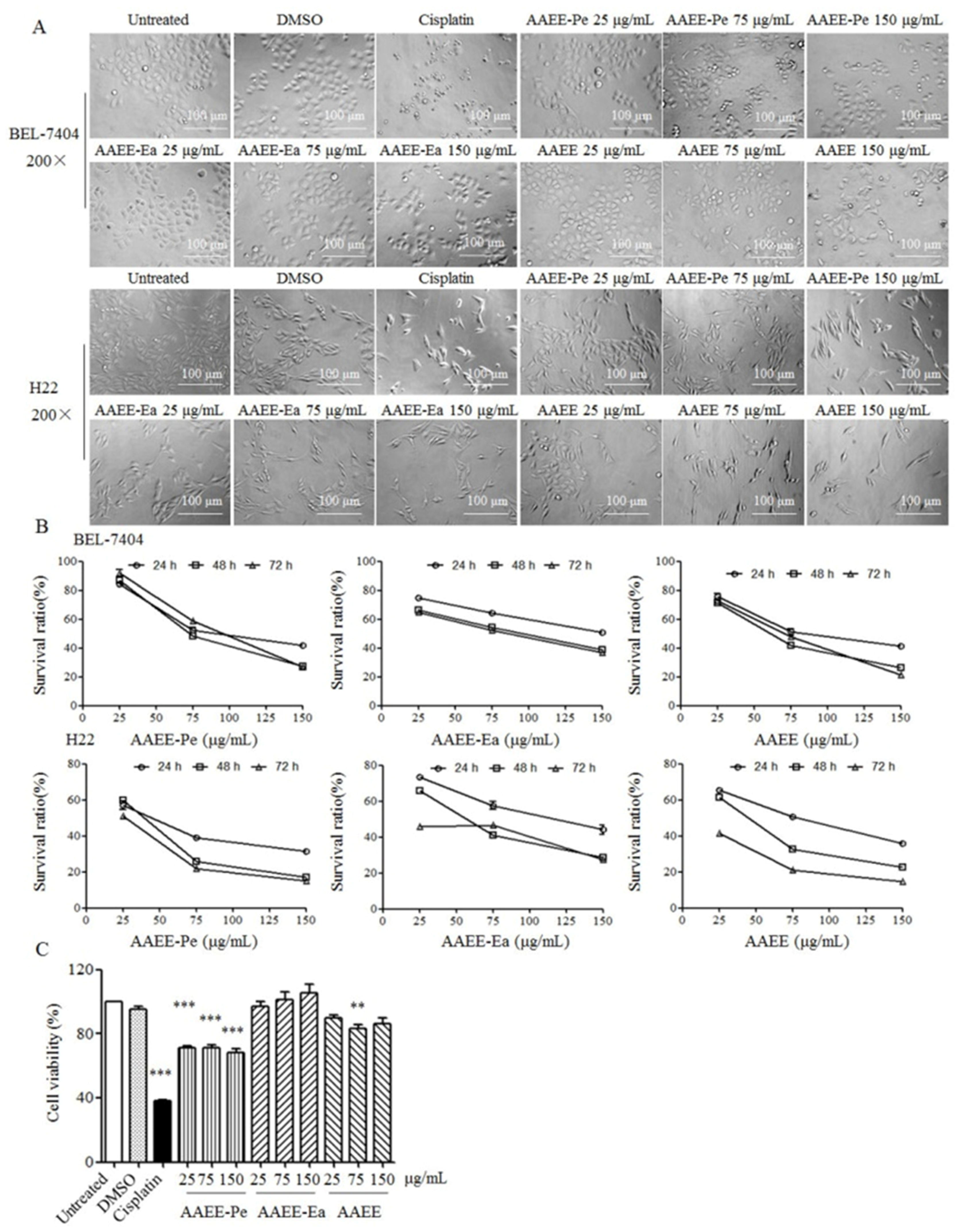
2.1. AAEE, AAEE-Pe, and AAEE-Ea Suppress the Growth of BEL-7404 and H22 Cells In Vitro
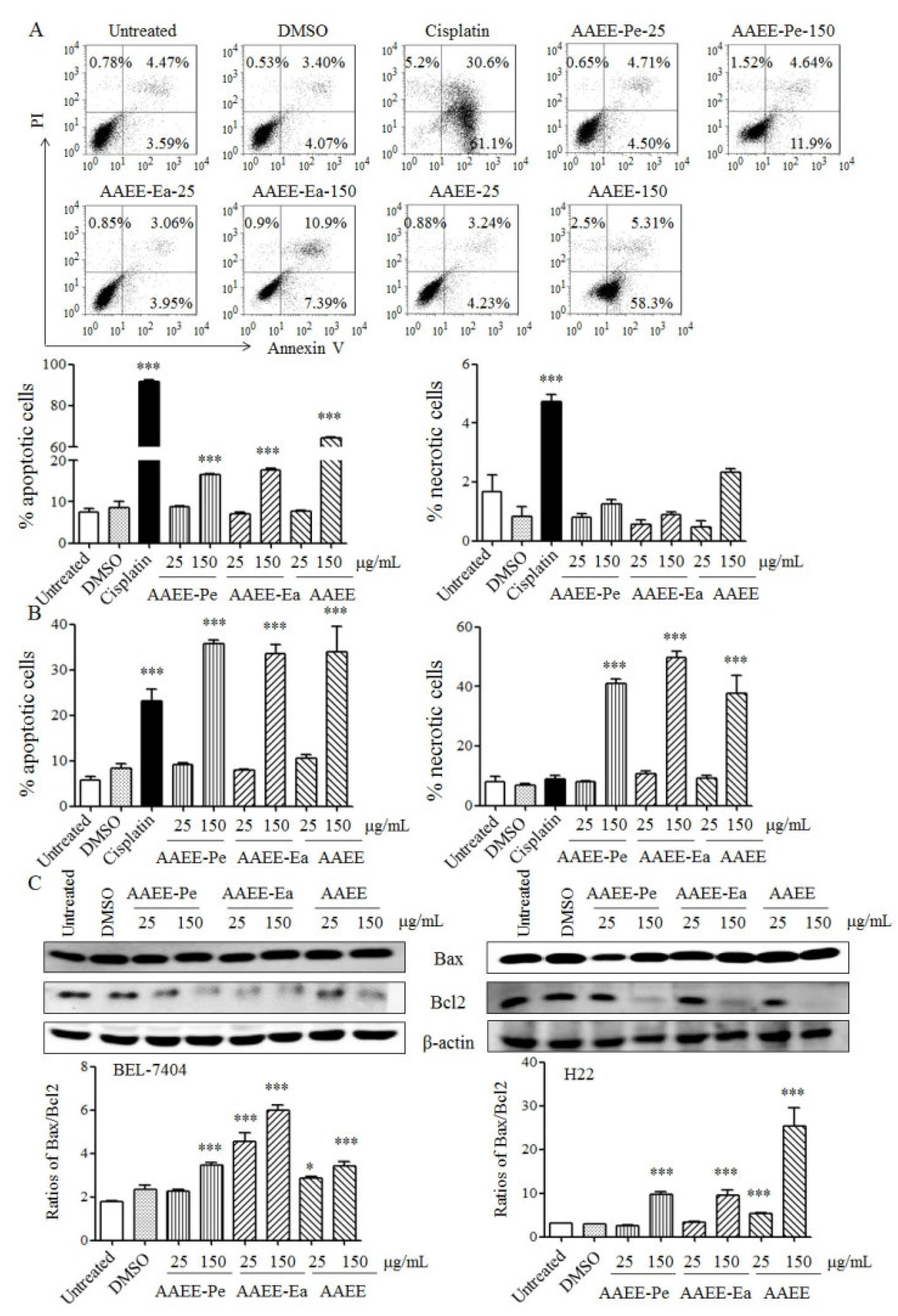
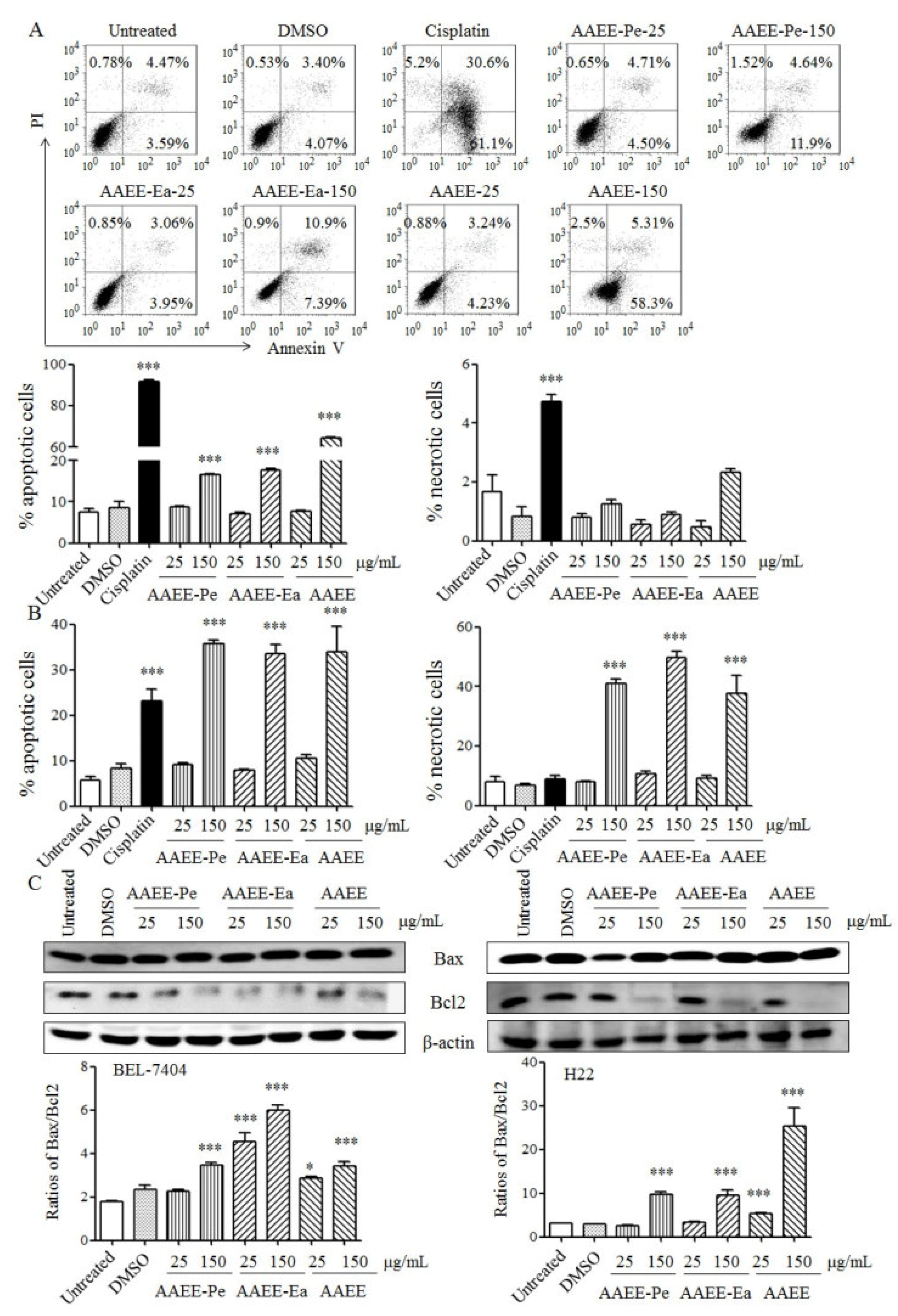
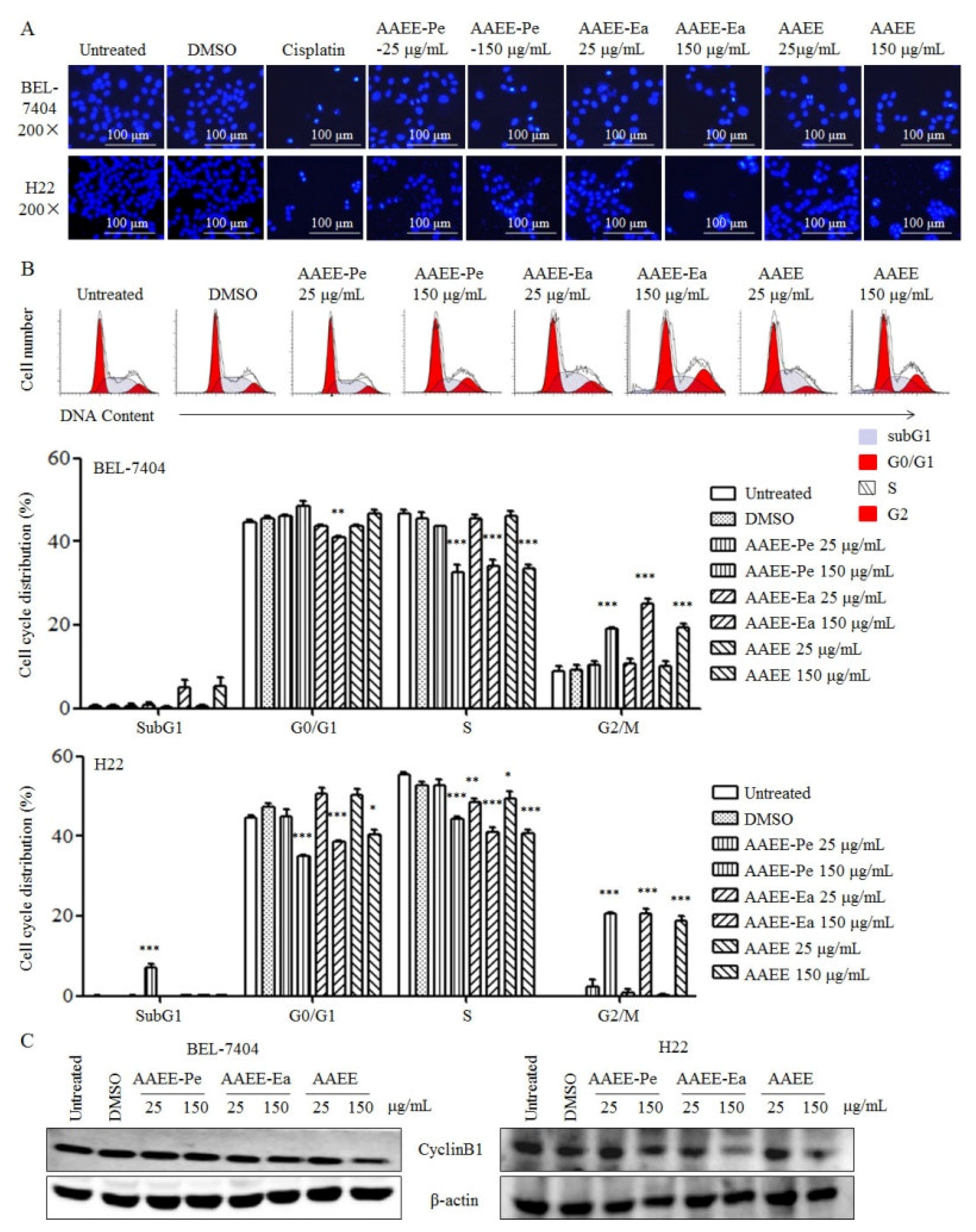
2.2. AAEE, AAEE-Pe, and AAEE-Ea Induce Apoptosis in BEL-7404 and H22 Cells
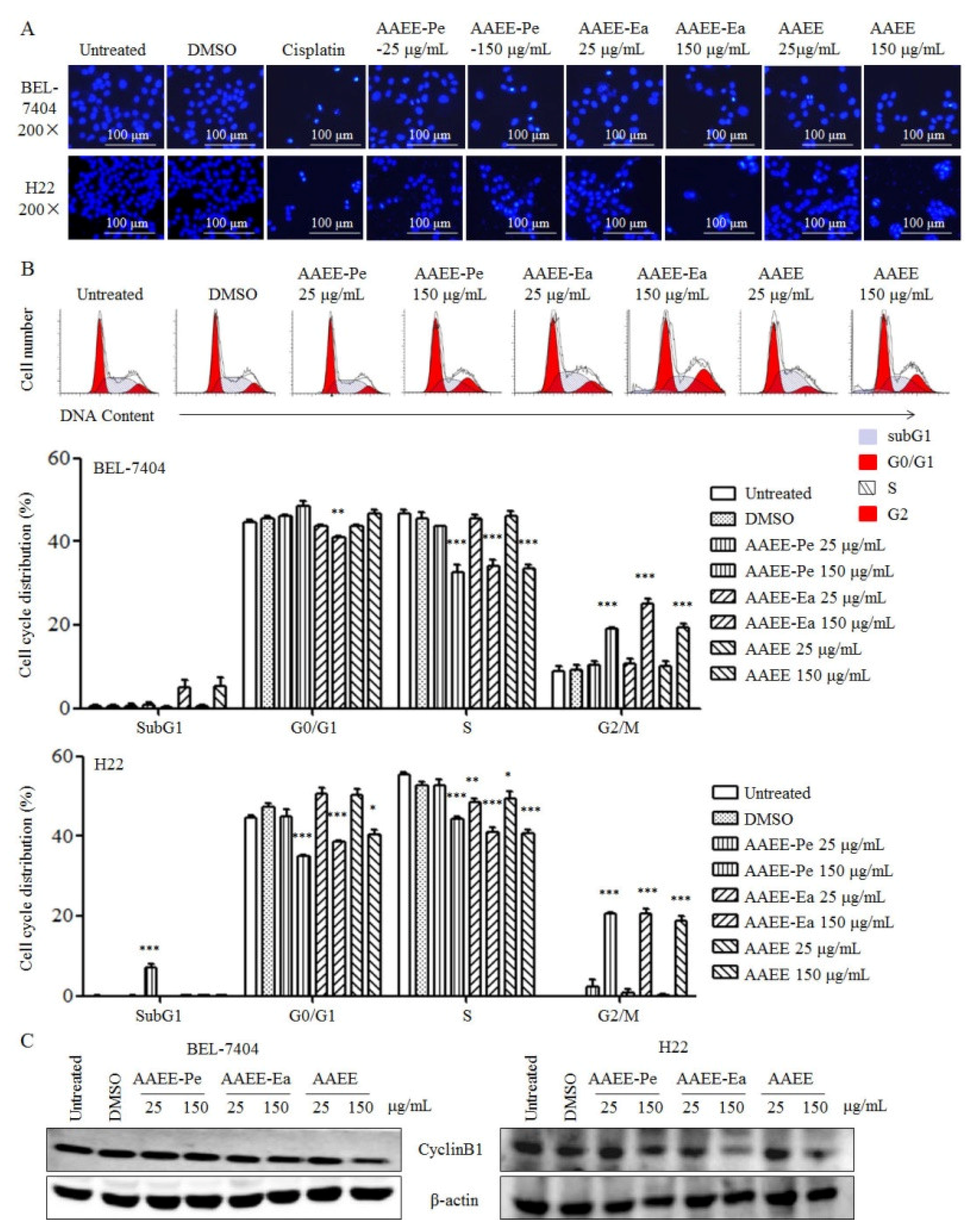
2.3. AAEE, AAEE-Pe, and AAEE-Ea Induce Cell Cycle Arrest in BEL-7404 and H22 Cells
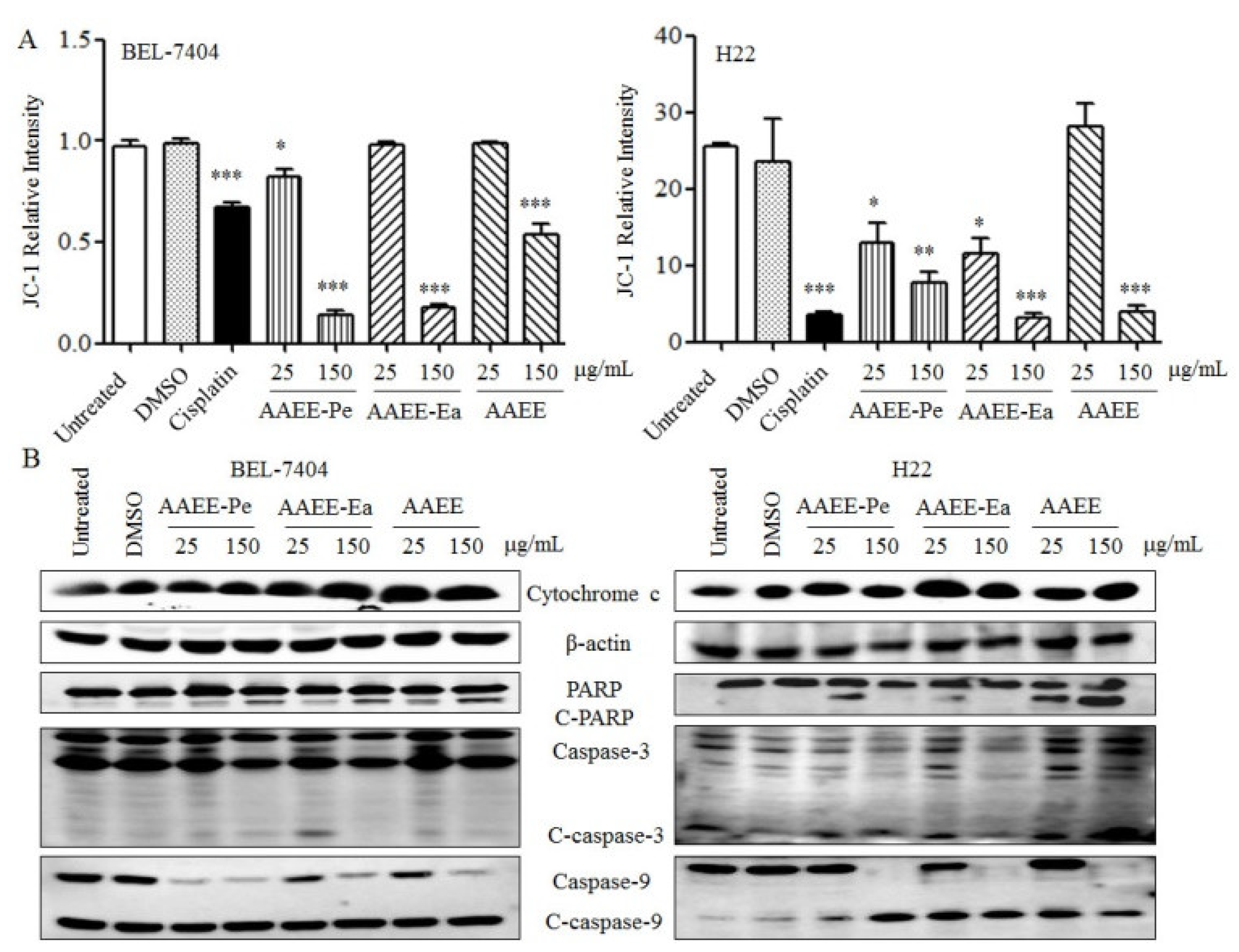
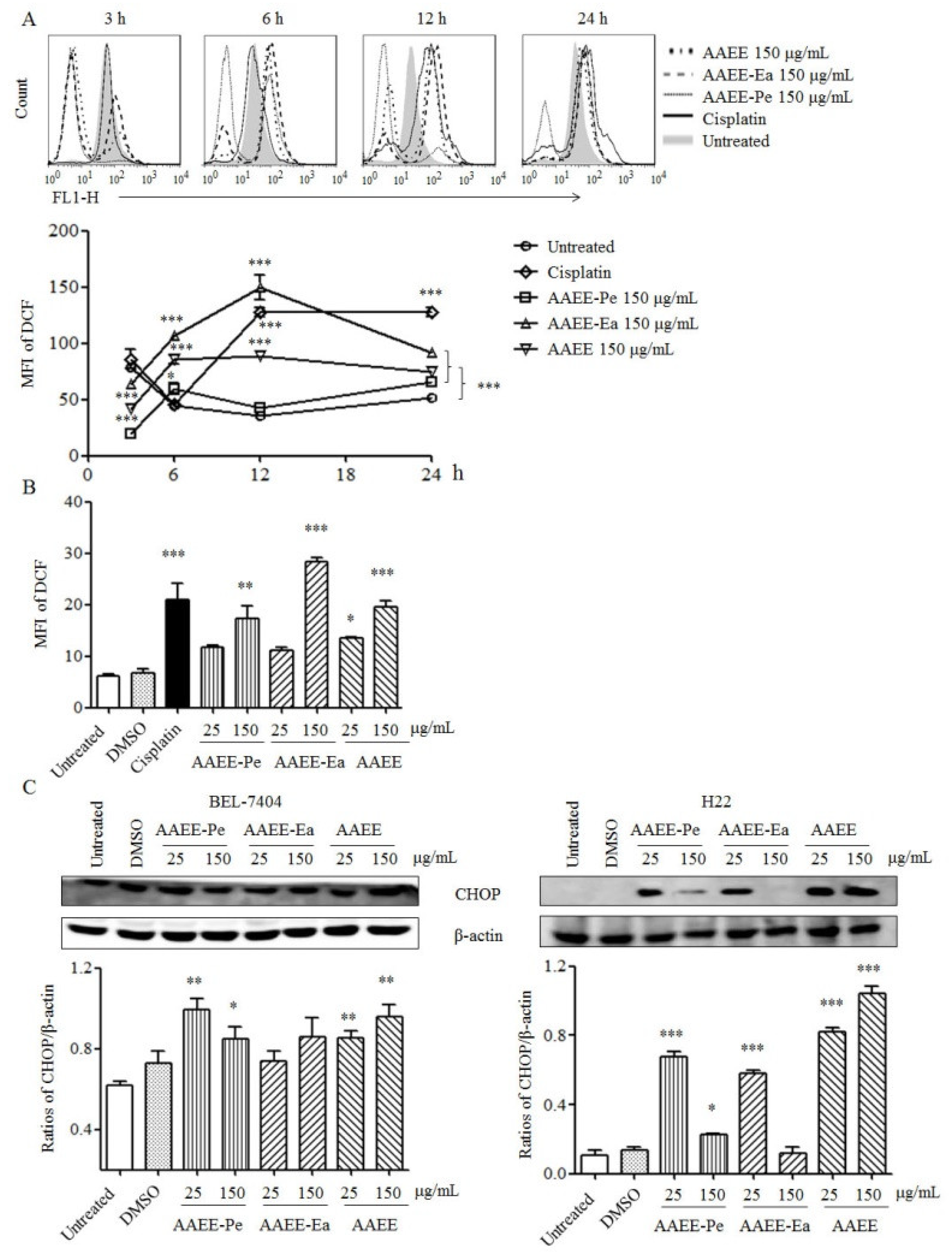
2.4. AAEE, AAEE-Pe, and AAEE-Ea Reduce Mitochondrial Membrane Potential (Δψm)
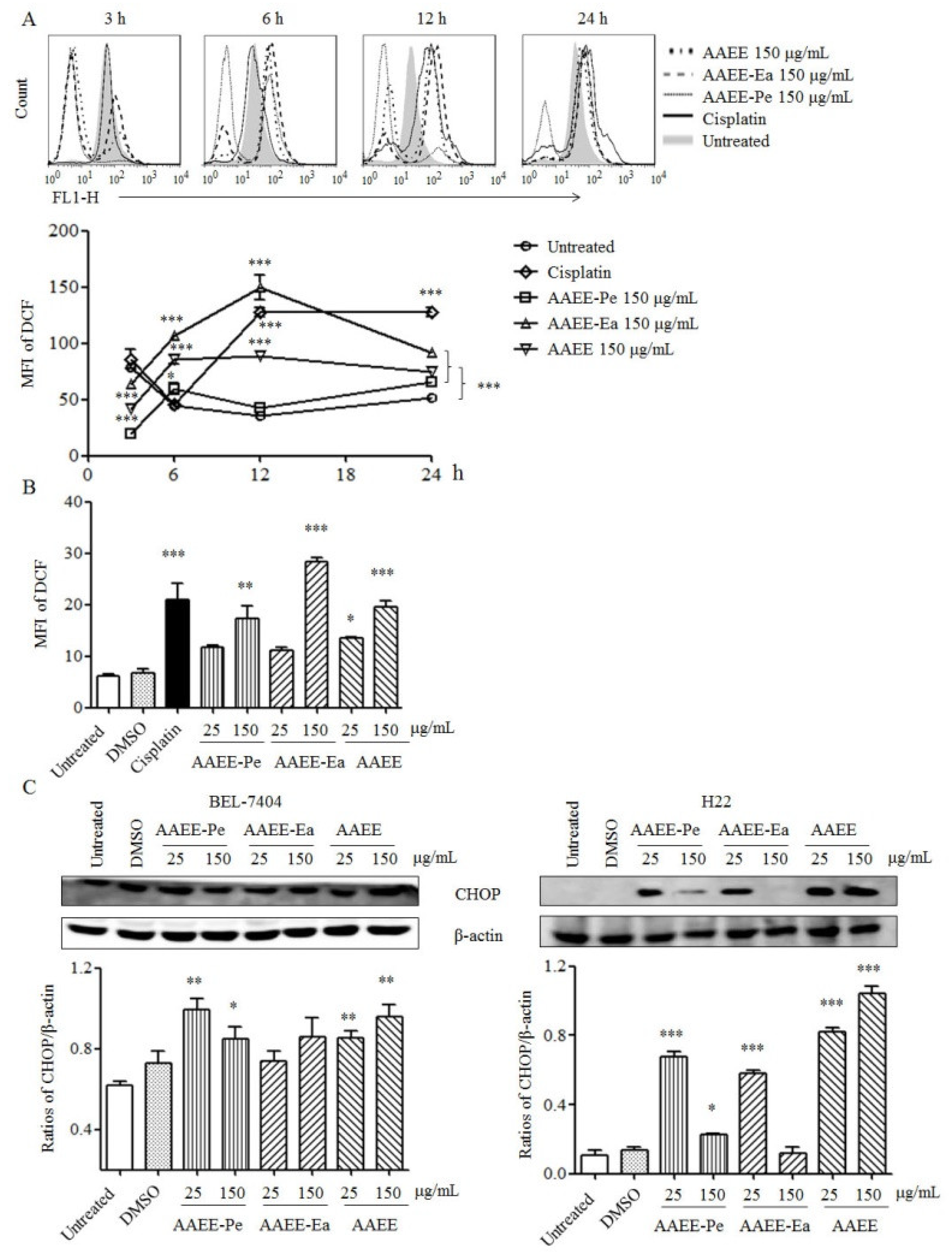
2.5. AAEE, AAEE-Pe, and AAEE-Ea Promote Reactive Oxygen Species (ROS) Generation and ER Stress
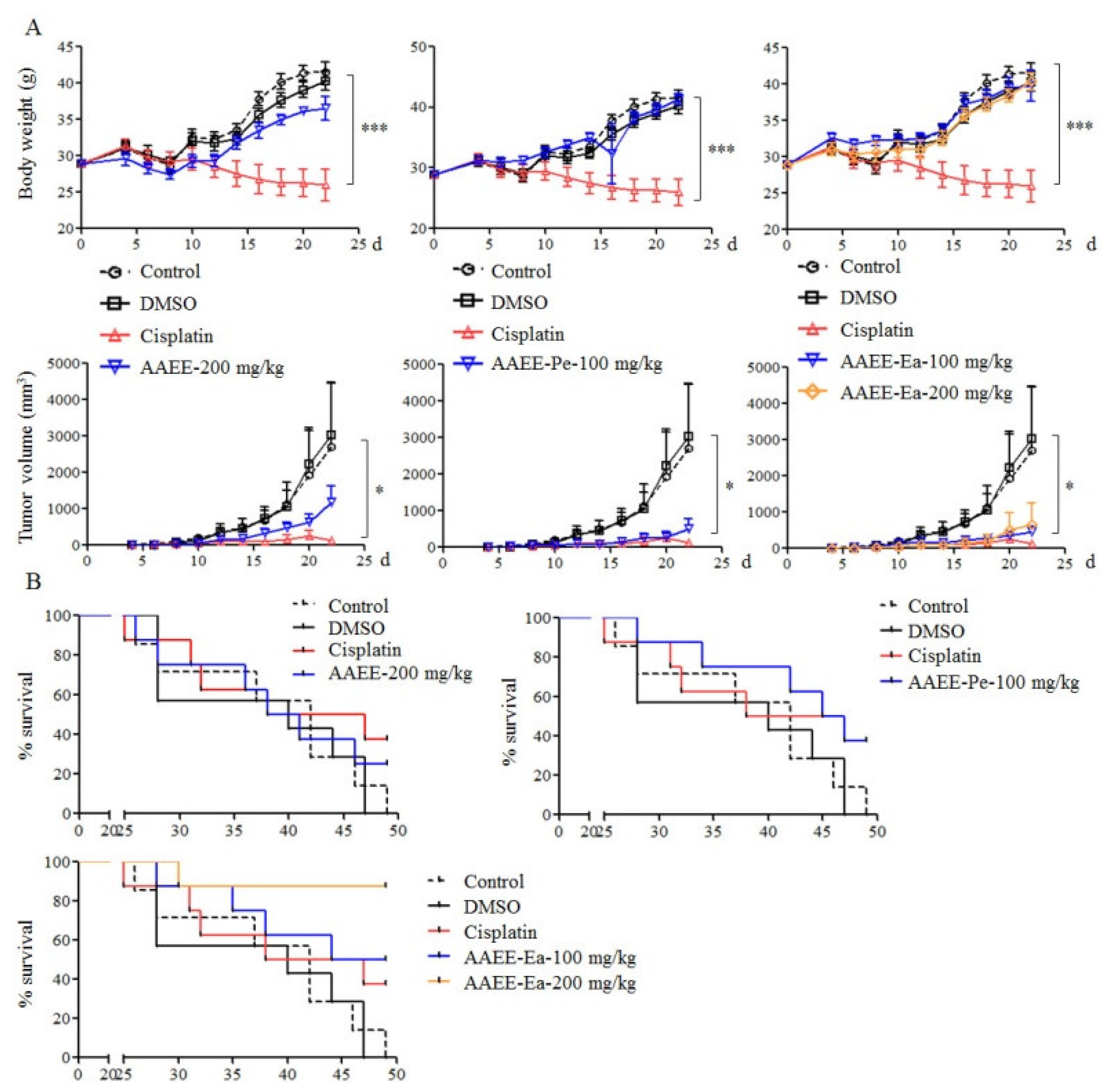
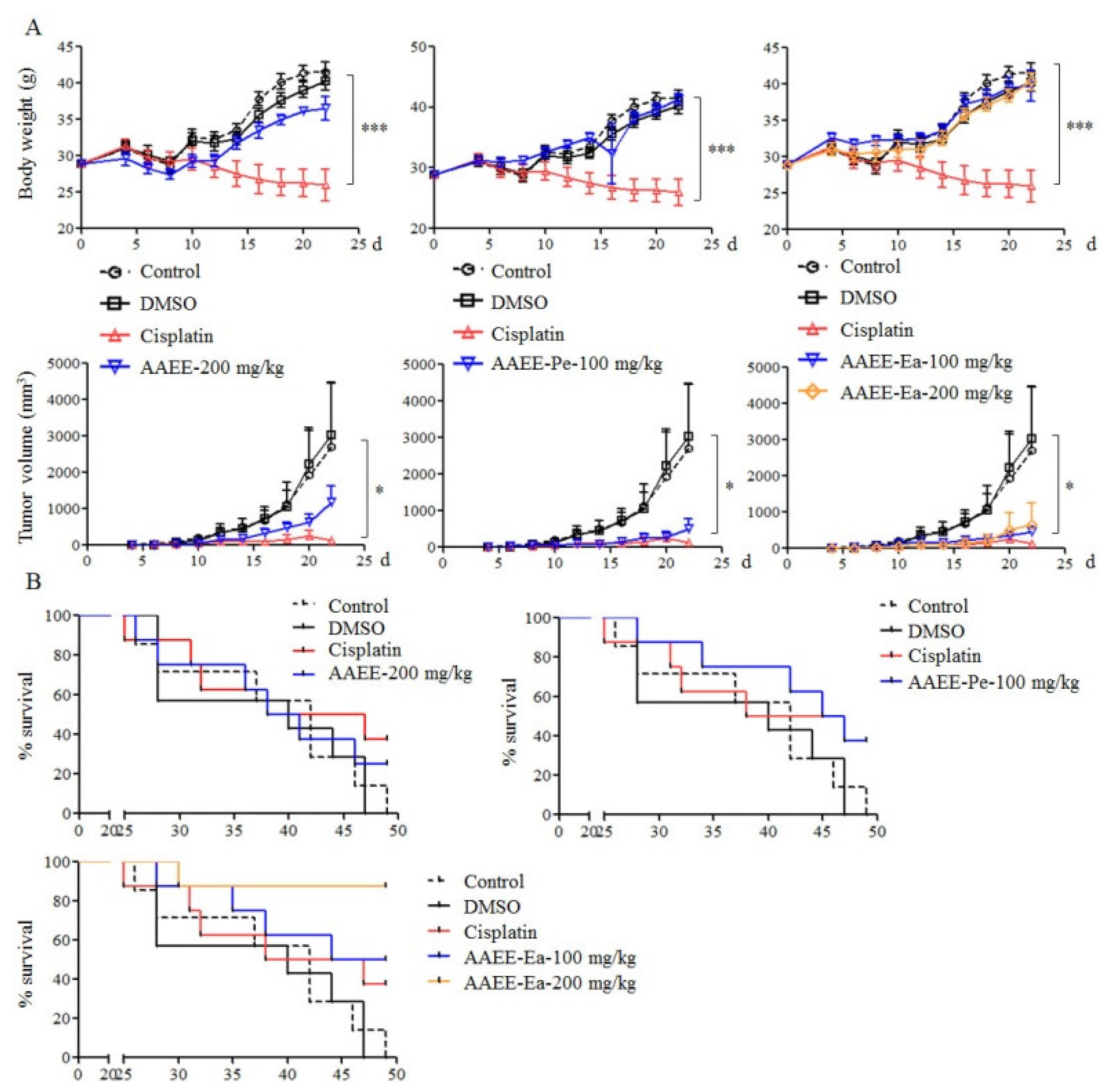
2.6. AAEE, AAEE-Pe, and AAEE-Ea Inhibit the Growth of H22 Cells In Vivo
3. Discussion
4. Materials and Methods
4.1. Preparation of AAEE, AAEE-Pe, and AAEE-Ea
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Observation of Cell Morphology
4.5. Analysis of Apoptosis
4.6. Hoechst 33258 Staining
4.7. Analysis of the Cell Cycle
4.8. Analysis of Δψm
4.9. Analysis of ROS
4.10. Western Blot
4.11. Animals and Ethics Statement
4.12. Tumor Mouse Study
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Wang, D.; Sun, Q.; Wu, J.; Wang, W.; Yao, G.; Li, T.; Li, X.; Li, L.; Zhang, Y.; Cui, W.; et al. A new Prenylated Flavonoid induces G0/G1 arrest and apoptosis through p38/JNK MAPKpathways in Human Hepatocellular Carcinoma cells. Sci. Rep. 2017, 7, 5736. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA-A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- London, W.T.; Petrick, J.L.; McGlynn, K.A.; Thun, M.J.; Linet, M.S.; Cerhan, J.R.; Haiman, C.A.; Schottenfeld, D. (Eds.) Cancer Epidemiology and Prevention, 4th ed.; Oxford University Press: New York, NY, USA, 2018; pp. 635–660. [Google Scholar]
- Mauer, K.; O’Kelley, R.; Podda, N.; Flanagan, S.; Gadani, S. New treatment modalities for hepatocellular cancer. Curr. Gastroenterol. Rep. 2015, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, T.; Zeng, L.; Chen, W. Efficacy and safety of radiofrequency ablation and transcatheter arterial chemoembolization for treatment of hepatocellular carcinoma: A meta-analysis. Hepatology 2016, 46, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Fan, X.; Tang, M.; Chen, R.; Wang, H.; Jia, R.; Zhou, X.; Jing, W.; Wang, H.J.; Yang, Y.; et al. Osteopontin induces autophagy to promote chemo-resistance in human hepatocellular carcinoma cells. Cancer Lett. 2016, 383, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Horgan, A.M.; Dawson, L.A.; Swaminath, A.; Knox, J.J. Sorafenib and radiation therapy for the treatment of advanced hepatocellular carcinoma. J. Gastrointest. Cancer 2012, 43, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Tabernero, J.; Garcia-Carbonero, R.; Cassidy, J.; Sobrero, A.; Van, C.E.; Kohne, C.H.; Tejpar, S.; Gladkov, O.; Davidenko, I.; Salazar, R.; et al. Sorafenib in combination with oxaliplatin, leucovorin, and fluorouracil (modified FOLFOX6) as first-line treatment of metastatic colorectal cancer: The respect trial. Clin. Cancer Res. 2013, 19, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Kamel, M.; Nidhal, S.; Olfa, B.; Slim, D.; Sonia, T.; Abdulkhaleg, A.; Khaldoun, A.S.; Wided, B.A.; Sana, A.; Adel, H.B.; et al. Chemical composition and antioxidant and antimicrobial activities of wormwood (Artemisia absinthium L.) essential oils and phenolics. J. Chem. 2015, 2015, 1–12. [Google Scholar]
- Azizi, K.; Shahidi-Hakak, F.; Asgari, Q.; Hatam, G.R.; Fakoorziba, M.R.; Miri, R.; Moemenbellah-Fard, M.D. In vitro efficacy of ethanolic extract of Artemisia absinthium (Asteraceae) against Leishmania major L. using cell sensitivity and flow cytometry assays. J. Parasit. Dis. 2016, 40, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, B.; Monzote, L.; Piñón, A.; Machín, L.; García, M.; Scull, R.; Setzer, W.N. In vitro and in vivo evaluation of essential oil from Artemisia absinthium L. formulated in nanocochleates against cutaneous Leishmaniasis. Medicines 2017, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Turak, A.; Shi, S.P.; Jiang, Y.; Tu, P.F. Dimeric guaianolides from Artemisia absinthium. Phytochemistry 2014, 105, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Caner, A.; Doskaya, M.; Degirmenci, A.; Can, H.; Baykan, S.; Uner, A.; Basdemir, G.; Zeybek, U.; Guruz, Y. Comparison of the effects of Artemisia vulgaris and Artemisia absinthium growing in western Anatolia against trichinellosis (Trichinella spiralis) in rats. Exp. Parasitol. 2008, 119, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Amat, N.; Upur, H.; Blazeković, B. In vivo hepatoprotective activity of the aqueous extract of Artemisia absinthium L. againstchemically and immunologically induced liver injuries in mice. J. Ethnopharmacol. 2010, 131, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Gilani, A.H.; Janbaz, K.H. Preventive and curative effects of Artemisia absinthium on acetaminophen and CCl4-induced hepatotoxicity. Gen. Pharmacol. 1995, 26, 309. [Google Scholar] [CrossRef]
- Craciunescu, O.; Constantin, D.; Gaspar, A.; Toma, L.; Utoiu, E.; Moldovan, L. Evaluation of antioxidant and cytoprotective activities of Arnica montana L. and Artemisia absinthium L. ethanolic extracts. Chem. Cent. J. 2012, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bora, K.S.; Sharma, A. Evaluation of antioxidant and free-radical scavenging potential of Artemisia absinthium. Pharm. Biol. 2011, 49, 1216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Z.; Du, H.X.; Huang, Z.X.; Zhang, P.; Yue, Y.Y.; Wang, W.Y.; Liu, W.; Zeng, J.; Ma, J.B.; Chen, G.Q.; et al. Thymoquinone induces apoptosis in bladder cancer cell via endoplasmic reticulum stress-dependent mitochondrial pathway. Chem.-Biol. Interact. 2018, 292, 65–75. [Google Scholar] [CrossRef]
- Vuyolwethu, S.; Georgia, S.; Roger, H.; Grafov, A.; Grafova, I.; Nieger, M.; Katz, A.A.; Parker, M.I.; Kaschula, C.H. The cytotoxicity of the ajoene analogue BisPMB in WHCO1 oesophageal cancer cells is mediated by CHOP/GADD153. Molecules 2017, 22, 892. [Google Scholar] [CrossRef]
- Zhang, H.P. Drug Standard of Ministry of Public Health of the People’s Republic of CHINA Xingjiang Technological and Health Publishing House, Xingjiang, Uighur Medicine Part; Xinjiang Publishing House of Science: Urumqi, China, 1999; p. 53. [Google Scholar]
- Song, G.C.; Yu, Y.J.; Wang, X.J. Experiments on antitumor activity of Scutellaria barbata polysaccharides and its immunological mechanisms. Open Access Libr. J. 2011, 13, 641–643. [Google Scholar]
- Wang, D.D.; Wu, Q.X.; Pan, W.J.; Hussain, S.; Mehmood, S.; Chen, Y. A novel polysaccharide from the Sarcodon aspratus triggers apoptosis in Hela cells via induction of mitochondrial dysfunction. Food Nutr. Res. 2018, 62, 1285. [Google Scholar] [CrossRef]
- Ashokkumar, R.; Jamuna, S.; Sakeena Sadullah, M.S.; Niranjali, D.S. Vitexin protects isoproterenol induced post myocardial injury by modulating hipposignaling and ER stress responses. Biochem. Biophys. Res. Commun. 2018, 496, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Yang, Y.; Liu, Y.; Jiang, S.; Di, S.Y.; Hu, W.; Ma, Z.Q.; Li, T.; Zhu, Y.F.; Xin, Z.L.; et al. Icariin displays anticancer activity against human esophageal cancer cells via regulating endoplasmic reticulum stress-mediated apoptotic signaling. Sci. Rep. 2016, 6, 21145. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.L.; Zheng, Y.F.; Gu, J.Y.; Wang, S.Q.; Wang, N.; Yang, B.W.; Zhang, F.X.; Wang, D.M.; Fu, W.J.; Wang, Z.Y. Betulinic acid chemosensitizes breast cancer by triggering ER stress-mediated apoptosis by directly targeting GRP78. Cell Death Dis. 2018, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhou, T.Y.; Nie, C.H.; Wang, D.L.; Zheng, S.S. Bigelovin, a sesquiterpene lactone, suppresses tumor growth through inducing apoptosis and autophagy via the inhibition of mTOR pathway regulated by ROS generation in liver cancer. Biochem. Biophys. Res. Commun. 2018, 499, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S. Effects of dietary flavonoids on apoptotic pathways related to cancer chemoprevention. J. Nutr. Biochem. 2007, 18, 427–442. [Google Scholar] [CrossRef]
- Marzo, I.; Naval, J. Bcl-2 family members as molecular targets in cancer therapy. Biochem. Pharmacol. 2008, 76, 939–946. [Google Scholar] [CrossRef]
- Garner, T.P.; Lopez, A.; Reyna, D.E.; Spitz, A.Z.; Gavathiotis, E. Progress in targeting the BCL-2 family of proteins. Curr. Opin. Chem. Biol. 2017, 39, 133–142. [Google Scholar] [CrossRef]
- Shafi, G.; Hasan, T.N.; Syed, N.A.; Al-Hazzani, A.A.; Alshatwi, A.A.; Jyothi, A.; Munshi, A. Artemisia absinthium(AA): A novel potential complementary and alternative medicine for breast cancer. Mol. Biol. Rep. 2012, 39, 7373–7379. [Google Scholar] [CrossRef]
- Banerjee, A.; Banerjee, V.; Czinn, S.; Blanchard, T. Increased reactive oxygen species levels cause ER stress and cytotoxicity in andrographolide treated colon cancer cells. Oncotarget 2017, 8, 26142–26153. [Google Scholar] [CrossRef]
- Yu, X.S.; Du, J.; Fan, Y.J.; Liu, F.J.; Cao, L.L.; Liang, N.; Xu, D.G.; Zhang, J.D. Activation of endoplasmic reticulum stress promotes autophagy and apoptosis and reverses chemoresistance of human small cell lung cancer cells by inhibiting the PI3K/AKT/mTOR signaling pathway. Oncotarget 2016, 7, 76827–76839. [Google Scholar] [CrossRef]
- Merlot, A.; Shafie, N.; Yu, Y.; Richardson, V.; Jansson, P.J.; Sahni, S.; Lane, D.J.; Kovacevic, D.S.; Richardson, D.R. Mechanism of the induction of endoplasmic reticulum stress by the anti-cancer agent, di-2-pyridylketone 4,4-dimethyl-3-thiosemicarbazone (Dp44mT): Activation of PERK/eIF2a, IRE1a, ATF6 and calmodulin kinase. Biochem. Pharmacol. 2016, 109, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.H.; Chiou, H.L.; Lin, C.L.; Lin, C.Y.; Yang, S.F.; Hsieh, Y.H. Induction of endoplasmic reticulum stress and mitochondrial dysfunction dependent apoptosis signaling pathway in human renal cancer cells by Norcantharidin. Oncotarget 2018, 9, 4787–4797. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, C.; Li, R.; Zhang, L.; Tian, J.K. TEOA, a triterpenoid from Actinidia eriantha, induces autophagy in SW620 cells via endoplasmic reticulum stress and ROS-dependent mitophagy. Arch. Pharm. Res. 2017, 40, 1–13. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 | ||||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||
| H22 cells | AAEE | 59.16 | 33.40 | 14.71 |
| AAEE-Pe | 32.91 | 29.64 | 23.38 | |
| AAEE-Ea | 97.76 | 43.57 | 25.69 | |
| BEL-7404 cells | AAEE | 89.86 | 56.86 | 60.39 |
| AAEE-Pe | 98.71 | 75.88 | 89.51 | |
| AAEE-Ea | 171.7 | 82.04 | 71.97 | |
| AAEE | AAEE-Pe | AAEE-Ea | |
|---|---|---|---|
| Polysaccharides | 31.15% | 0.68% | 9.62% |
| Flavonoids | 16.89% | 10.26% | 24.97% |
| Triterpenes | 22.57% | 31.59% | 29.89% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.; Xia, L.; Ziyayiding, D.; Chen, Q.; Liu, R.; Xu, X.; Li, J. The Extracts of Artemisia absinthium L. Suppress the Growth of Hepatocellular Carcinoma Cells through Induction of Apoptosis via Endoplasmic Reticulum Stress and Mitochondrial-Dependent Pathway. Molecules 2019, 24, 913. https://doi.org/10.3390/molecules24050913
Wei X, Xia L, Ziyayiding D, Chen Q, Liu R, Xu X, Li J. The Extracts of Artemisia absinthium L. Suppress the Growth of Hepatocellular Carcinoma Cells through Induction of Apoptosis via Endoplasmic Reticulum Stress and Mitochondrial-Dependent Pathway. Molecules. 2019; 24(5):913. https://doi.org/10.3390/molecules24050913
Chicago/Turabian StyleWei, Xianxian, Lijie Xia, Dilinigeer Ziyayiding, Qiuyan Chen, Runqing Liu, Xiaoyu Xu, and Jinyao Li. 2019. "The Extracts of Artemisia absinthium L. Suppress the Growth of Hepatocellular Carcinoma Cells through Induction of Apoptosis via Endoplasmic Reticulum Stress and Mitochondrial-Dependent Pathway" Molecules 24, no. 5: 913. https://doi.org/10.3390/molecules24050913
APA StyleWei, X., Xia, L., Ziyayiding, D., Chen, Q., Liu, R., Xu, X., & Li, J. (2019). The Extracts of Artemisia absinthium L. Suppress the Growth of Hepatocellular Carcinoma Cells through Induction of Apoptosis via Endoplasmic Reticulum Stress and Mitochondrial-Dependent Pathway. Molecules, 24(5), 913. https://doi.org/10.3390/molecules24050913