
New Metabolic Influencer on Oxytocin Release: The Ghrelin


 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
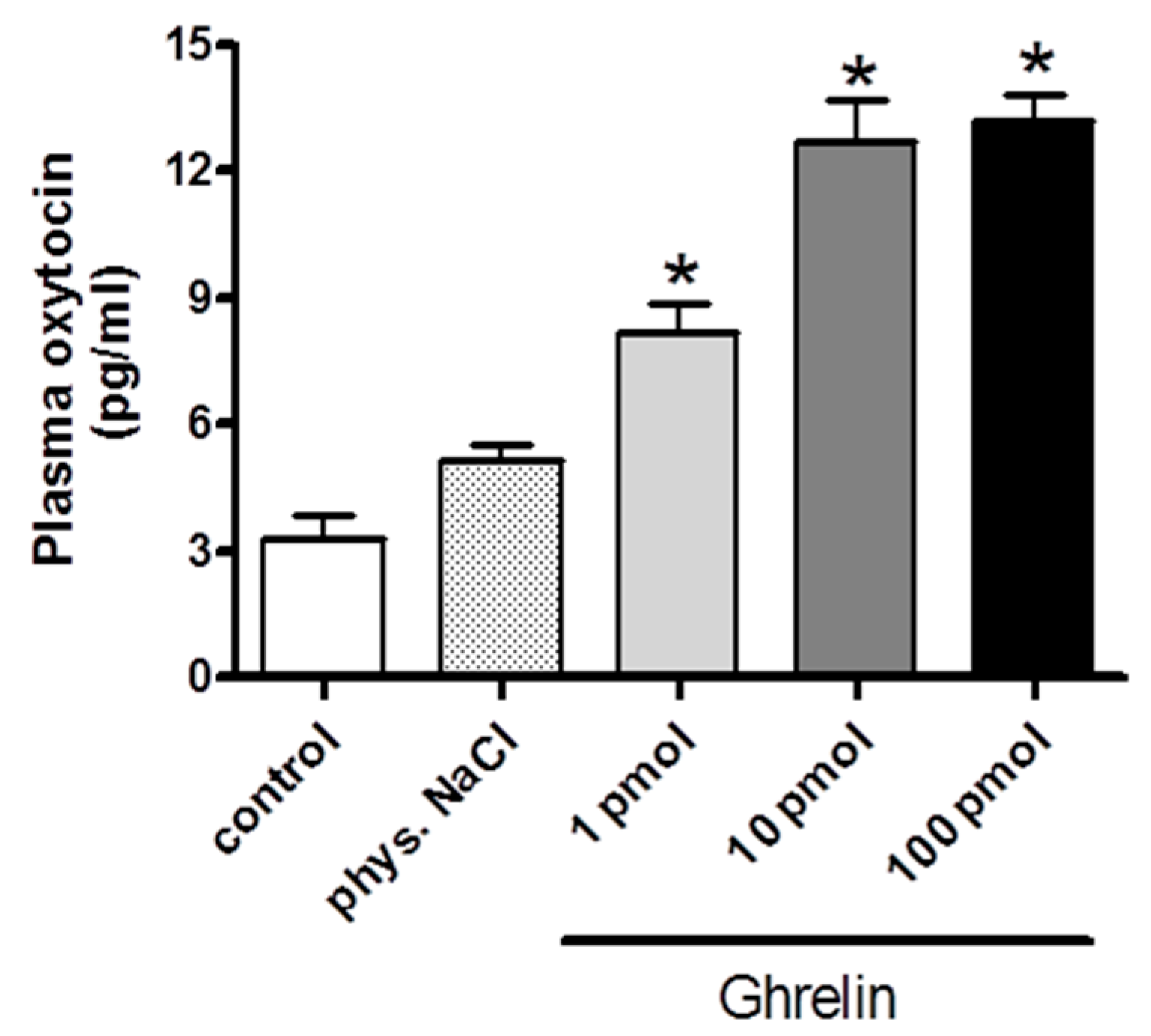
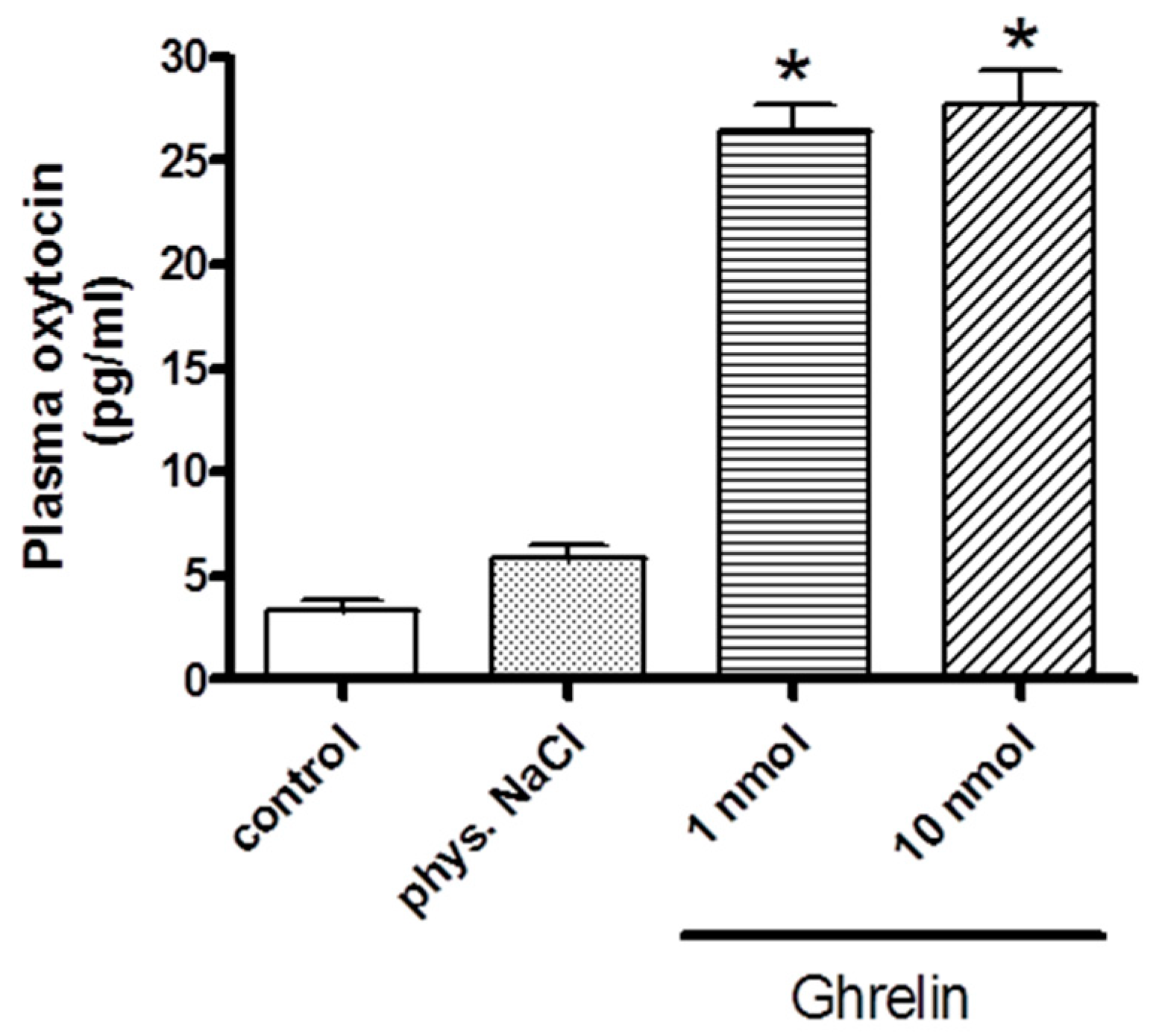
2.1. Effects of Centrally (i.c.v.) and Systemically (i.v.) Administrated Ghrelin on Oxytocin Secretion
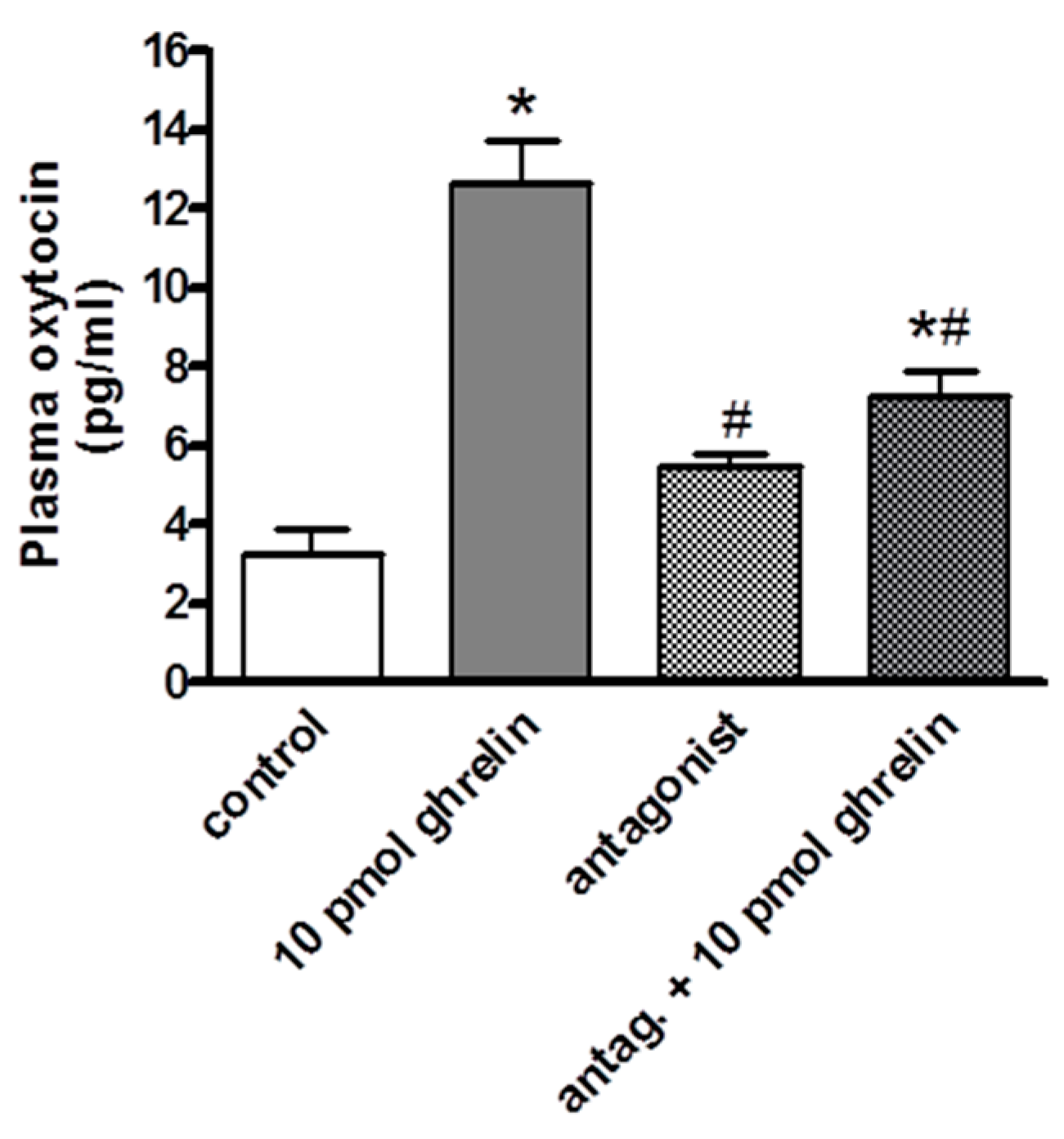
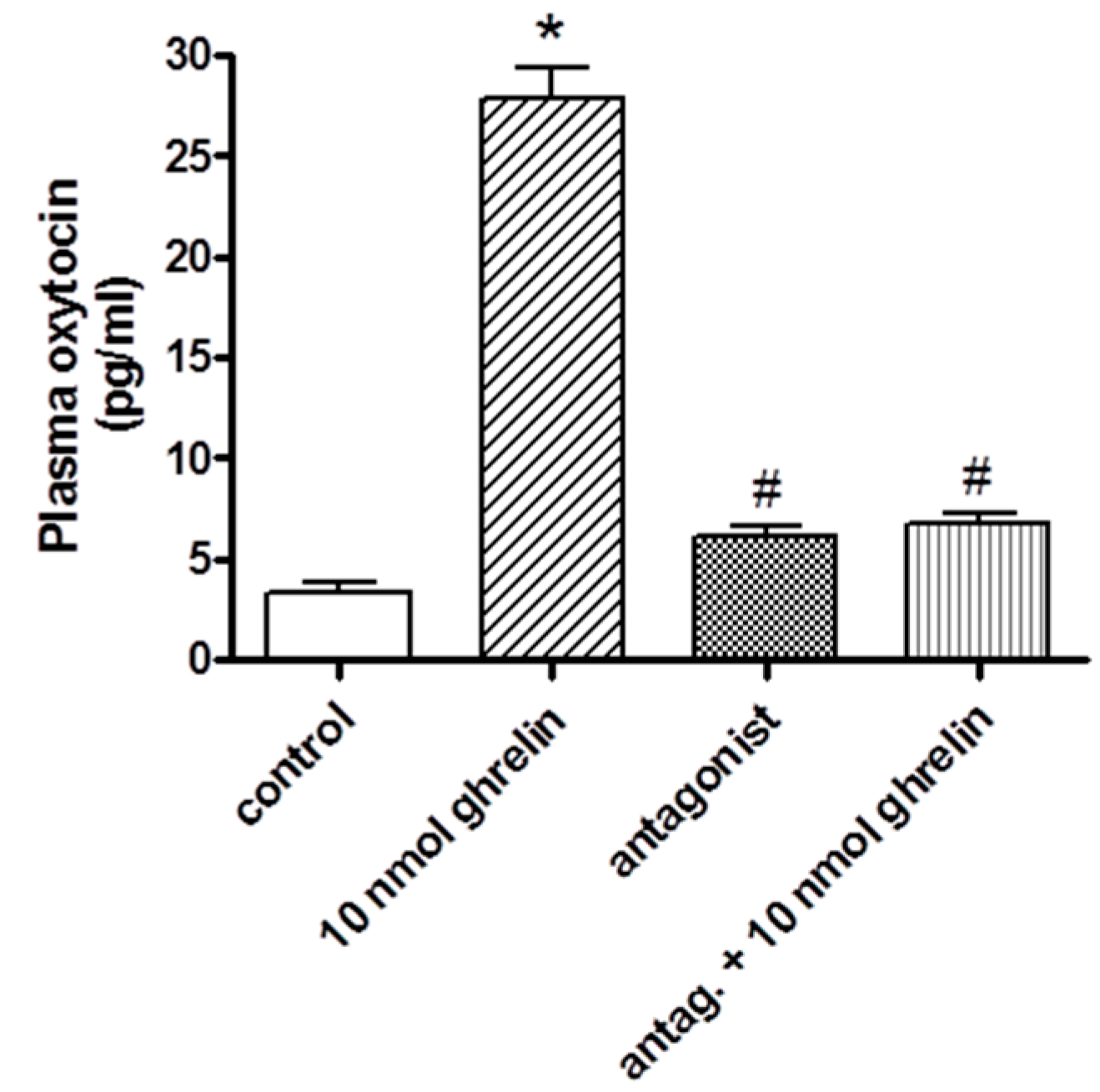
2.2. Effects of Centrally (i.c.v.) and Systemically (i.v.) Administrated Ghrelin Antagonist on Oxytocin Secretion
2.3. Behavioral Changes at the End of the Experimental Period
3. Discussion
4. Materials and Methods
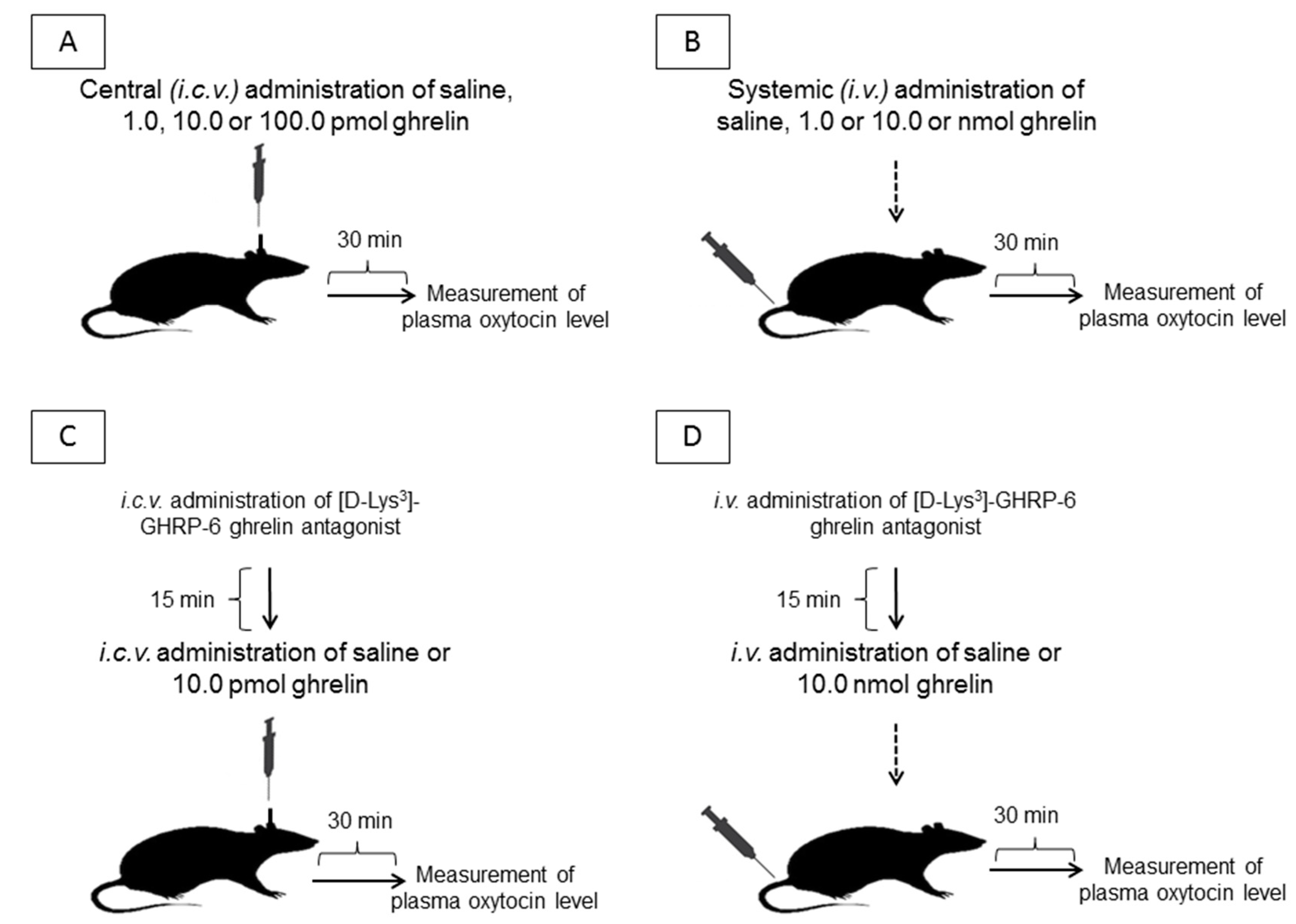
4.1. Experimental Protocol
4.2. Plasma Oxytocin Determination
4.3. Protein Measurement
4.4. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Coll, A.P.; Yeo, G.S. The hypothalamus and metabolism: Integrating signals to control energy and glucose homeostasis. Curr. Opin. Pharmacol. 2013, 13, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Cong, W.N.; Golden, E.; Pantaleo, N.; White, C.M.; Maudsley, S.; Martin, B. Ghrelin receptor signaling: A promising therapeutic target for metabolic syndrome and cognitive dysfunction. CNS Neurol. Disord. Drug Targets 2010, 9, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Poykko, S.M.; Kellokoski, E.; Horkko, S.; Kauma, H.; Kesaniemi, Y.A.; Ukkola, O. Low plasma ghrelin is associated with insulin resistance, hypertension, and the prevalence of type 2 diabetes. Diabetes 2003, 52, 2546–2553. [Google Scholar] [CrossRef] [PubMed]
- Barazzoni, R.; Zanetti, M.; Ferreira, C.; Vinci, P.; Pirulli, A.; Mucci, M.; Dore, F.; Fonda, M.; Ciocchi, B.; Cattin, L.; et al. Relationships between desacylated and acylated ghrelin and insulin sensitivity in the metabolic syndrome. J. Clin. Endocrinol. Metab. 2007, 92, 3935–3940. [Google Scholar] [CrossRef] [PubMed]
- Galfi, M.; Radács, M.; Molnár, Z.; Budai, I.; Tóth, G.; Pósa, A.; Kupai, K.; Szalai, Z.; Szabó, R.; Molnár, H.A.; et al. Ghrelin-Induced Enhancement of Vasopressin and Oxytocin Secretion in Rat Neurohypophyseal Cell Cultures. J. Mol. Neurosci. 2016, 60, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.M.; Blevins, J.E. Coming full circle: Contributions of central and peripheral oxytocin actions to energy balance. Endocrinology 2013, 154, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Klement, J.; Ott, V.; Rapp, K.; Brede, S.; Piccinini, F.; Cobelli, C.; Lehnert, H.; Hallschmid, M. Oxytocin Improves beta-Cell Responsivity and Glucose Tolerance in Healthy Men. Diabetes 2017, 66, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, C.; Chen, Q.; Chen, X.; Xu, Z.; Wu, J.; Cai, D. Treatment of obesity and diabetes using oxytocin or analogs in patients and mouse models. PLoS ONE 2013, 8, e61477. [Google Scholar] [CrossRef] [PubMed]
- Bali, A.; Jaggi, A.S. An Integrative Review on Role and Mechanisms of Ghrelin in Stress, Anxiety and Depression. Curr. Drug Targets 2016, 17, 495–507. [Google Scholar] [CrossRef]
- Olszewski, P.K.; Bomberg, E.M.; Martell, A.; Grace, M.K.; Levine, A.S. Intraventricular ghrelin activates oxytocin neurons: Implications in feeding behavior. Neuroreport 2007, 18, 499–503. [Google Scholar] [CrossRef]
- Carmean, C.M.; Cohen, R.N.; Brady, M.J. Systemic regulation of adipose metabolism. Biochim. Biophys. Acta 2014, 1842, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Purkayastha, S. A New Horizon: Oxytocin as a Novel Therapeutic Option for Obesity and Diabetes. Drug Discov. Today Dis. Mech. 2013, 10, e63–e68. [Google Scholar] [CrossRef] [PubMed]
- Camerino, C. Low sympathetic tone and obese phenotype in oxytocin-deficient mice. Obesity (Silver Spring) 2009, 17, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Deblon, N.; Veyrat-Durebex, C.; Bourgoin, L.; Caillon, A.; Bussier, A.L.; Petrosino, S.; Piscitelli, F.; Legros, J.J.; Geenen, V.; Foti, M.; et al. Mechanisms of the anti-obesity effects of oxytocin in diet-induced obese rats. PLoS ONE 2011, 6, e25565. [Google Scholar] [CrossRef] [PubMed]
- Ott, V.; Finlayson, G.; Lehnert, H.; Heitmann, B.; Heinrichs, M.; Born, J.; Hallschmid, M. Oxytocin reduces reward-driven food intake in humans. Diabetes 2013, 62, 3418–3425. [Google Scholar] [CrossRef] [PubMed]
- Lawson, E.A. The effects of oxytocin on eating behaviour and metabolism in humans. Nat. Rev. Endocrinol. 2017, 13, 700–709. [Google Scholar] [CrossRef]
- Vila, G.; Riedl, M.; Resl, M.; van der Lely, A.J.; Hofland, L.J.; Clodi, M.; Luger, A. Systemic administration of oxytocin reduces basal and lipopolysaccharide-induced ghrelin levels in healthy men. J. Endocrinol. 2009, 203, 175–179. [Google Scholar] [CrossRef]
- Fry, M.; Ferguson, A.V. Ghrelin: Central nervous system sites of action in regulation of energy balance. Int. J. Pept. 2010, 2010, 616757. [Google Scholar] [CrossRef]
- Olszewski, P.K.; Grace, M.K.; Billington, C.J.; Levine, A.S. Hypothalamic paraventricular injections of ghrelin: Effect on feeding and c-Fos immunoreactivity. Peptides 2003, 24, 919–923. [Google Scholar] [CrossRef]
- Galfi, M.; Radács, M.; Juhász, A.; László, F.; Molnár, A.; László, F.A. Serotonin-induced enhancement of vasopressin and oxytocin secretion in rat neurohypophyseal tissue culture. Regul. Pept. 2005, 127, 225–231. [Google Scholar] [CrossRef]
- Galfi, M.; Janáky, T.; Tóth, R.; Prohászka, G.; Juhász, A.; Varga, C.; László, F.A. Effects of dopamine and dopamine-active compounds on oxytocin and vasopressin production in rat neurohypophyseal tissue cultures. Regul. Pept. 2001, 98, 49–54. [Google Scholar] [CrossRef]
- Patel, K.; Dixit, V.D.; Lee, J.H.; Kim, J.W.; Schaffer, E.M.; Nguyen, D.; Taub, D.D. Identification of ghrelin receptor blocker, D-[Lys3] GHRP-6 as a CXCR4 receptor antagonist. Int. J. Biol. Sci. 2012, 8, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, R.; Ménesi, R.; H. Molnár, A.; Szalai, Z.; Daruka, L.; Tóth, G.; Gardi, J.; Gálfi, M.; Börzsei, D.; Kupai, K.; et al. New Metabolic Influencer on Oxytocin Release: The Ghrelin. Molecules 2019, 24, 735. https://doi.org/10.3390/molecules24040735
Szabó R, Ménesi R, H. Molnár A, Szalai Z, Daruka L, Tóth G, Gardi J, Gálfi M, Börzsei D, Kupai K, et al. New Metabolic Influencer on Oxytocin Release: The Ghrelin. Molecules. 2019; 24(4):735. https://doi.org/10.3390/molecules24040735
Chicago/Turabian StyleSzabó, Renáta, Rudolf Ménesi, Andor H. Molnár, Zita Szalai, Lejla Daruka, Gábor Tóth, János Gardi, Márta Gálfi, Denise Börzsei, Krisztina Kupai, and et al. 2019. "New Metabolic Influencer on Oxytocin Release: The Ghrelin" Molecules 24, no. 4: 735. https://doi.org/10.3390/molecules24040735
APA StyleSzabó, R., Ménesi, R., H. Molnár, A., Szalai, Z., Daruka, L., Tóth, G., Gardi, J., Gálfi, M., Börzsei, D., Kupai, K., Juhász, A., Radács, M., László, F. A., Varga, C., & Pósa, A. (2019). New Metabolic Influencer on Oxytocin Release: The Ghrelin. Molecules, 24(4), 735. https://doi.org/10.3390/molecules24040735