Comparison among Activities and Isoflavonoids from Pueraria thunbergiana Aerial Parts and Root

Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolic Contents
2.2. Antioxidant Activity
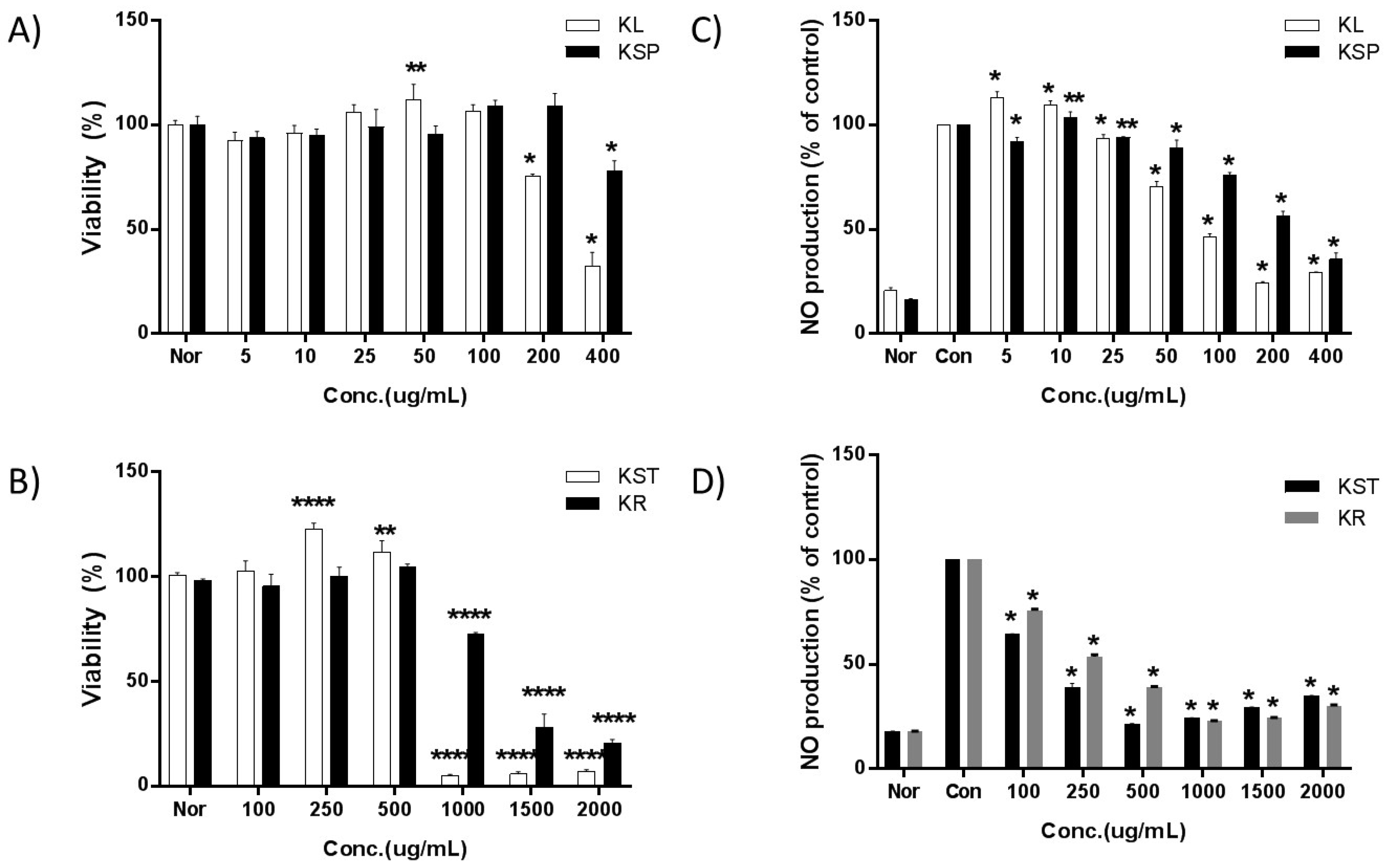
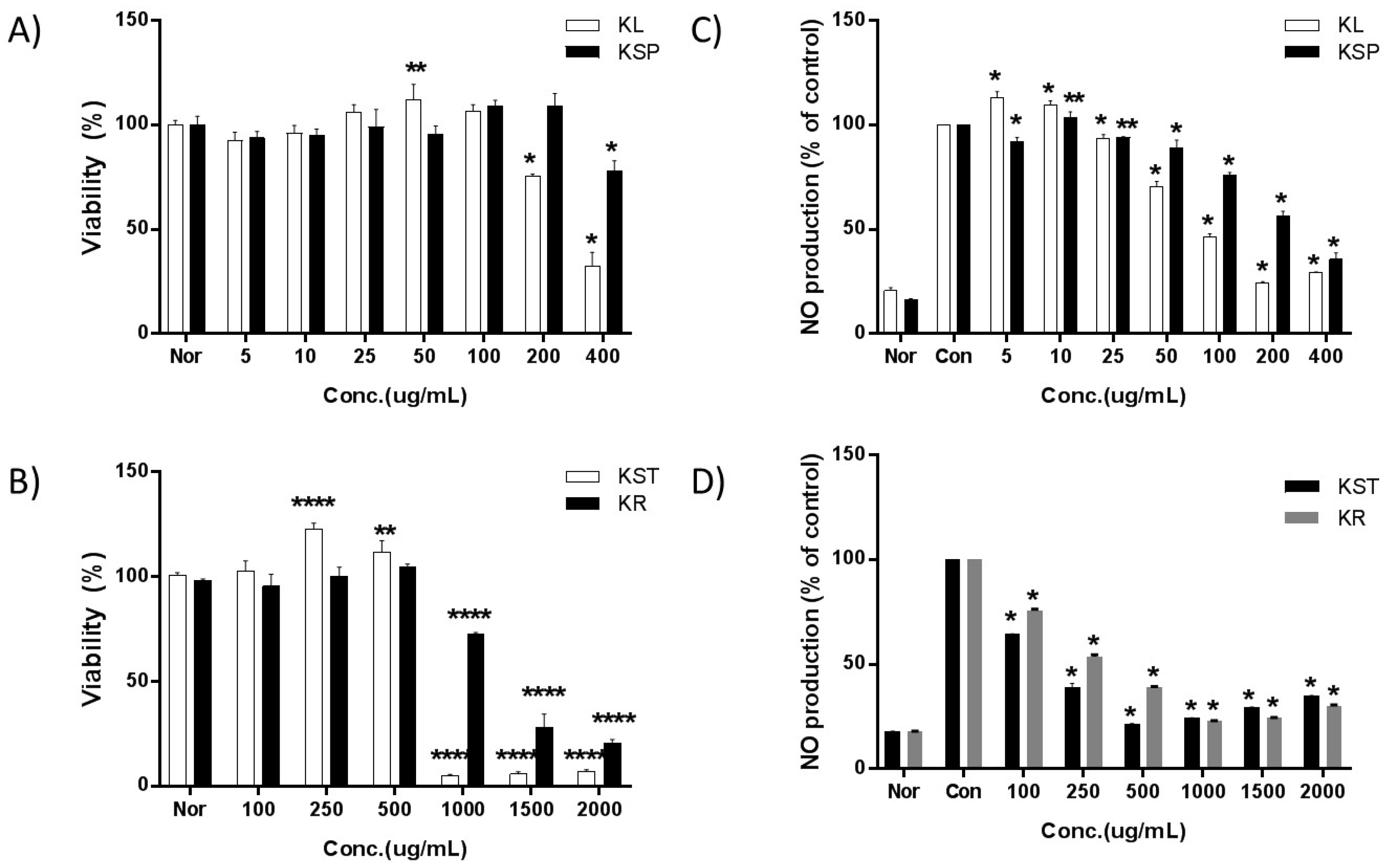
2.3. Cell Viability
2.4. Effect of Kudzu Extracts by Part on NO Production
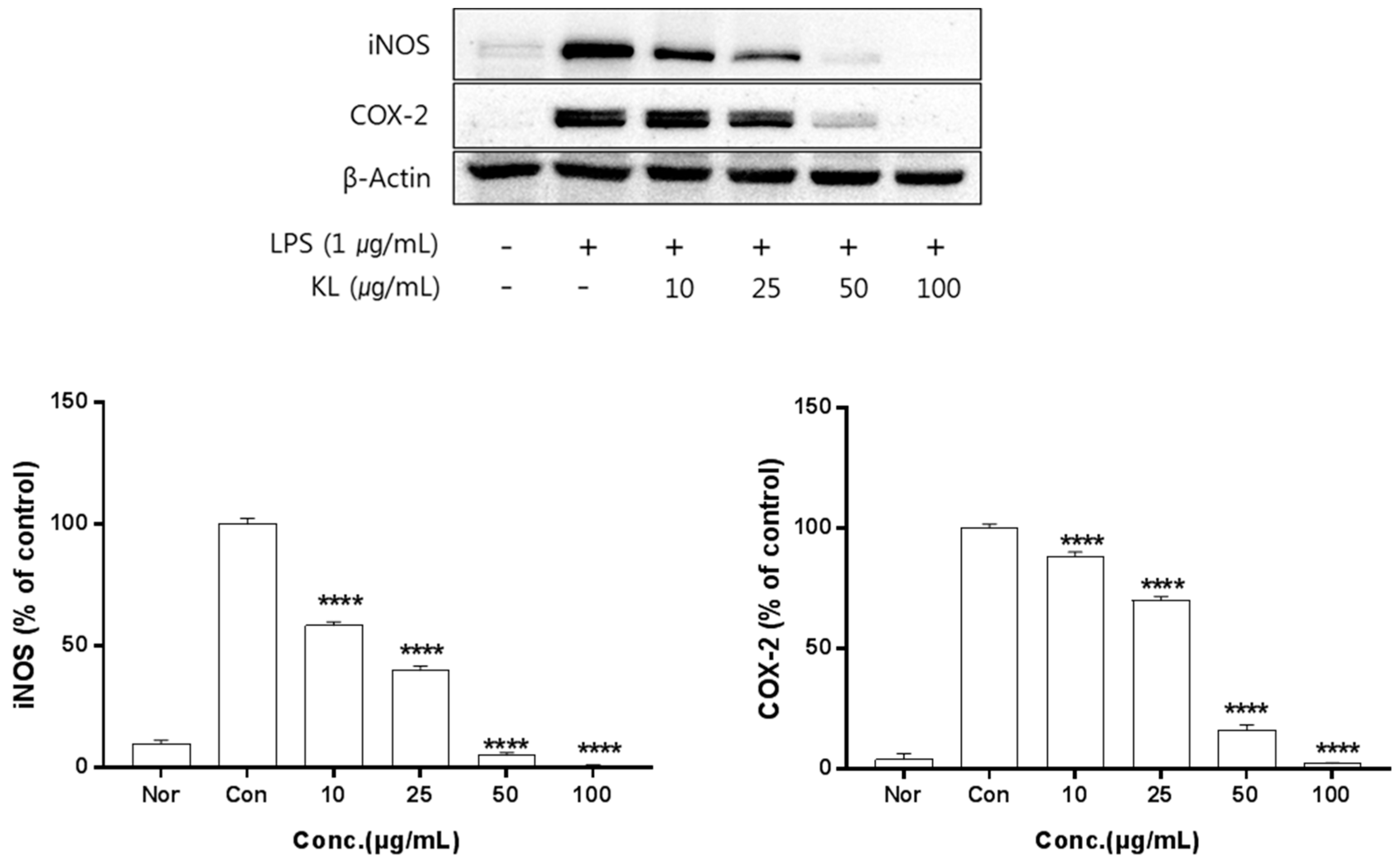
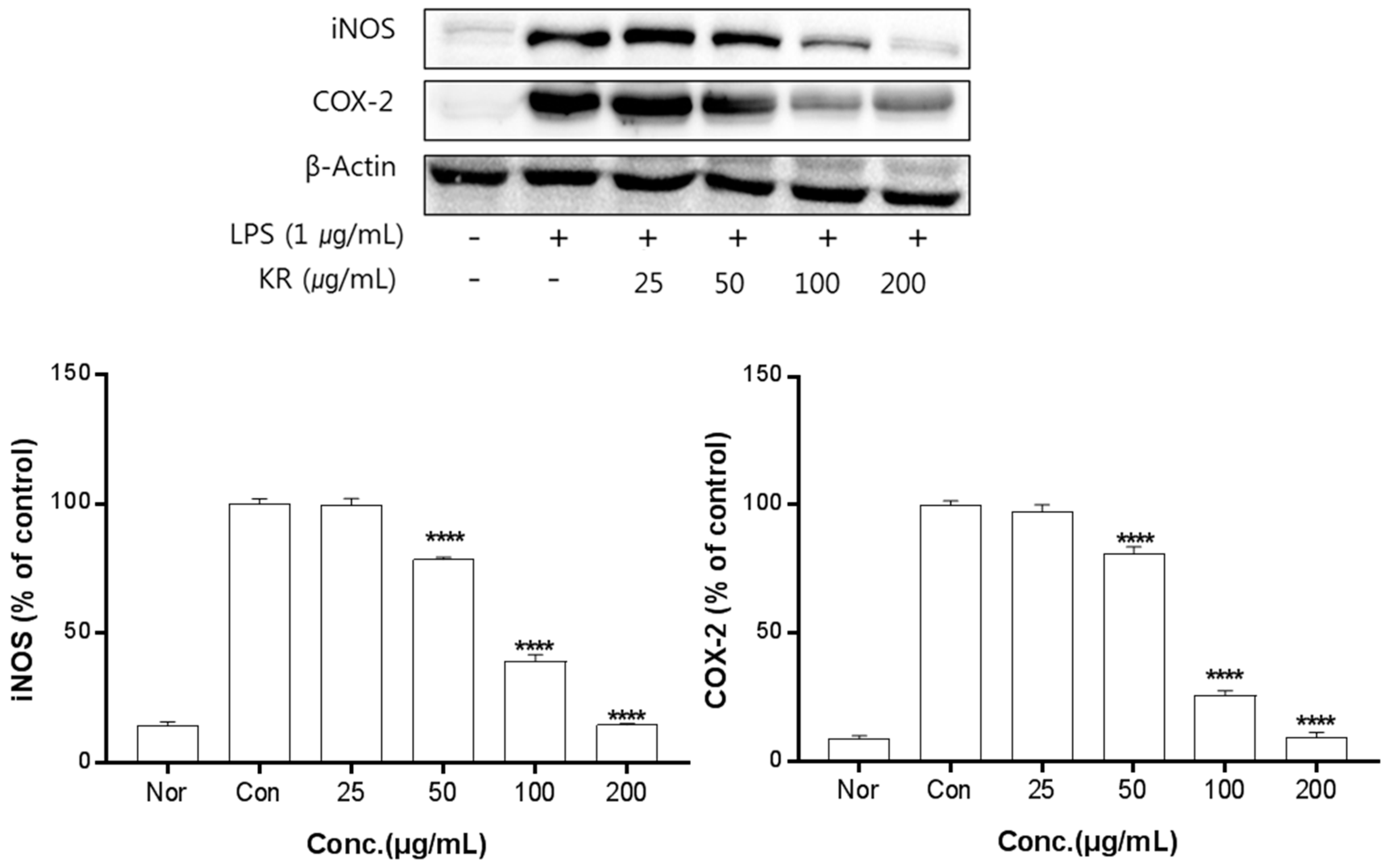
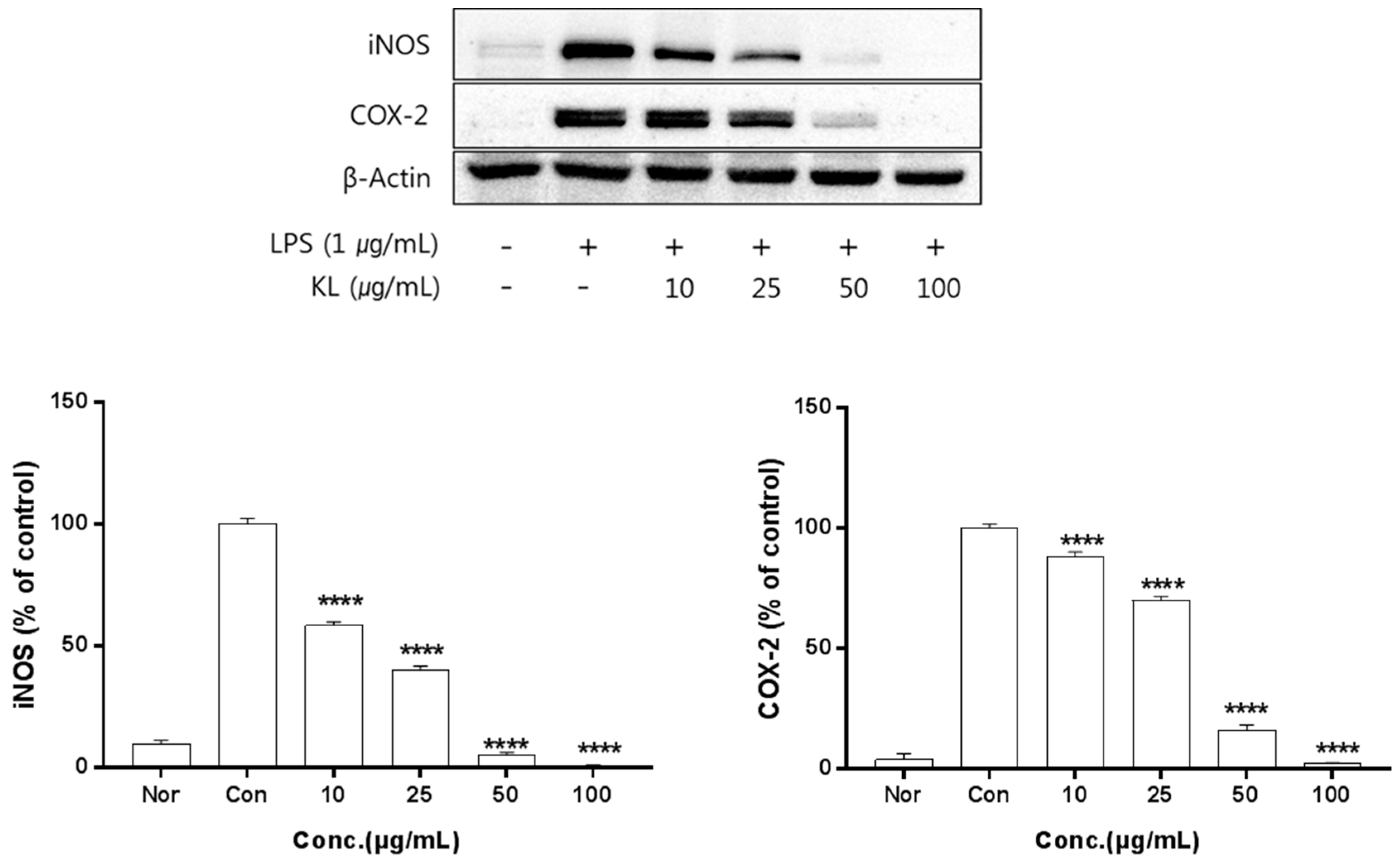
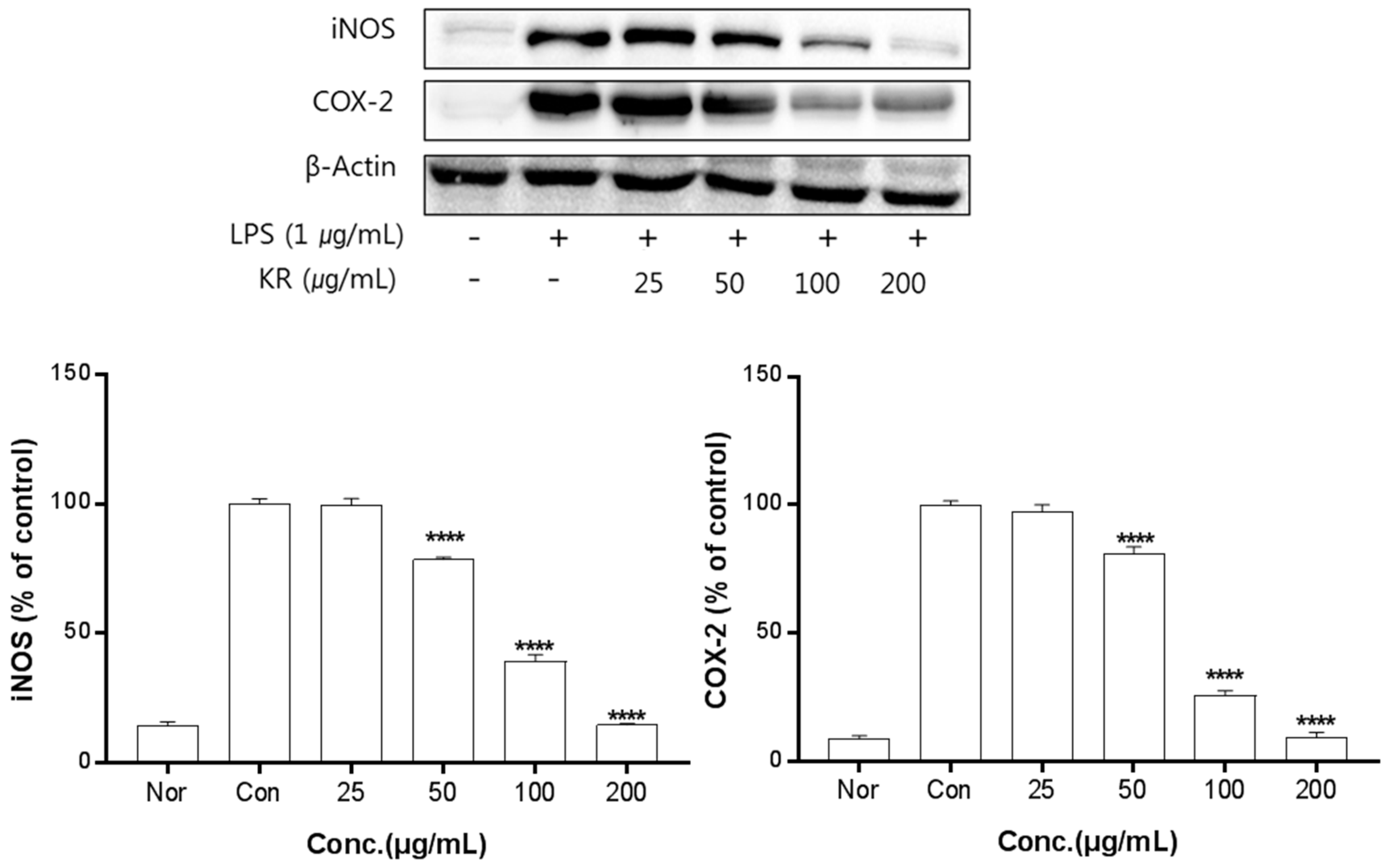
2.5. Anti-Inflammatory Activity of Kudzu Extracts by Part
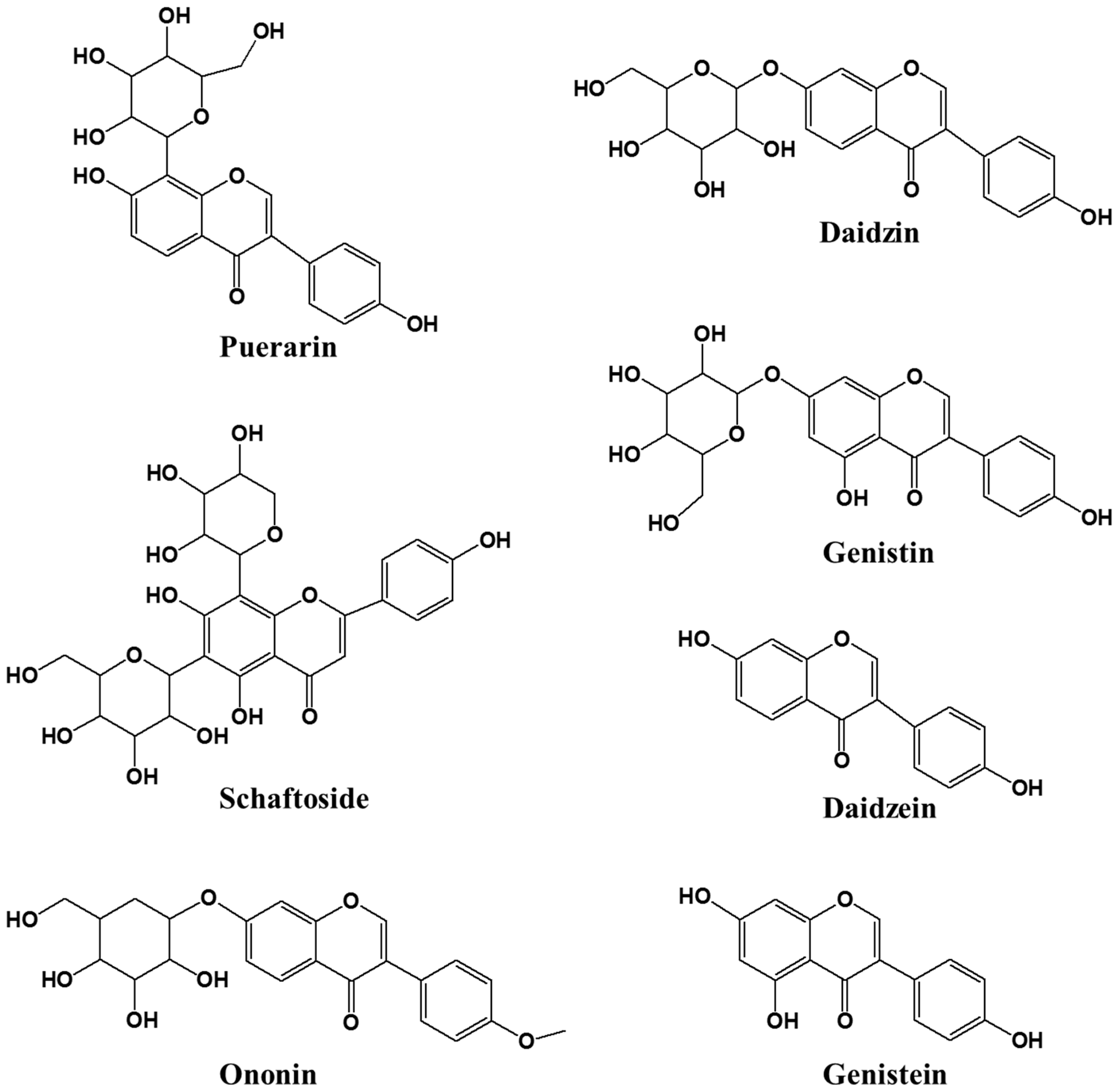
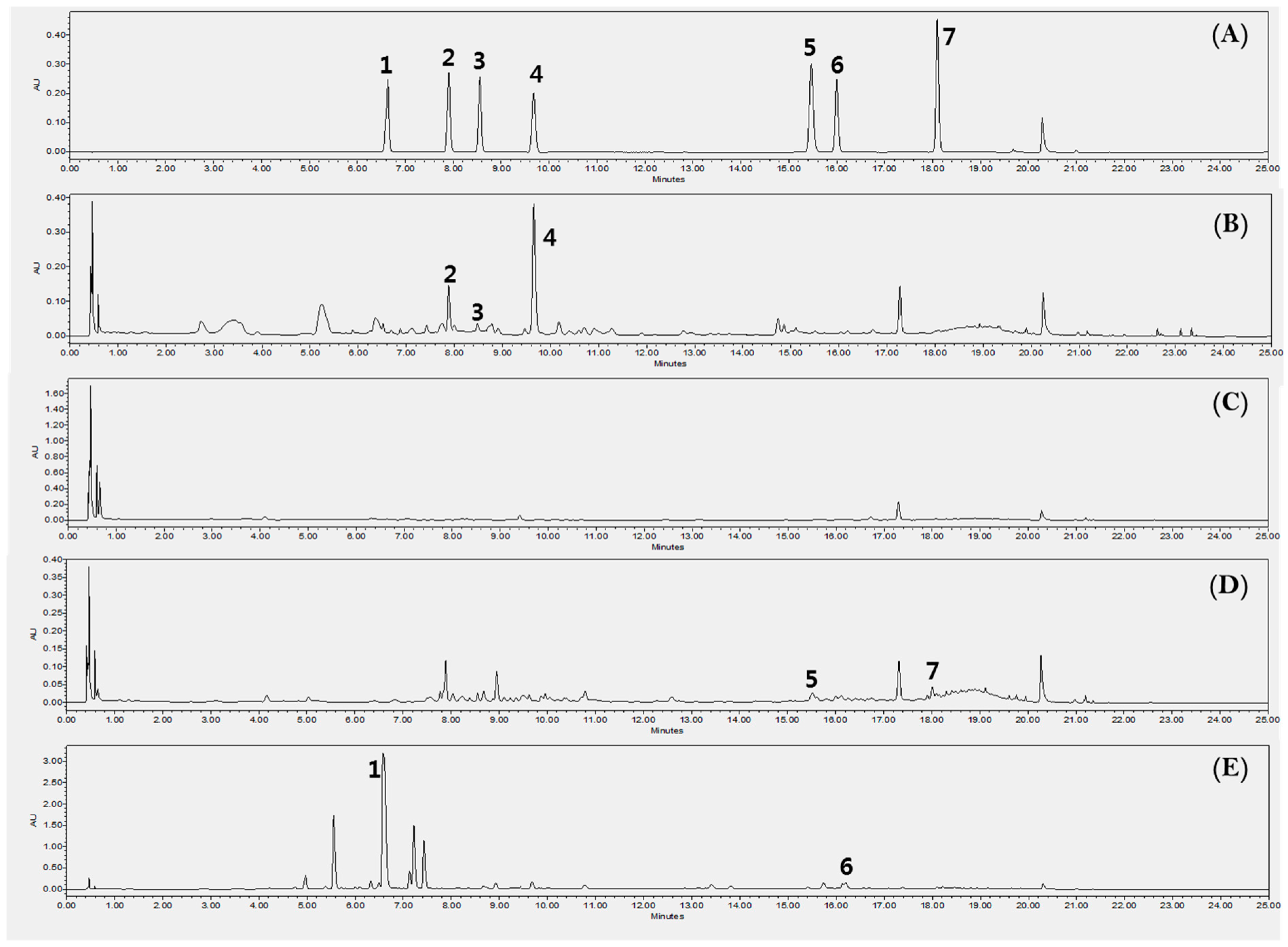
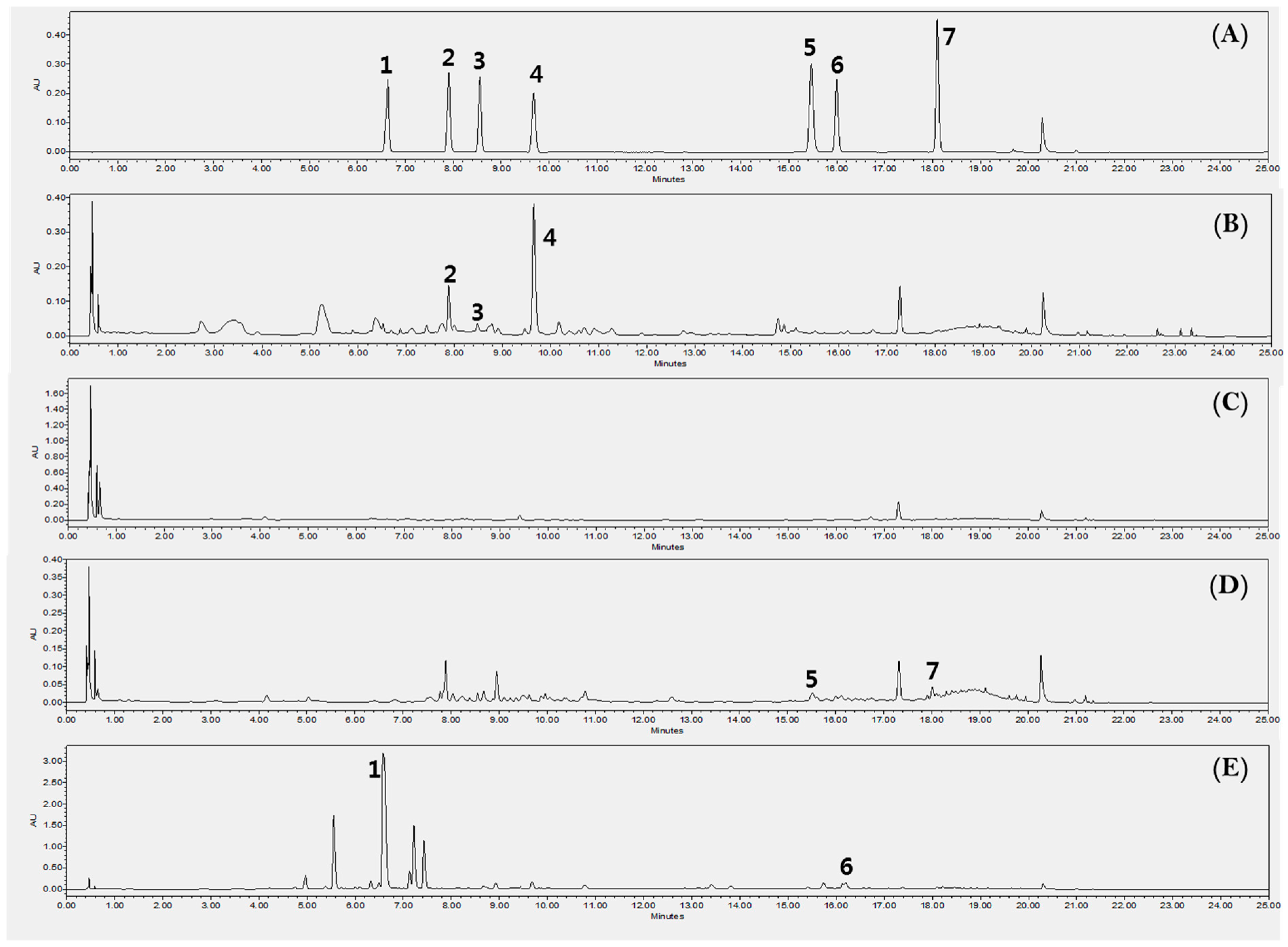
2.6. HPLC for Quantitative Analysis
3. Materials and Methods
3.1. Materials
3.2. Chemicals
3.3. Total Phenolic Content Assay (TPC)
3.4. DPPH Scavenging Activity Assay
- DPPH FRS activity (%) = (1 − ) ×100
- A1: The value for solvent absorbance with DPPH solution
- A2: The value for solvent absorbance without DPPH solution
- A3: The value for sample absorbance with DPPH solution
- A4: The value for sample absorbance without DPPH solution
3.5. ABTS Radical Scavenging Activity Assay
- ABTS+• FRS activity (%) = (1 − ) ×100
- A1: The value for solvent absorbance with ABTS+ radical solution
- A2: The value for solvent absorbance without ABTS+ radical solution
- A3: The value for sample absorbance with ABTS+ radical solution
- A4: The value for sample absorbance without ABTS+ radical solution
3.6. Xanthine Oxidase Inhibitory Activity Assay
3.7. Cell Culture and Viability
3.8. Immunoblotting Analysis
3.9. NO Production Assay
3.10. High-Performance Liquid Chromatography Analysis
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yuan, R.; Lin, Y. Traditional Chinese medicine: An approach to scientific proof and clinical validation. Pharmacol. Ther. 2000, 86, 191–198. [Google Scholar] [CrossRef]
- Chen, Z.F.L.; Huang, Q.; Li, Y.Y.; Zhang, Y.; Ren, Y.; Li, K.S.; Fu, Z.J.; Xu, S.Q. Nutrient starved incubation conditions enhance pyrazinamide activity against Mycobacterium tuberculosis. Zhonghua Jie He He Hu Xi Za Zhi 2007, 30, 359–362. (In Chinese) [Google Scholar] [PubMed]
- Prasain, J.K.; Peng, N.; Rajbhandari, R.; Wyss, J.M. The Chinese Pueraria root extract (Pueraria lobata) ameliorates impaired glucose and lipid metabolism in obese mice. Phytomedicine 2012, 20, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Li, G.Q.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K. Kudzu root: Traditional uses and potential medicinal benefits in diabetes and cardiovascular diseases. J. Ethnopharmacol. 2011, 134, 584–607. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.L.; Li, M.; Xie, S.H.; Song, Y.; Zou, Z.M.; Zhu, C.Y.; Qi, Y. Antihypertensive effect of total flavone extracts from Puerariae Radix. J. Ethnopharmacol. 2011, 133, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, X.; Gao, Y.; Guo, G.; Xu, C.; Li, G.; Liu, S.; Huang, A.; Tu, G.; Peng, H.; et al. Effects of puerarin on the inflammatory role of burn-related procedural pain mediated by P2X(7) receptors. Burns 2013, 39, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.M.; Zhao, C.S.; He, Z.G. Assessment of Pueraria lobata isoflavone with self-microemulsifying drug delivery systems in vitro and in vivo. Zhong Yao Cai 2007, 30, 684–687. (In Chinese) [Google Scholar] [PubMed]
- Xue, C.; Zhang, J.; Jing, W.; Wang, Y.; Si, N.; Liu, A. Determination of five components in Pueraria labta decoction with reference extraction method. Zhongguo Zhong Yao Za Zhi 2012, 37, 2388–2391. (In Chinese) [Google Scholar] [PubMed]
- Tanaka, T.; Tang, H.J.; Yu, F.N.; Michihara, S.; Uzawa, Y.; Zaima, N.; Moriyama, T.; Kawamura, Y. Kudzu (Pueraria lobata) Vine Ethanol Extracts Improve Ovariectomy-Induced Bone Loss in Female Mice. J. Agric. Food Chem. 2011, 59, 13230–13237. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.H.; Li, X.L.; Mei, Z.G.; Xiong, L.; Mei, Q.X.; Wang, J.F.; Tan, L.J.; Yang, S.B.; Feng, Z.T. Efficacy and safety of puerarin injection in curing acute ischemic stroke: A meta-analysis of randomized controlled trials. Medicine (Baltimore) 2017, 96, e5803. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Y.; Han, J.; Zai, D.; Ji, M.; Cheng, W.; Xu, L.; Yang, L.; He, M.; Ni, J.; et al. Puerarin suppresses invasion and vascularization of endometriosis tissue stimulated by 17beta-estradiol. PLoS ONE 2011, 6, e25011. [Google Scholar]
- Zou, Y.; Hong, B.; Fan, L.; Zhou, L.; Liu, Y.; Wu, Q.; Zhang, X.; Dong, M. Protective effect of puerarin against beta-amyloid-induced oxidative stress in neuronal cultures from rat hippocampus: Involvement of the GSK-3beta/Nrf2 signaling pathway. Free Radic Res. 2013, 47, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.D.; Li, C.Y.; Cui, W.P.; Guo, Q.Y.; Dong, C.Q.; Zou, H.B.; Liu, S.J.; Dong, W.P.; Miao, L.N. Review of Herbal Traditional Chinese Medicine for the Treatment of Diabetic Nephropathy. J. Diabetes Res. 2016, 2016, 5749857. [Google Scholar] [CrossRef] [PubMed]
- Strong, A.L.; Ohlstein, J.F.; Jiang, Q.; Zhang, Q.; Zheng, S.; Boue, S.M.; Elliott, S.; Gimble, J.M.; Burow, M.E.; Wang, G.; et al. Novel daidzein analogs enhance osteogenic activity of bone marrow-derived mesenchymal stem cells and adipose-derived stromal/stem cells through estrogen receptor dependent and independent mechanisms. Stem Cell Res. Ther. 2014, 5, 105. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Payton-Stewart, F.; Elliott, S.; Driver, J.; Rhodes, L.V.; Zhang, Q.; Zheng, S.; Bhatnagar, D.; Boue, S.M.; Collins-Burow, B.M.; et al. Effects of 7-O substitutions on estrogenic and anti-estrogenic activities of daidzein analogues in MCF-7 breast cancer cells. J. Med. Chem. 2010, 53, 6153–6163. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Woo, M.S.; Qin, L.; Ma, T.; Beltran, C.D.; Bao, Y.; Bailey, J.A.; Corbett, D.; Ratan, R.R.; Lahiri, D.K.; et al. Daidzein Augments Cholesterol Homeostasis via ApoE to Promote Functional Recovery in Chronic Stroke. J. Neurosci. 2015, 35, 15113–15126. [Google Scholar] [CrossRef] [PubMed]
- Bakari, S.; Ncir, M.; Felhi, S.; Hajlaoui, H.; Saoudi, M.; Gharsalah, N.; Kadri, A. Chemical composition and in vitro evaluation of total phenolic, flavonoid, and antioxidant properties of essential oil and solvent extract from the aerial parts of Teucrium polium grown in Tunisia. Food Sci. Biotechnol. 2015, 24, 1943–1949. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Esmat, A. Antioxidant and anti-inflammatory activities of the major phenolics from Zygophyllum simplex, L. J. Ethnopharmacol. 2017, 205, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Prochazkova, D.; Bousova, I.; Wilhelmova, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Bourgou, S.; Ksouri, R.; Bellila, A.; Skandrani, I.; Falleh, H.; Marzouk, B. Phenolic composition and biological activities of Tunisian Nigella sativa, L. shoots and roots. C. R. Biol. 2008, 331, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Filipiak-Szok, A.; Kurzawa, M.; Szlyk, E. Determination of anti-oxidant capacity and content of phenols, phenolic acids, and flavonols in Indian and European gooseberry. Chem. Pap. 2012, 66, 259–268. [Google Scholar] [CrossRef]
- Gujral, H.S.; Sharma, P.; Kumar, A.; Singh, B. Total Phenolic Content and Antioxidant Activity of Extruded Brown Rice. Int. J. Food Prop. 2012, 15, 301–311. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Hammond, R.A.; Hannon, R.; Frean, S.P.; Armstrong, S.J.; Flower, R.J.; Bryant, C.E. Endotoxin induction of nitric oxide synthase and cyclooxygenase-2 in equine alveolar macrophages. Am. J. Vet. Res. 1999, 60, 426–431. [Google Scholar] [PubMed]
- Rehman, M.U.; Yoshihisa, Y.; Miyamoto, Y.; Shimizu, T. The anti-inflammatory effects of platinum nanoparticles on the lipopolysaccharide-induced inflammatory response in RAW 264.7 macrophages. Inflamm. Res. 2012, 61, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Cheng, Y.J.; Chen, C.K.; Yang, B.C. Coexposure of lead- and lipopolysaccharide-induced liver injury in rats: Involvement of nitric oxide-initiated oxidative stress and TNF-alpha. Shock 2005, 23, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Bartsch, H. Chronic infections and inflammatory processes as cancer risk factors: Possible role of nitric oxide in carcinogenesis. Mutat. Res. 1994, 305, 253–264. [Google Scholar] [CrossRef]
- Dai, Z.J.; Ma, X.B.; Kang, H.F.; Gao, J.; Min, W.L.; Guan, H.T.; Diao, Y.; Lu, W.F.; Wang, X.J. Antitumor activity of the selective cyclooxygenase-2 inhibitor, celecoxib, on breast cancer in Vitro and in Vivo. Cancer Cell Int. 2012, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zong, J.; Zhou, H.; Bian, Z.Y.; Deng, W.; Dai, J.; Gan, H.W.; Yang, Z.; Li, H.; Tang, Q.Z. Puerarin attenuates pressure overload-induced cardiac hypertrophy. J. Cardiol. 2014, 63, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, E.; Jakobkiewicz-Banecka, J.; Tylki-Szymanska, A.; Liberek, A.; Maryniak, A.; Malinowska, M.; Czartoryska, B.; Puk, E.; Kloska, A.; Liberek, T.; et al. Genistin-rich soy isoflavone extract in substrate reduction therapy for Sanfilippo syndrome: An open-label, pilot study in 10 pediatric patients. Curr. Ther. Res. Clin. Exp. 2008, 69, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Farmakalidis, E.; Hathcock, J.N.; Murphy, P.A. Oestrogenic potency of genistin and daidzin in mice. Food Chem. Toxicol. 1985, 23, 741–745. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Kim, D.S.; Kim, S.H.; Kim, H.K. Anti-obesity activity, acute toxicity, and chemical constituents of aqueous and ethanol Viola mandshurica extracts. BMC Complement Altern. Med. 2017, 17, 297. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Yin, L.; Zhang, Y.; Fu, X.; Lu, J. Anti-inflammatory effects of ononin on lipopolysaccharide-stimulated RAW 264.7 cells. Mol. Immunol. 2017, 83, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Singh, A.P. In vitro antioxidant and free radical scavenging activity of Nardostachys jatamansi DC. J. Acupunct. Meridian Stud. 2012, 5, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.X.; Wang, Y.W.; Han, J.G. Determination of Polyphenols in Sorghum Grains by Near Infrared Reflectance Spectroscopy. Chin. J. Anal. Chem. 2009, 37, 1275–1280. [Google Scholar]
- Huang, M.H.; Huang, S.S.; Wang, B.S.; Wu, C.H.; Sheu, M.J.; Hou, W.C.; Lin, S.S.; Huang, G.J. Antioxidant and anti-inflammatory properties of Cardiospermum halicacabum and its reference compounds ex vivo and in vivo. J. Ethnopharmacol. 2011, 133, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.; Kartal, M.; Naz, Q.; Ejaz, A.; Yilmaz, G.; Kan, Y.; Konuklugil, B.; Sener, B.; Choudhary, M.I. Antioxidant and anticholinesterase evaluation of selected Turkish Salvia species. Food Chem. 2007, 103, 1247–1254. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | FRS50 (μg/mL) | |
|---|---|---|
| DPPH FRS Activity | ABTS+• FRS Activity | |
| KL | 437 ± 10.9 b | 121 ± 6.6 b |
| KST | 1,136 ± 14.2 e | 455.3 ± 17.1 e |
| KR | 582 ± 16.4 c | 138.0 ± 2.7 c |
| KSP | 755 ± 8.6 d | 341.7 ± 1.4 d |
| Control | 17 ± 0.3 a | 5.5 ± 1.0 a |
| Content (mg/g) | ||||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | SUM | |
| KL | 5.5 ± 0.3 | 0.8 ± 0.0 | 60.8 ± 1.1 | 66.3 ± 1.42 | ||||
| KST | 0.6 ± 0.1 | 0.8 ± 0.1 | 0.1 ± 0.0 | 2.3 ± 0.2 | ||||
| KR | 67.1 ± 2.2 | 4.3 ± 0.3 | 2.0 ± 0.2 | 73.4 ± 2.7 | ||||
| KSP | * | * | * | * | * | * | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, E.; Yoon, J.-M.; An, B.-J.; Lee, Y.M.; Cha, J.; Chi, G.-Y.; Kim, D.-S. Comparison among Activities and Isoflavonoids from Pueraria thunbergiana Aerial Parts and Root. Molecules 2019, 24, 912. https://doi.org/10.3390/molecules24050912
Son E, Yoon J-M, An B-J, Lee YM, Cha J, Chi G-Y, Kim D-S. Comparison among Activities and Isoflavonoids from Pueraria thunbergiana Aerial Parts and Root. Molecules. 2019; 24(5):912. https://doi.org/10.3390/molecules24050912
Chicago/Turabian StyleSon, Eunjung, Jong-Moon Yoon, Bong-Jeun An, Yun Mi Lee, Jimin Cha, Gyeong-Yup Chi, and Dong-Seon Kim. 2019. "Comparison among Activities and Isoflavonoids from Pueraria thunbergiana Aerial Parts and Root" Molecules 24, no. 5: 912. https://doi.org/10.3390/molecules24050912
APA StyleSon, E., Yoon, J.-M., An, B.-J., Lee, Y. M., Cha, J., Chi, G.-Y., & Kim, D.-S. (2019). Comparison among Activities and Isoflavonoids from Pueraria thunbergiana Aerial Parts and Root. Molecules, 24(5), 912. https://doi.org/10.3390/molecules24050912