
Amylase and Xylanase from Edible Fungus Neurospora intermedia: Production and Characterization

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results and Discussion
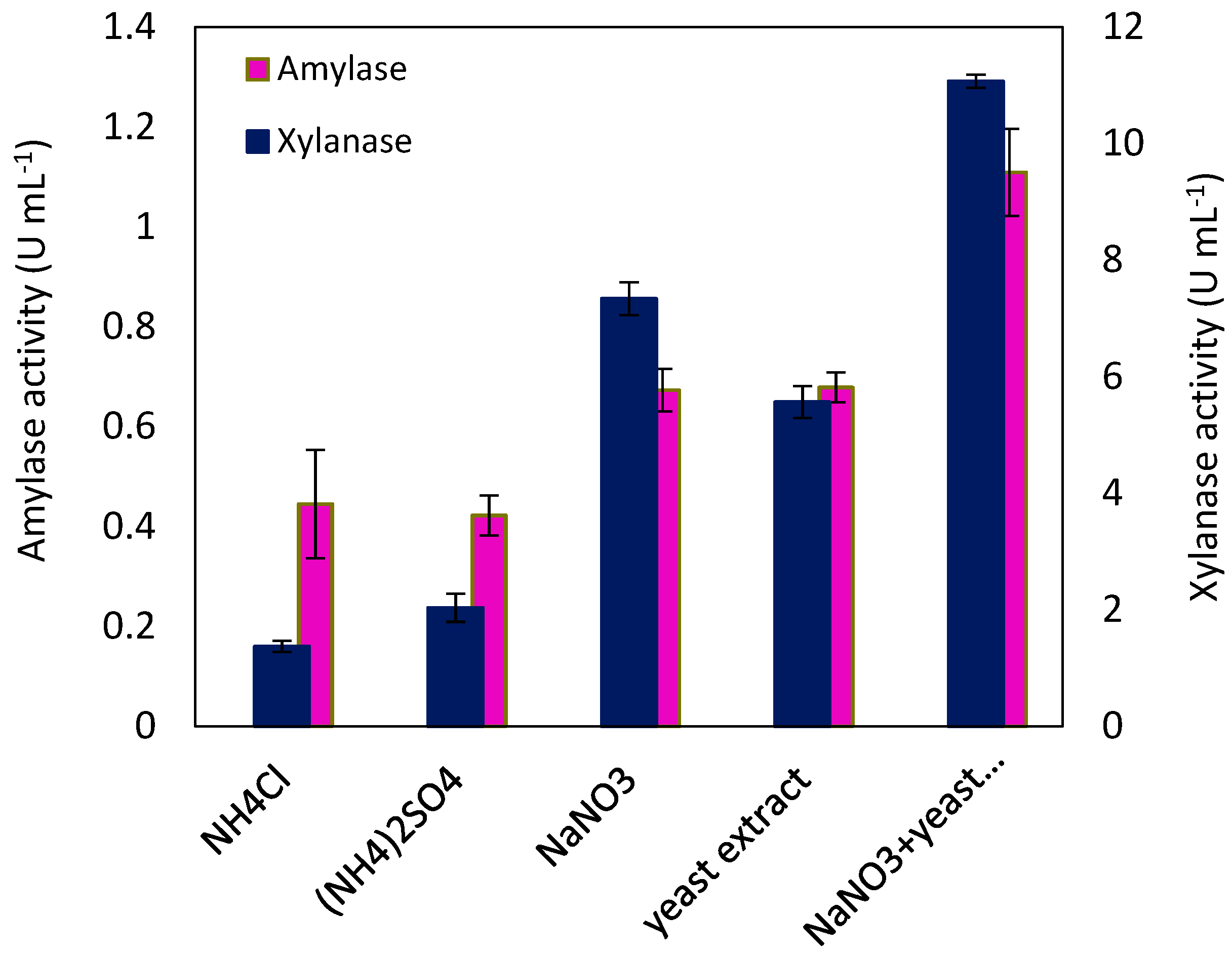
2.1. Optimization of Enzyme Production in a Shake-Flask System
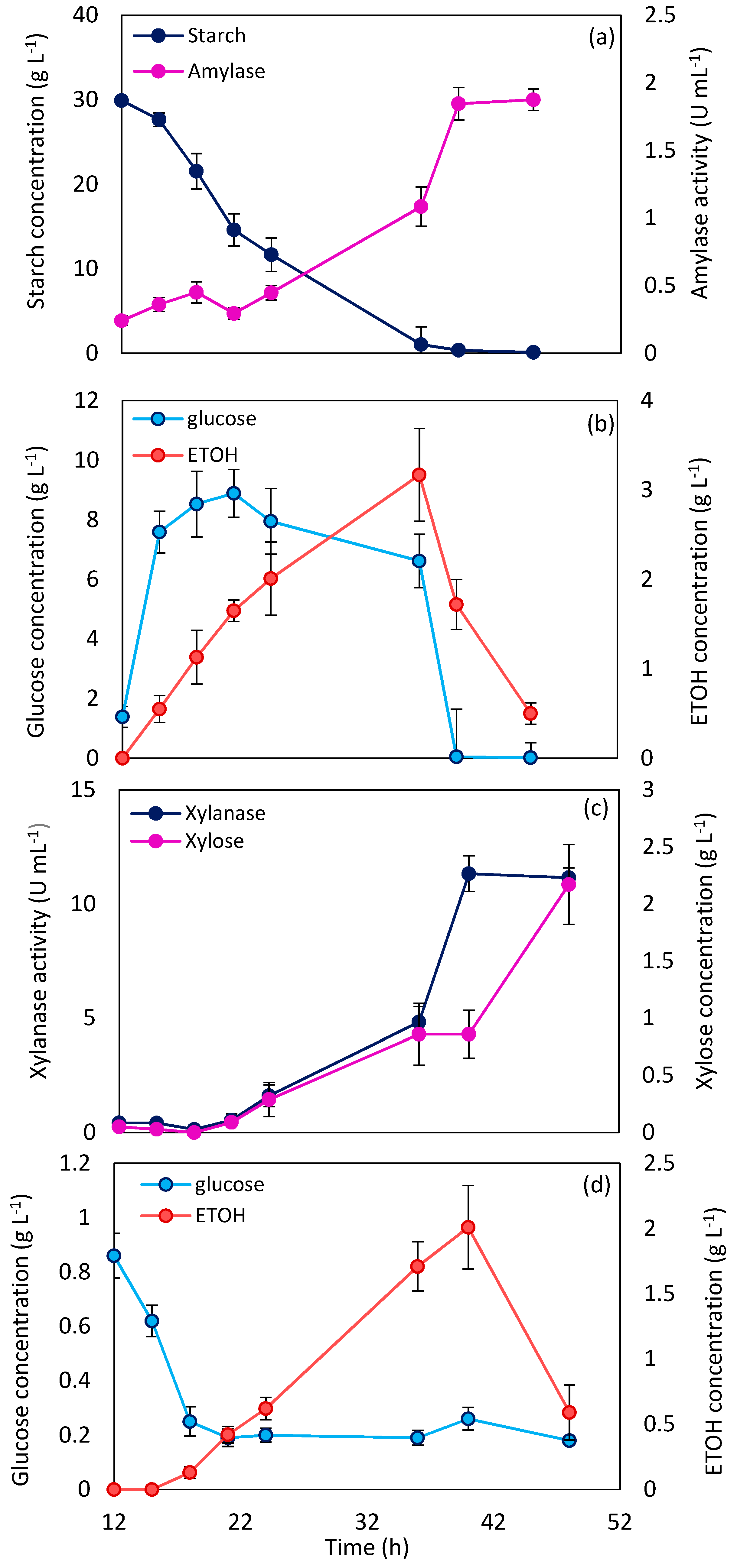
2.2. Enzyme Production in a Bubble Column Reactor
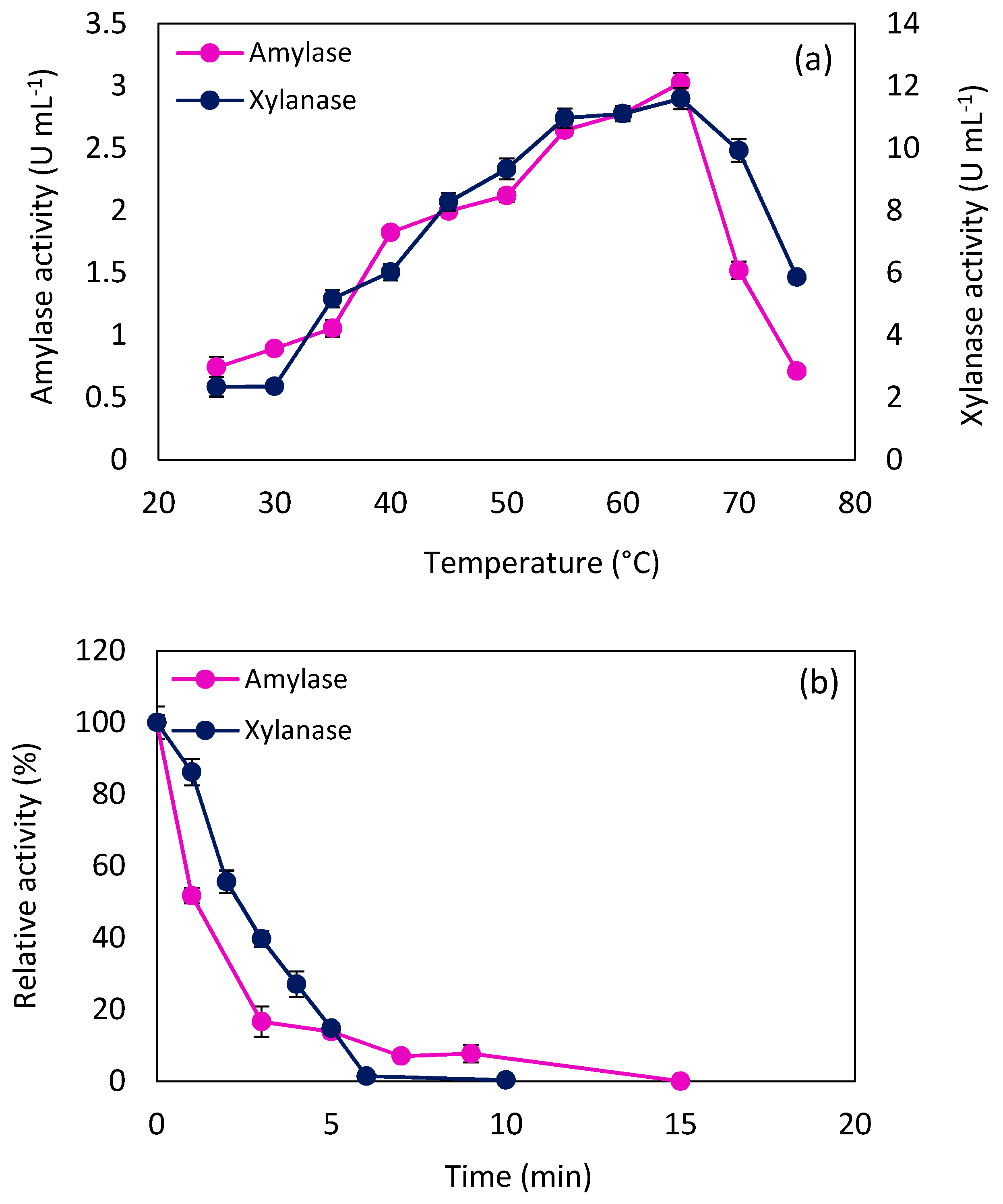
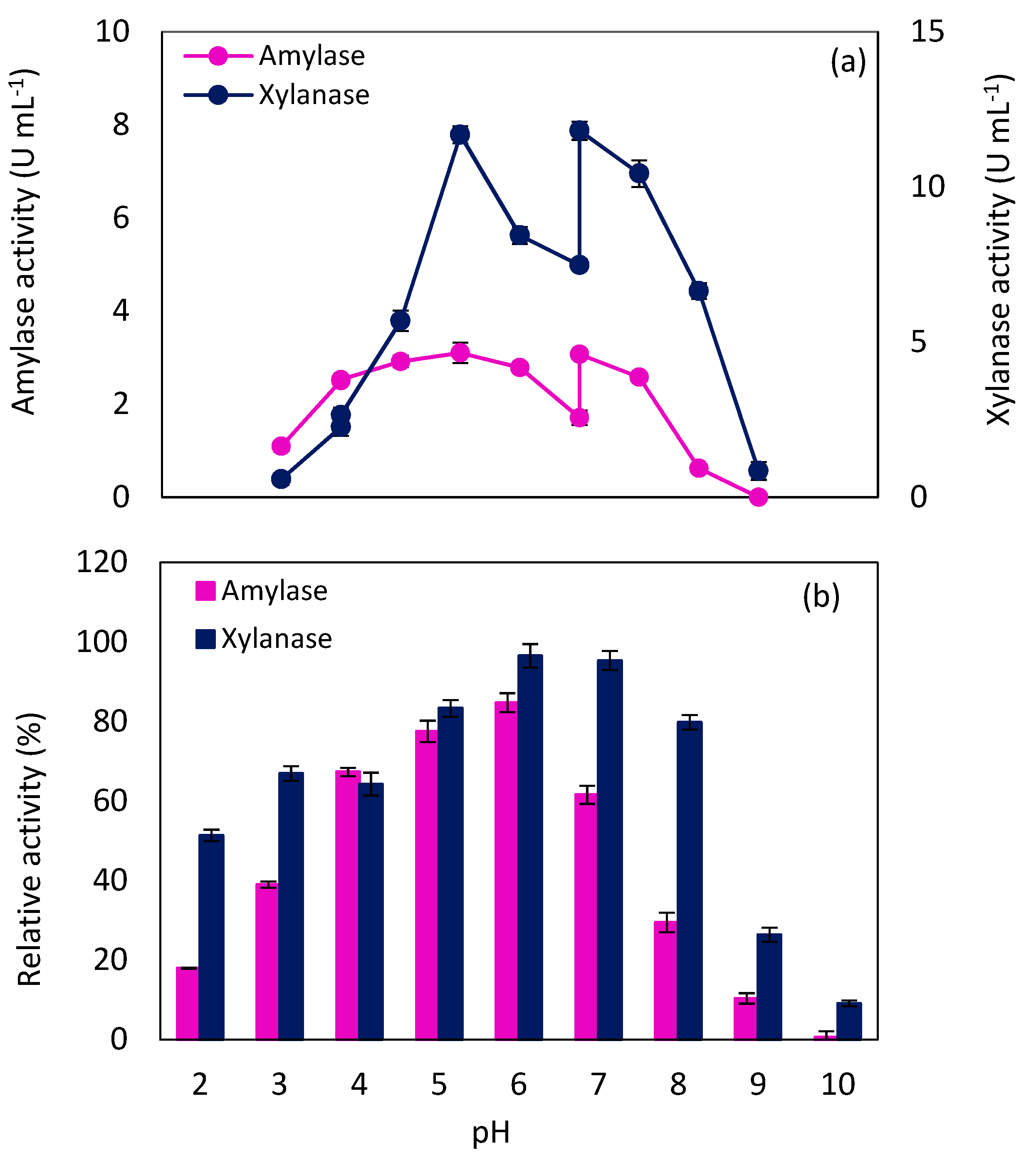
2.3. Amylase and Xylanase Characteristics
2.3.1. Effect of Temperature on Enzyme Activity
2.3.2. Effect of pH on Enzyme Activity
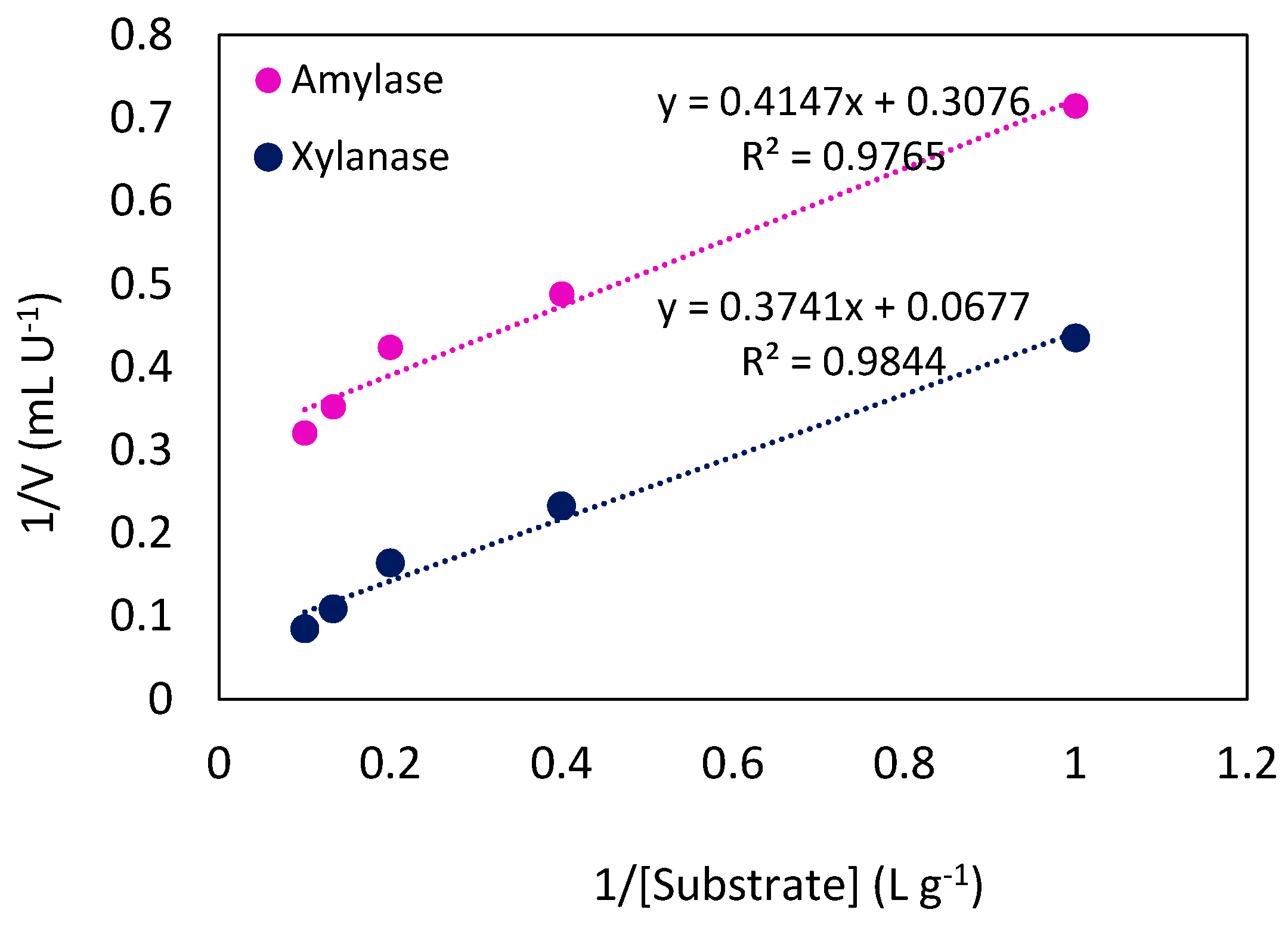
2.3.3. Kinetic Characteristics of Produced Enzymes
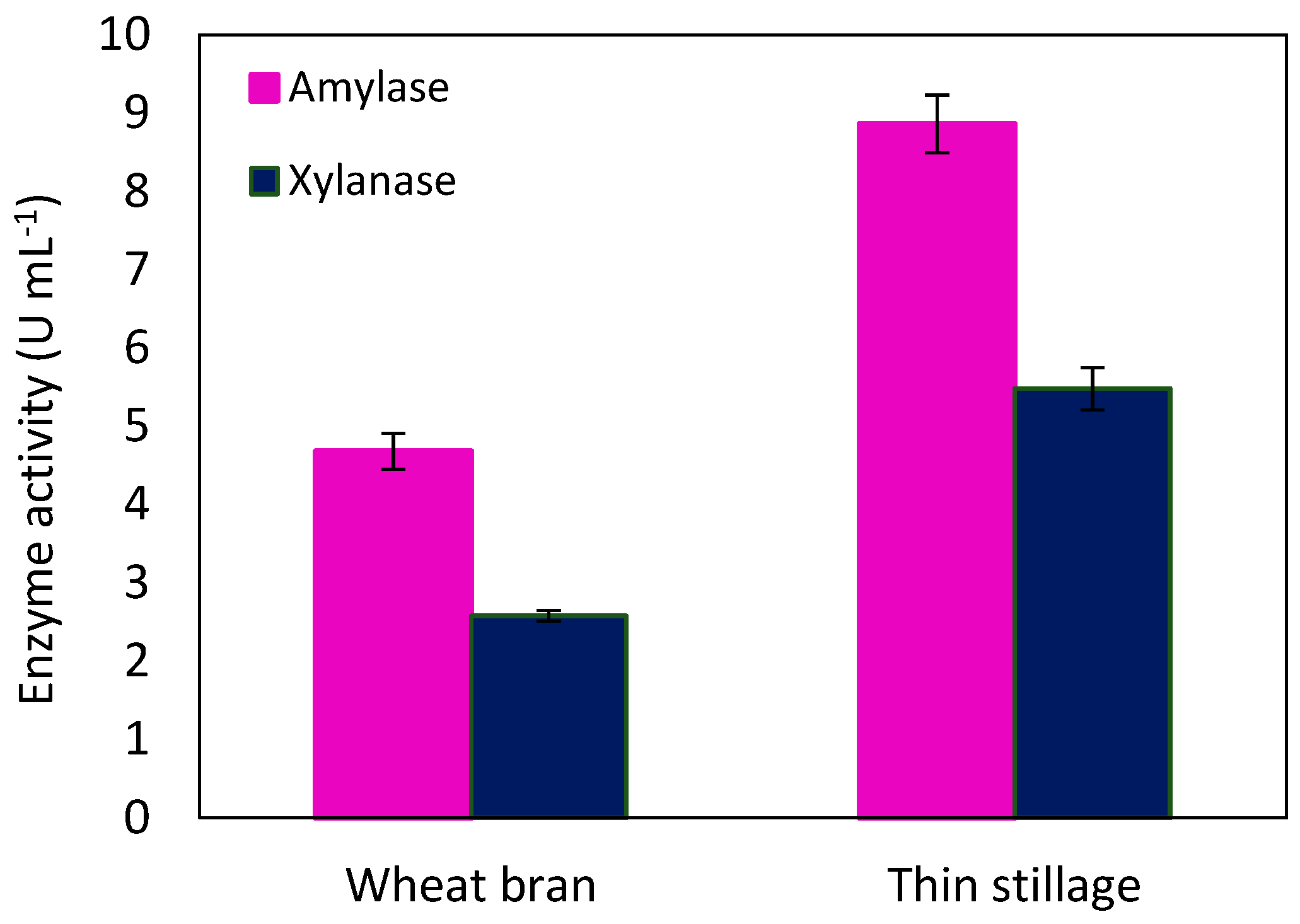
2.4. Enzyme Production by N. Intermedia through Wheat-Based Biorefinery
3. Materials and Methods
3.1. Microorganism
3.2. Substrate Media
3.3. Enzyme Production in Shake Flasks and Nitrogen Source
3.4. Enzyme Production in a Bubble Column Bioreactor
3.5. Enzyme Characteristics
3.5.1. Effect of pH and Temperature on Enzyme Activity
3.5.2. Kinetic Properties of the Enzyme-Substrate System
3.6. Analytical Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Souza, P.M. Application of microbial α-amylase in industry-A review. Braz. J. Microbiol. 2010, 41, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Purohit, A.; Rai, S.K.; Chownk, M.; Sangwan, R.S.; Yadav, S.K. Xylanase from Acinetobacter pittii MASK 25 and developed magnetic cross-linked xylanase aggregate produce predominantly xylopentose and xylohexose from agro biomass. Bioresour. Technol. 2017, 244, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, S.S.; Kumar, J.; Kumar, O.; Mishra, S.V.; Malyan, S.; Kumar, R. Xylanase and their industrial applications: A review. Biochem. Cell. Arch. 2017, 17, 353–360. [Google Scholar]
- Fath, M.; Fazaelipoor, M.H. Production of proteases in a novel trickling tray bioreactor. Waste Biomass Valoriz. 2015, 6, 475–480. [Google Scholar] [CrossRef]
- Beg, Q.; Kapoor, M.; Mahajan, L.; Hoondal, G. Microbial xylanases and their industrial applications: A review. Appl. Microbiol. Biotechnol. 2001, 56, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: A biotechnological perspective. Process Biochem. 2003, 38, 1599–1616. [Google Scholar] [CrossRef]
- Shahryari, Z.; Fazaelipoor, M.H.; Setoodeh, P.; Nair, R.B.; Taherzadeh, M.J.; Ghasemi, Y. Utilization of wheat straw for fungal phytase production. Int. J. Recycl. Org. Waste Agric. 2018, 7, 345–355. [Google Scholar] [CrossRef]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous ascomycetes fungi as a source of natural pigments. Fungal. Biol. Biotechnol. 2017, 4, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bátori, V.; Ferreira, J.A.; Taherzadeh, M.J.; Lennartsson, P.R. Ethanol and protein from ethanol plant by-products using edible fungi Neurospora intermedia and Aspergillus oryzae. BioMed Res. Int. 2015, 1–10. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Production of ethanol and biomass from thin stillage by neurospora intermedia: A pilot study for process diversification. Eng. Life Sci. 2015, 15, 751–759. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Production of ethanol and biomass from thin stillage using food-grade zygomycetes and ascomycetes filamentous fungi. Energies 2014, 7, 3872–3885. [Google Scholar] [CrossRef]
- Gmoser, R.; Ferreira, J.A.; Lundin, M.; Taherzadeh, M.J.; Lennartsson, P.R. Pigment production by the edible filamentous fungus Neurospora intermedia. Fermentation 2018, 4, 1–11. [Google Scholar]
- Nair, R.B.; Kabir, M.M.; Lennartsson, P.R.; Taherzadeh, M.J.; Horváth, I.S. Integrated process for ethanol, biogas, and edible filamentous fungi-based animal feed production from dilute phosphoric acid-pretreated wheat straw. Appl. Biochem. Biotechnol. 2018, 184, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.B.; Lundin, M.; Brandberg, T.; Lennartsson, P.R.; Taherzadeh, M.J. Dilute phosphoric acid pretreatment of wheat bran for enzymatic hydrolysis and subsequent ethanol production by edible fungi Neurospora intermedia. Ind Crops Prod. 2015, 69, 314–323. [Google Scholar] [CrossRef]
- Nair, R.B.; Lundin, M.; Lennartsson, P.R.; Taherzadeh, M.J. Optimizing dilute phosphoric acid pretreatment of wheat straw in the laboratory and in a demonstration plant for ethanol and edible fungal biomass production using Neurospora intermedia. J. Chem. Technol. Biotechnol. 2017, 92, 1256–1265. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Mahboubi, A.; Lennartsson, P.R.; Taherzadeh, M.J. Waste biorefineries using filamentous ascomycetes fungi: Present status and future prospects. Bioresour. Technol. 2016, 215, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.B.; Taherzadeh, M.J. Valorization of sugar-to-ethanol process waste vinasse: A novel biorefinery approach using edible ascomycetes filamentous fungi. Bioresour. Technol. 2016, 221, 469–476. [Google Scholar] [CrossRef]
- Mahboubi, A.; Ferreira, J.A.; Taherzadeh, M.J.; Lennartsson, P.R. Value-added products from dairy waste using edible fungi. Waste Manag. 2017, 59, 518–525. [Google Scholar] [CrossRef]
- Thunuguntla, R.; Mahboubi, A.; Ferreira, J.; Taherzadeh, M. Integration of membrane bioreactors with edible filamentous fungi for valorization of expired milk. Sustainability 2018, 10, 1940. [Google Scholar] [CrossRef]
- Adhyaru, D.N.; Bhatt, N.S.; Modi, H.A. Optimization of upstream and downstream process parameters for cellulase-poor-thermo-solvent-stable xylanase production and extraction by Aspergillus tubingensis fdhn1. Bioresour Bioprocess. 2015, 2, 1–14. [Google Scholar] [CrossRef]
- Van der Kaaij, R.; Janeček, Š.; van der Maarel, M.; Dijkhuizen, L. Phylogenetic and biochemical characterization of a novel cluster of intracellular fungal α-amylase enzymes. Microbiology 2007, 153, 4003–4015. [Google Scholar] [CrossRef] [PubMed]
- Mishra, C.; Keskar, S.; Rao, M. Production and properties of extracellular endoxylanase from Neurospora crassa. Appl. Environ. Microbiol. 1984, 48, 224–228. [Google Scholar] [PubMed]
- Kanti, A. Effect of nitrogen addition on the α-amylase production by Aspergillus niger, Rhizopus oligosporus and Neurospora crassa in media contained sargassum and rice seed on solid state fermentation. J. Biolog. Indones 2016, 12, 249–256. [Google Scholar]
- Pino, M.S.; Rodríguez-Jasso, R.M.; Michelin, M.; Flores-Gallegos, A.C.; Morales-Rodriguez, R.; Teixeira, J.A.; Ruiz, H.A. Bioreactor design for enzymatic hydrolysis of biomass under the biorefinery concept. Chem. Eng. J. 2018, 347, 119–136. [Google Scholar] [CrossRef]
- Gangadharan, D.; Nampoothiri, K.M.; Pandey, A. α-amylase production by Bacillus amyloliquefaciens using agro wastes as feed stock. Food Technol. Biotechnol. 2011, 49, 336–340. [Google Scholar]
- Melikoglu, M.; Lin, C.S.K.; Webb, C. Kinetic studies on the multi-enzyme solution produced via solid state fermentation of waste bread by Aspergillus awamori. Biochem. Eng. J. 2013, 80, 76–82. [Google Scholar] [CrossRef]
- Pasin, T.M.; Benassi, V.M.; Heinen, P.R.; de Lima Damasio, A.R.; Cereia, M.; Jorge, J.A.; de Moraes, M.d.L.T. Purification and functional properties of a novel glucoamylase activated by manganese and lead produced by Aspergillus japonicus. Int. J. Biol. Macromol. 2017, 102, 779–788. [Google Scholar] [CrossRef] [PubMed]
- de Lima Damásio, A.R.; Silva, T.M.; dos Reis Almeida, F.B.; Squina, F.M.; Ribeiro, D.A.; Leme, A.F.P.; Segato, F.; Prade, R.A.; Jorge, J.A.; Terenzi, H.F. Heterologous expression of an Aspergillus niveus xylanase gh11 in Aspergillus nidulans and its characterization and application. Process Biochem. 2011, 46, 1236–1242. [Google Scholar] [CrossRef]
- Shen, H.; Mo, X.; Chen, X.; Han, D.; Zhao, C. Purification and enzymatic identification of an acid stable and thermostable α-amylase from Rhizopus microsporus. J. Inst. Brew 2012, 118, 309–314. [Google Scholar] [CrossRef]
- Sugumaran, K.; Srivastava, S.; Ponnusami, V. Kinetic and thermodynamic characterization of Pleurotus eryngii mtcc 1798 xylanase and Fusarium oxysporum mtcc 3300 xylanase: A comparision. Int. J. Chem. Sci. 2012, 10, 1626–1636. [Google Scholar]
- Ward, O.; Moo-Young, M. Thermostable enzymes. Biotechnol. Adv. 1988, 6, 39–69. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Q.; Liao, S.; He, B.; Huang, R. Thermal stability and activity improvements of a ca-independent α-amylase from Bacillus subtilis cn7 by c-terminal truncation and hexahistidine-tag fusion. Biotechnol. Appl. Biochem. 2014, 61, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Negi, S.; Banerjee, R. Characterization of amylase and protease produced by Aspergillus awamori in a single bioreactor. Food Res. Int. 2009, 42, 443–448. [Google Scholar] [CrossRef]
- Terrasan, C.R.F.; Guisan, J.M.; Carmona, E.C. Xylanase and β-xylosidase from Penicillium janczewskii: Purification, characterization and hydrolysis of substrates. Electron. J. Biotechnol. 2016, 23, 54–62. [Google Scholar] [CrossRef]
- Irfan, M.; Gonzalez, C.F.; Raza, S.; Rafiq, M.; Hasan, F.; Khan, S.; Shah, A.A. Improvement in thermostability of xylanase from Geobacillus thermodenitrificans c5 by site directed mutagenesis. Enzyme Microb. Technol. 2018, 111, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Kayastha, A.M. An antioxidant rich novel β-amylase from peanuts (arachis hypogaea): Its purification, biochemical characterization and potential applications. Int. J. Biol. Macromol. 2018, 111, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Li, M.; Hu, Y. High-activity production of xylanase by Pichia stipitis: Purification, characterization, kinetic evaluation and xylooligosaccharides production. Int. J. Biol. Macromol. 2018, 117, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Le Mense, E.; Corman, J.; Van Lanen, J.; Langlykke, A. Production of mold amylases in submerged culture. J. Bacteriol. 1947, 54, 149–159. [Google Scholar]
- Koutinas, A.; Arifeen, N.; Wang, R.; Webb, C. Cereal-based biorefinery development: Integrated enzyme production for cereal flour hydrolysis. Biotechnol. Bioeng. 2007, 97, 61–72. [Google Scholar] [CrossRef]
- Nair, R.B.; Kalif, M.; Ferreira, J.A.; Taherzadeh, M.J.; Lennartsson, P.R. Mild-temperature dilute acid pretreatment for integration of first and second generation ethanol processes. Bioresour. Technol. 2017, 245, 145–151. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Yeng, A.L.Y.; Ab Kadir, M.S.; Ghazali, H.M.; Rahman, R.N.Z.R.A.; Saari, N. A comparative study of extraction techniques for maximum recovery of glutamate decarboxylase (GAD) from Aspergillus oryzae nsk. BMC Res. Notes 2013, 6, 1–9. [Google Scholar]
- Xiao, Z.; Storms, R.; Tsang, A. A quantitative starch-iodine method for measuring alpha-amylase and glucoamylase activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature Range (°C) | Ea, app (kJ mol−1) | |
|---|---|---|
| Amylase | Xylanase | |
| 25–65 | 30.97 | 36.65 |
| 65–75 | −141.570 | −66.52 |
| Enzyme | kd (min−1) | (min) |
|---|---|---|
| Amylase | 0.2862 | 2.42 |
| Xylanase | 0.3390 | 2.04 |
| Enzyme | Km, app (g L−1) | Vmax, app (U mL−1) |
|---|---|---|
| Amylase | 1.35 | 3.25 |
| Xylanase | 5.52 | 14.77 |
| Component | g g−1 Wheat Bran (dry basis) [40] | g L−1 Thin Stillage [10] |
|---|---|---|
| Solid fraction | ||
| Arabinan | 0.088 ± 0.001 | 0.242 ± 0.140 |
| Crude protein | 0.147 ± 0.001 | 27.5 ± 1.25 |
| Galactan | <0.001 | 0.175 ± 0.0420 |
| Glucan | 0.204 ± 0.02 | 3.182 ± 0.383 |
| Mannan | <0.001 | 0.488 ± 0.234 |
| Starch | 0.17 ± 0.0016 | <0.001 |
| Xylan | 0.156 ± 0.019 | 0.907 ± 0.419 |
| Dissolved saccharides | ||
| Arabinos | 4.4 ± 0.2 | |
| Galactose | 1.6 ± 0.2 | |
| Glucose | 9.8 ± 0.7 | |
| Mannose | 1.4 ± 0.2 | |
| Xylose | 6.1 ± 0.4 | |
| Starch (using glucose as an indicator) | 8.82 ± 0.63 | |
| Xylan (using xylose as an indicator) | 5.36 ± 0.35 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahryari, Z.; Fazaelipoor, M.H.; Ghasemi, Y.; Lennartsson, P.R.; Taherzadeh, M.J. Amylase and Xylanase from Edible Fungus Neurospora intermedia: Production and Characterization. Molecules 2019, 24, 721. https://doi.org/10.3390/molecules24040721
Shahryari Z, Fazaelipoor MH, Ghasemi Y, Lennartsson PR, Taherzadeh MJ. Amylase and Xylanase from Edible Fungus Neurospora intermedia: Production and Characterization. Molecules. 2019; 24(4):721. https://doi.org/10.3390/molecules24040721
Chicago/Turabian StyleShahryari, Zohre, Mohammad H. Fazaelipoor, Younes Ghasemi, Patrik R. Lennartsson, and Mohammad J. Taherzadeh. 2019. "Amylase and Xylanase from Edible Fungus Neurospora intermedia: Production and Characterization" Molecules 24, no. 4: 721. https://doi.org/10.3390/molecules24040721
APA StyleShahryari, Z., Fazaelipoor, M. H., Ghasemi, Y., Lennartsson, P. R., & Taherzadeh, M. J. (2019). Amylase and Xylanase from Edible Fungus Neurospora intermedia: Production and Characterization. Molecules, 24(4), 721. https://doi.org/10.3390/molecules24040721