Inhibitory Effects of Roseoside and Icariside E4 Isolated from a Natural Product Mixture (No-ap) on the Expression of Angiotensin II Receptor 1 and Oxidative Stress in Angiotensin II-Stimulated H9C2 Cells

 and
and 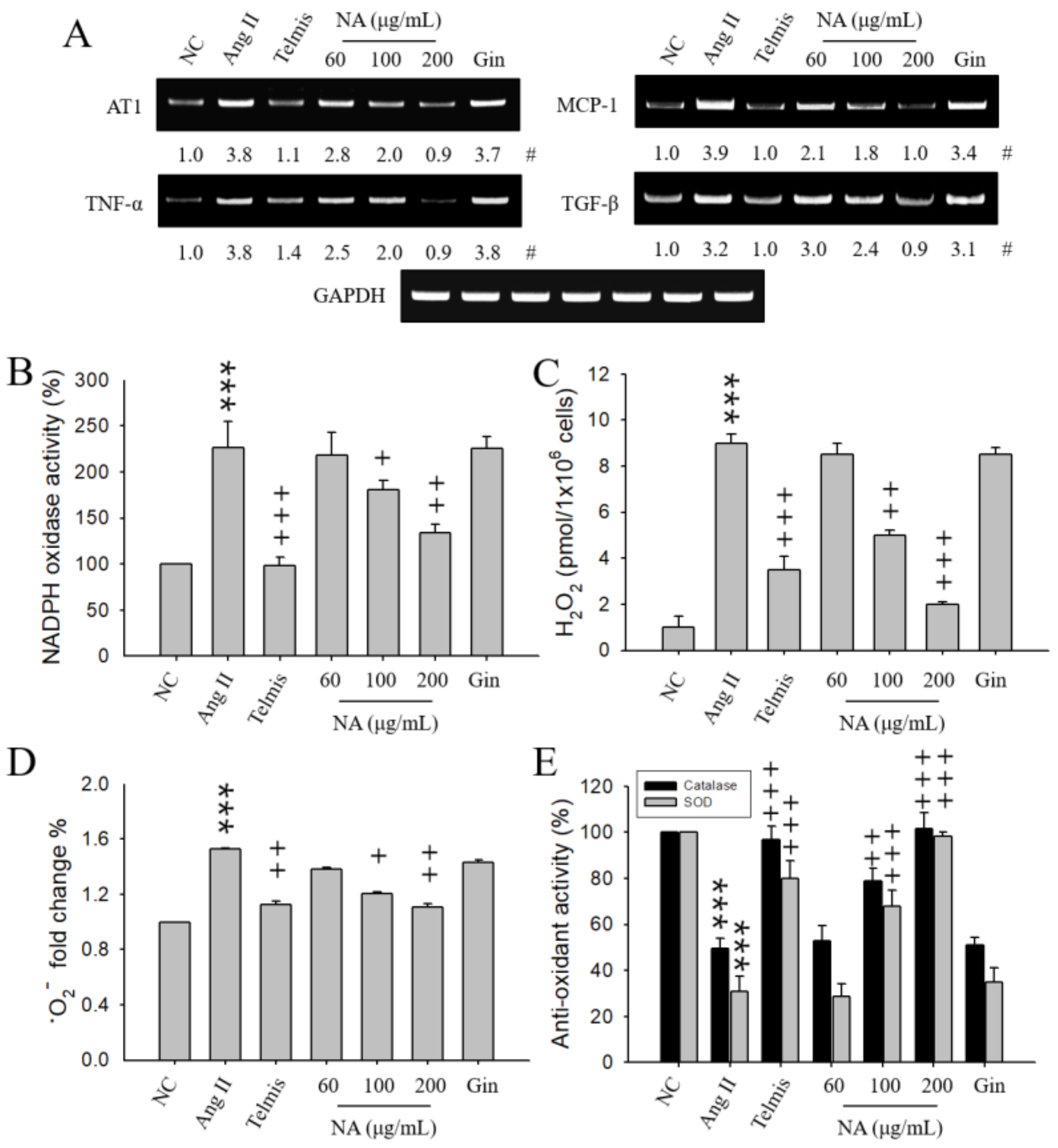{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Effects of NA on the Expression of Hypertension-Related Molecules in Ang II-Stimulated H9C2 Cells
2.2. Effects of NA on Oxidative Stress in Ang II-Stimulated H9C2 Cells
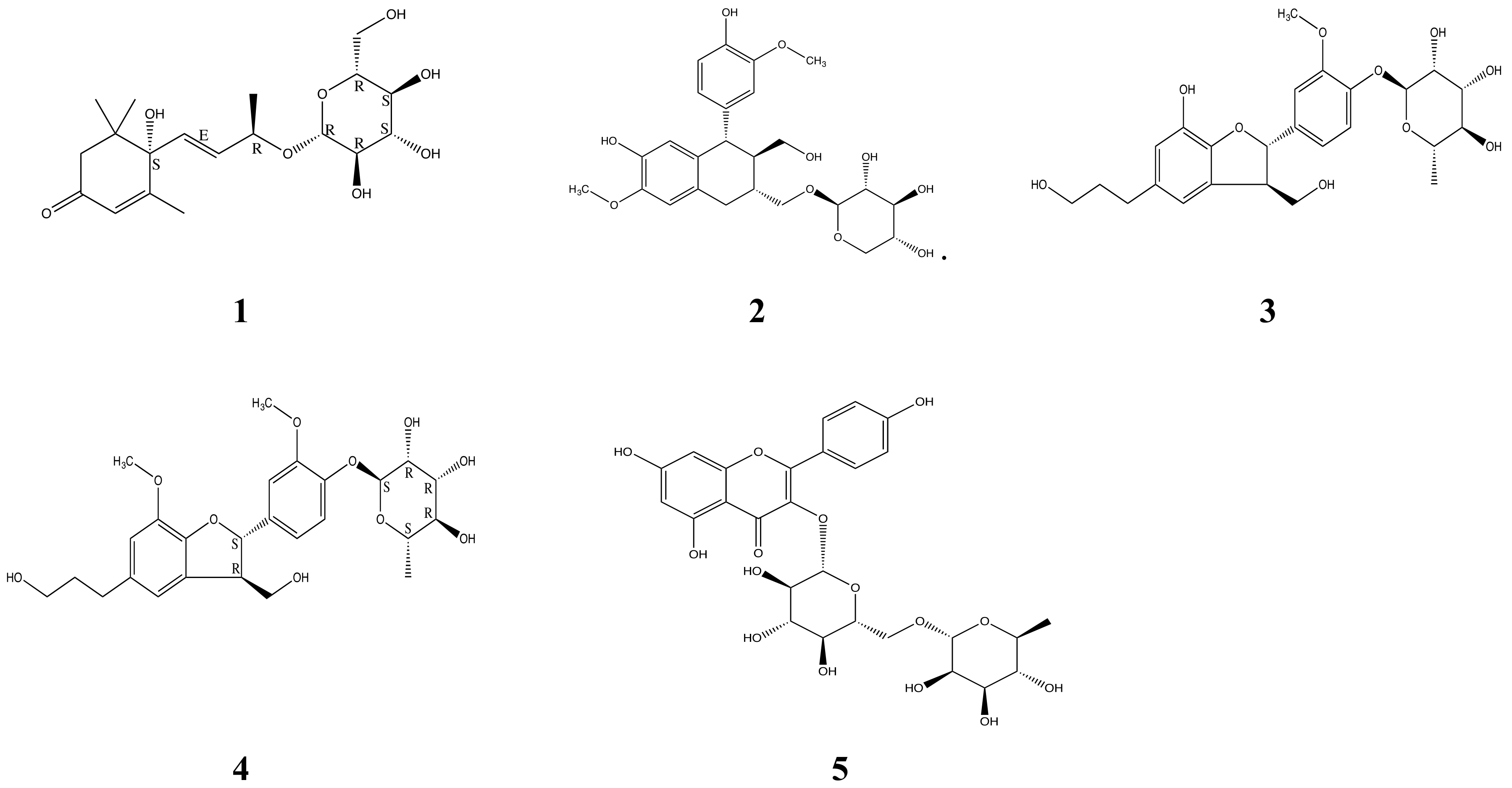
2.3. Purification and Identification of Bioactive Ingredients in NA
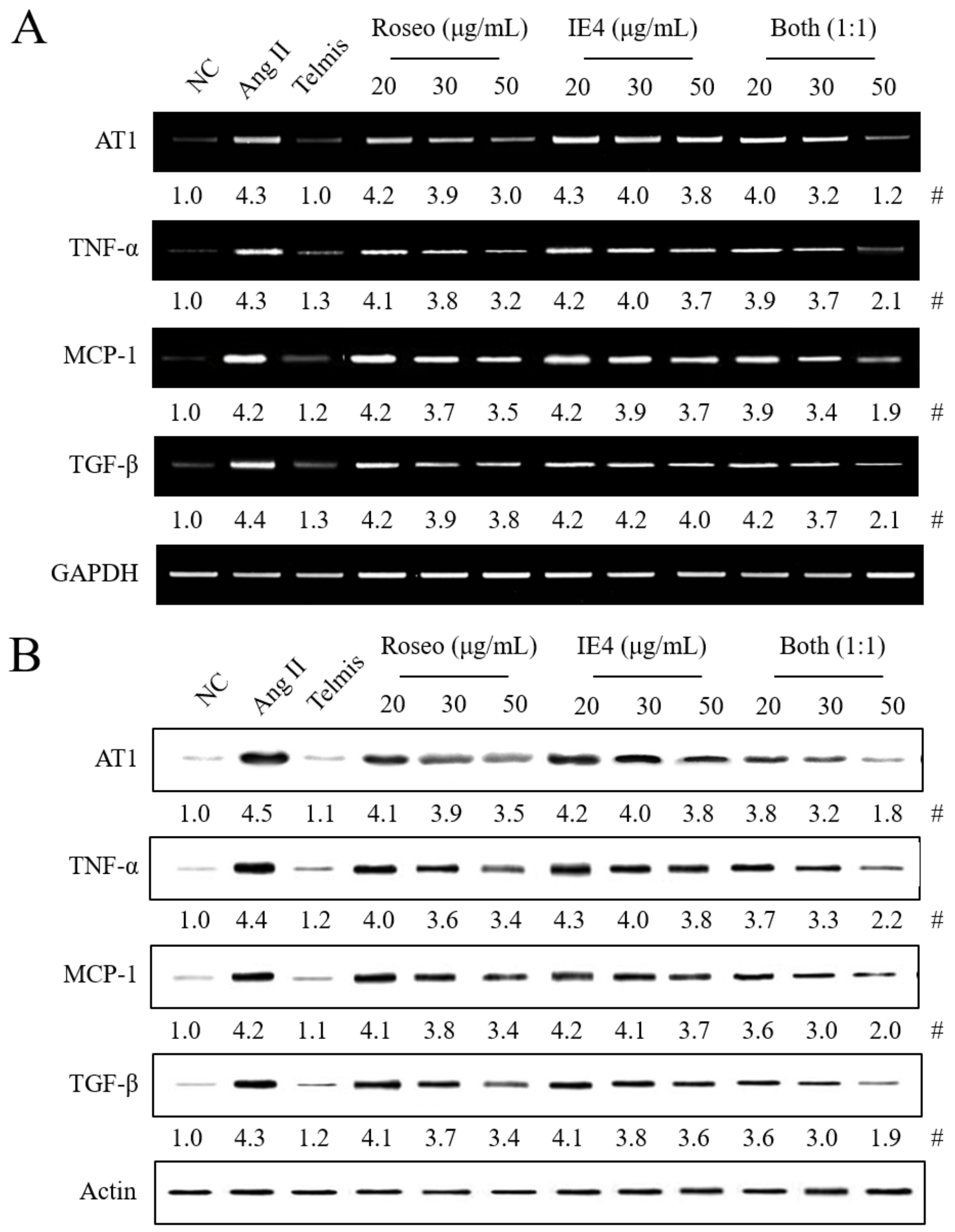
2.4. Effects of the Isolated Components Roseoside and Icariside E4 on the Expression of Hypertension-Related Molecules in Ang II-Stimulated H9C2 Cells
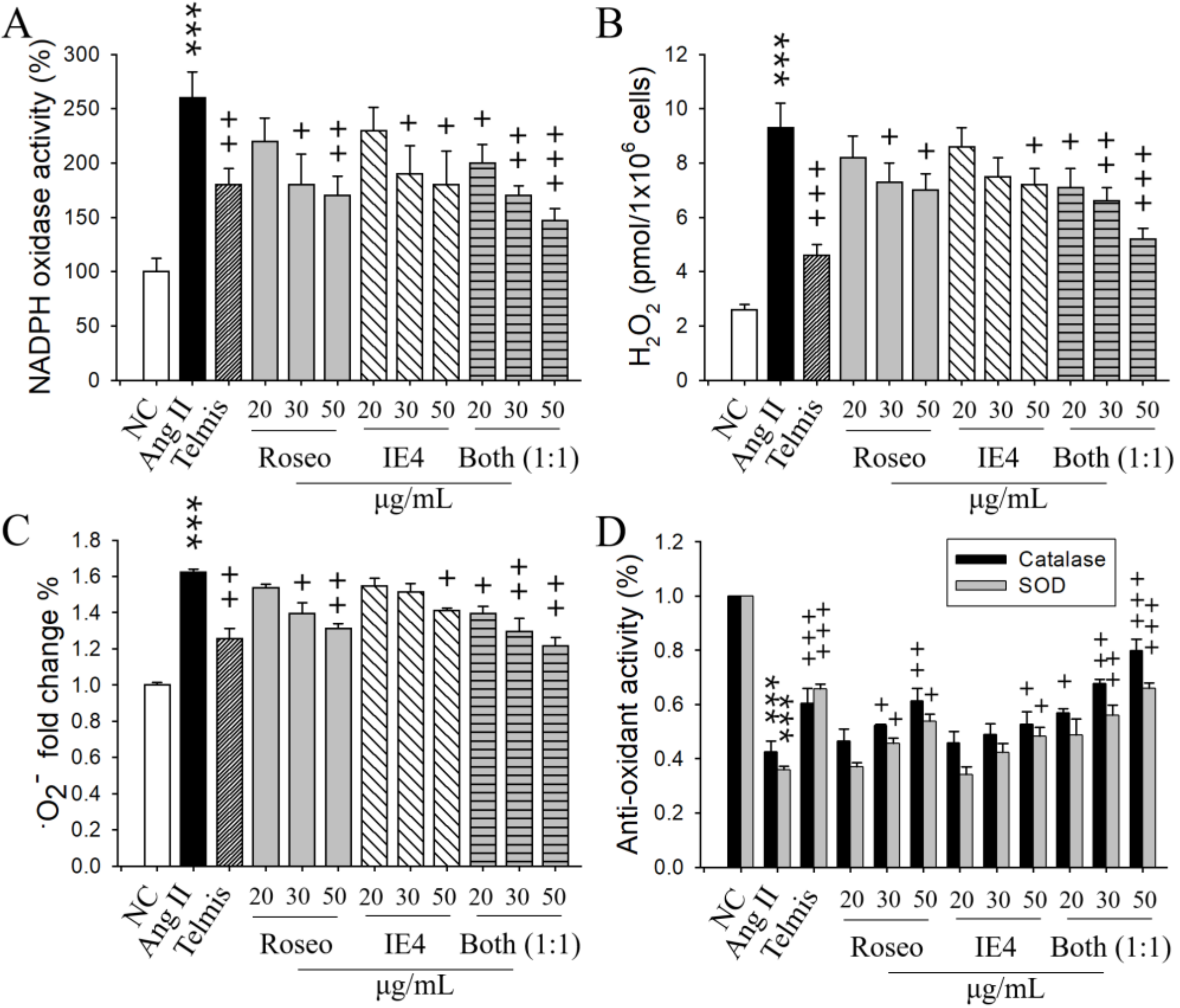
2.5. Respective Effects of Roseoside and Icariside E4 on Oxidative stress in Ang II-Stimulated H9C2 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Purification and Identification of Bioactive Ingredients
4.3. Cardiomyocytic H9C2 Cell Line Culture
4.4. Cell line Stimulation and Treatment
4.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.6. Determination of the Activity of NADPH Oxidase, Superoxide Dismutase, and Catalase and the Levels of Hydrogen Peroxide and Superoxide Anion
4.7. Western Blot Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lackland, D.T.; Weber, M.A. Global burden of cardiovascular disease and stroke: Hypertension at the core. Can. J. Cardiol. 2015, 31, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Dinh Cat, A.; Montezano, A.C.; Burger, D.; Touyz, R.M. Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid. Redox Signal. 2013, 19, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Zablocki, D.; Sadoshima, J. Angiotensin II and oxidative stress in the failing heart. Antioxid. Redox Signal. 2013, 19, 1095–1109. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Takemoto, M.; Liao, J.K. NADPH oxidase-derived superoxide anion mediates angiotensin II-induced cardiac hypertrophy. J. Mol. Cell. Cardiol. 2003, 35, 851–859. [Google Scholar] [CrossRef]
- Takimoto, E.; Kass, D.A. Role of oxidative stress in cardiac hypertrophy and remodeling. Hypertension 2007, 49, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Liu, C.; Xue, R.; Wang, Y.; Sun, Y.; Liang, Z.; Fan, W.; Jiang, J.; Zhao, J.; Su, Q.; et al. Fisetin inhibits cardiac hypertrophy by suppressing oxidative stress. J. Nutr. Biochem. 2018, 62, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Oparil, S.; Williams, D.; Chrysant, S.G.; Marbury, T.C.; Neutel, J. Comparative efficacy of olmesartan, losartan, valsartan, and irbesartan in the control of essential hypertension. J. Clin. Hypertens. 2001, 3, 283–291. [Google Scholar] [CrossRef]
- Burnier, M.; Brunner, H.R. Angiotensin II receptor antagonists. Lancet 2000, 355, 637–645. [Google Scholar] [CrossRef]
- Shang, P.; Liu, T.; Liu, W.; Li, Y.; Dou, F.; Zhang, Y.; Sun, L.; Zhang, T.; Zhu, Z.; Mu, F.; et al. Telmisartan improves vascular remodeling through ameliorating prooxidant and profibrotic mechanisms in hypertension via the involvement of transforming growth factor-β1. Mol. Med. Reports. 2017, 16, 4537–4544. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, J.; Xu, Z.; Qiao, X.; Huang, L. Disappearance of polycyclic aromatic hydrocarbons sorbed on surfaces of pine [Pinua thunbergii] needles under irradiation of sunlight: Volatilization and photolysis. Atmos. Environ. 2005, 39, 4583–4591. [Google Scholar] [CrossRef]
- Yu, L.; Zhao, M.; Wang, J.S.; Cui, C.; Yang, B.; Jiang, Y.; Zhao, Q. Antioxidant, immunomodulatory and anti-breast cancer activities of phenolic extract from pine (Pinus massoniana Lamb) bark. Innov. Food Sic. Emerg. Technol. 2009, 9, 122–128. [Google Scholar] [CrossRef]
- Kim, J.-S. Evaluation of in vitro antioxidant activity of the water extract obtained from dried pine needle (Pinus densiflora). Prev. Nutr. Food Sci. 2018, 23, 134–143. [Google Scholar] [CrossRef]
- Venkatesan, T.; Choi, Y.-W.; Lee, J.; Kim, Y.-K. Pinus densiflora needle supercritical fluid extract suppresses the expression of pro-inflammatory mediators iNOS, IL-6 and IL-1β, and activation of inflammatory STAT1 and STAT3 signaling proteins in bacterial lipopolysaccharide-challenged murine macrophages. Daru. J. Pharmaceut. Sci. 2017, 25, 18–27. [Google Scholar]
- Cheong, H.S.; Lim, D.Y. Pine needle extracts inhibit contractile responses of the isolated rat aortic strips. Nat. Prod. Sci. 2010, 16, 123–132. [Google Scholar]
- Moghadamtousi, S.Z.; Fadaeinasab, M.; Nikzad, S.; Mohan, G.; Ali, H.M.; Kadir, H.A. Annona muricata (Annonaceae): A review of its traditional uses, isolated acetogenin and biological activities. Int. J. Mol. Sci. 2015, 16, 15625–15658. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Oyeleye, S.I.; Oboh, G. Distribution of phenolic contents, antidiabetic potentials, antihypertensive properties, and antioxidative effects of Soursop (Annona muricata L.) fruit parts in vitro. Biochem. Res. Int. 2015. [Google Scholar] [CrossRef]
- Ojewole, J.A.; Adewole, S.O.; Olayiwola, G. Hypoglycaemic and hypotensive effects of Momordica charantia Linn (Cucurbitaceae) whole-plant aqueous extract in rats. Cardiovasc. J. S. Afr. 2006, 17, 227–232. [Google Scholar]
- Shodehinde, S.A.; Adefegha, S.A.; Oboh, G.; Oyeleye, S.I.; Olasehinde, T.A.; Nwanna, E.E.; Adedayo, B.C.; Boligon, A.A. Phenolic composition and evaluation of methanol and aqueous extracts of bitter gourd (Momordica charantia L.) leaves on angiotensin-I-converting enzyme and some pro-oxidant-induced lipid peroxidation in vitro. J. Evid. Based Complementary Altern. Med. 2016, 21, 67–76. [Google Scholar] [CrossRef]
- Savoia, C.; Schiffrin, E.L. Inflammation in hypertension. Curr. Opin. Nephrol. Hypertens. 2006, 15, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Dinh, Q.N.; Drummond, G.R.; Sobey, C.G.; Chrissobolis, S. Roles of inflammation, oxidative stress, and vascular dysfunction in hypertension. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef]
- Agita, A.; Alsagaff, M.T. Inflammation, Immunity, and Hypertension. Acta. Med. Indones. 2017, 49, 158–165. [Google Scholar] [PubMed]
- Prathapan, A.; Vineetha, V.P.; Abhilash, P.A.; Raghu, K.G. Boerhaavia diffusa L. attenuates angiotensin II-induced hypertrophy in H9C2 cardiac myoblast cells via modulating oxidative stress and down-regulating NF-κβ and transforming growth factor β1. Br. J. Nutr. 2013, 110, 1201–1210. [Google Scholar] [CrossRef]
- Gray, S.P.; Jandeleit-Dahm, K.A. The role of NADPH oxidase in vascular disease--hypertension, atherosclerosis & stroke. Curr. Pharm. Des. 2015, 21, 5933–5944. [Google Scholar] [PubMed]
- Kuang, H.-X.; Yang, B.-Y.; Xia, Y.-G.; Feng, W.-S. Chemical constituents from the flower of Datura metel L. Arch. Pharm. Res. 2008, 31, 1094–1097. [Google Scholar] [CrossRef]
- Feng, W.-S.; Bi, Y.-F.; Zheng, X.-K.; Wang, S.-L.; Li, J. Studies on the lignin chemical constituents from pine needles of Pinus massomiana Lamb. Acta Pharmaceutica Sinica. 2003, 38, 199–202. [Google Scholar] [PubMed]
- Nakanishi, T.; Iida, N.; Inatomi, Y.; Murata, H.; Inada, A.; Murata, J.; Lang, F.A.; Iinuma, M.; Tanaka, M. Neolignan and flavonoid glycosides in Juniperus communis var. depressa. Phytochemistry 2004, 65, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Kim, J.H.; Choi, S.E.; Park, K.H.; Lee, M.W. Inhibitory effects of phenolic compounds from needles of Pinus densiflora on nitric oxide and PGE2 production. Arch. Pharm. Res. 2010, 33, 2011–2016. [Google Scholar] [CrossRef]
- Hou, W.-C.; Lin, R.-D.; Lee, T.-H.; Huang, Y.-H.; Hsu, F.-L.; Lee, M.-H. The phenolic constituents and free radical scavenging activities of Gynura formosana Kiamnra. J. Sci. Food Agri. 2005, 85, 615–621. [Google Scholar] [CrossRef]
- Lee, T.H.; Wang, G.J.; Lee, C.K.; Kuo, Y.H.; Chou, C.H. Inhibitory effects of glycosides from the leaves of Melaleuca quinquenervia on vascular contraction of rats. Planta Med. 2002, 68, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Frankish, N.; de Sousa Menezes, F.; Mills, C.; Sheridan, H. Enhancement of insulin release from the beta-cell line INS-1 by an ethanolic extract of Bauhinia variegata and its major constituent roseoside. Planta Med. 2010, 76, 995–997. [Google Scholar] [CrossRef]
- Bermudez, J.; Rodriguez, M.; Hasegawa, M.; Gonzalez-Mujica, F.; Duque, S.; Ito, Y. (6R,9S)-6′′-(4′′-hydroxybenzoyl)-roseoside, a new megastigmane derivative from Ouratea polyantha and its effect on hepatic glucose-6-phosphatase. Nat. Prod. Commun. 2012, 7, 973–976. [Google Scholar] [PubMed]
- Simaratanamongkol, A.; Umehara, K.; Noguchi, H.; Panichayupakaranant, P. Identification of a new angiotensin-converting enzyme (ACE) inhibitor from Thai edible plants. Food Chem. 2014, 165, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Huong, N.T.; Vien, L.T.; Hanh, T.T.; Dang, N.H.; Thanh, N.V.; Cuong, N.X.; Nam, N.H.; Truong, L.H.; Ban, N.K.; Kiem, P.V.; et al. Triterpene saponins and megastigmane glucosides from Camellia bugiamapensis. Bioorg. Med. Chem. Lett. 2017, 27, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.; Spinola, V.; Llorent-Martinerz, E.J.; Fernandez-de Cordova, M.L.; Molina-Garcia, L.; Castilho, P.C. Polyphenolic profile and antioxidant activities of Madeiran elderberry (Sambucus lanceolata) as affected by simulated in vitro digestion. Food Res. Int. 2017, 100, 404–410. [Google Scholar] [CrossRef]
- Lee, Y.R.; Park, J.H.; Castaneda Molina, R.; Nam, Y.H.; Lee, Y.G.; Hong, B.N.; Baek, N.I.; Kang, T.H. Skin depigmenting action of silkworm (Bombyx mori L.) droppings in zebrafish. Arch. Dermatol. Res. 2018, 310, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Joo, T.; Sowndhararajan, K.; Hong, S.; Lee, J.; Park, S.Y.; Kim, S.; Jhoo, J.W. Inhibition of nitric oxide production in LPS-stimulated RAW 264.7 cells by stem bark of Ulmus pumila L. Saudi J. Biol. Sci. 2014, 21, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Mina, S.A.; Melek, F.R.; Adeeb, R.M.; Hagag, E.G. LC/ESI-MS/MS profiling of Ulmus parvifolia extracts and evaluation of its anti-inflammatory, cytotoxic, and antioxidant activities. Z. Naturforsch. C. 2016, 71, 415–421. [Google Scholar] [CrossRef]
- Ferreira-Junior, J.C.; Conserva, L.M.; Lyra Lemos, R.P.; de Omena-Neta, G.C.; Cavalcante-Neto, A.; Barreto, E. Isolation of a dihydrobenzofuran lignan, icariside E4, with an antinociceptive effect from Tabebuia roseo-alba (Ridley) Sandwith (Bignoniaceae) bark. Arch. Pharm. Res. 2015, 38, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Dubois, R.; Young, C.; Lien, E.J.; Adams, J.D. Heteromeles arbutifolia, a traditional treatment for alzheimer’s disease, phytochemistry and safety. Medicines 2016, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.J.; Kang, B.W.; Jeong, Y.K. Identification of a neolignan glycoside from the pine tree, Pinus densiflora showed antithrombotic activity. J. Life Sci. 2014, 24, 873–879. [Google Scholar] [CrossRef]
- Hong, S.S.; Jeong, W.; Kim, J.K.; Kwon, J.G.; Lee, J.Y.; Ahn, E.K.; Oh, J.; Seo, D.W.; Oh, J.S. Neolignan inhibitors of antigen-induced degranulation in RBL-2H3 cells from the needles of Pinus thunbergii. Fitoterapia 2014, 99, 347–351. [Google Scholar] [CrossRef]
- Chen, G.-H.; Li, Y.-C.; Lin, N.-H.; Kuo, P.-C.; Tzen, J.T.C. Characterization of vasorelaxant principles from the needles of Pinus morrisonicola Hayata. Molecules 2018, 23, 86. [Google Scholar] [CrossRef] [PubMed]
- Montezano, A.C.; Nguyen Dinh Cat, A.; Rios, F.J.; Touyz, R.M. Angiotensin II and vascular injury. Curr. Hypertens. Rep. 2014, 16, 431. [Google Scholar] [CrossRef]
- Lee, H.; Kim, K.C.; Hong, Y.M. Changes of bax, Bcl2, chemokine receptor-2, monocyte chemoattractant protein-1, and transforming growth factor β1 genes in the left ventricle of spontaneously hypertensive rat after losartan treatment. Kor. J. Pediat. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lijnen, P.J.; van Pelt, J.F.; Fagard, R.H. Downregulation of manganese superoxide dismutase by angiotensin II in cardiac fibroblasts of rats: Association with oxidative stress in myocardium. Am. J. Hypertens. 2010, 23, 1128–1135. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhang, J.D.; Wang, B.; Lv, Y.J.; Jiang, H.; Liu, G.L.; Qiao, Y.; Ren, M.; Guo, X.F. Quercetin inhibits left ventricular hypertrophy in spontaneously hypertensive rats and inhibits angiotensin II-induced H9C2 cells hypertrophy by enhancing PPAR-gamma expression and suppressing AP-1 activity. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.U.; Kim, N.G.; Kim, T.J.; Ro, J.Y. CD1d expressed in mast cell surface enhances IgE production in B cells by up-regulating CD40L expression and mediator release in allergic asthma in mice. Cell. Signal. 2014, 26, 1105–1117. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the corresponding authors (Y.-I.K.; J.Y.R.). |




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, E.Y.; Kim, T.Y.; Hong, G.U.; Kang, H.; Lee, J.-Y.; Park, J.Y.; Kim, S.-C.; Kim, Y.H.; Chung, M.-H.; Kwon, Y.-I.; et al. Inhibitory Effects of Roseoside and Icariside E4 Isolated from a Natural Product Mixture (No-ap) on the Expression of Angiotensin II Receptor 1 and Oxidative Stress in Angiotensin II-Stimulated H9C2 Cells. Molecules 2019, 24, 414. https://doi.org/10.3390/molecules24030414
Hong EY, Kim TY, Hong GU, Kang H, Lee J-Y, Park JY, Kim S-C, Kim YH, Chung M-H, Kwon Y-I, et al. Inhibitory Effects of Roseoside and Icariside E4 Isolated from a Natural Product Mixture (No-ap) on the Expression of Angiotensin II Receptor 1 and Oxidative Stress in Angiotensin II-Stimulated H9C2 Cells. Molecules. 2019; 24(3):414. https://doi.org/10.3390/molecules24030414
Chicago/Turabian StyleHong, Eun Young, Tae Yang Kim, Gwan Ui Hong, Hanna Kang, Jung-Yun Lee, Jae Yeo Park, Se-Chan Kim, Young Ho Kim, Myung-Hee Chung, Young-In Kwon, and et al. 2019. "Inhibitory Effects of Roseoside and Icariside E4 Isolated from a Natural Product Mixture (No-ap) on the Expression of Angiotensin II Receptor 1 and Oxidative Stress in Angiotensin II-Stimulated H9C2 Cells" Molecules 24, no. 3: 414. https://doi.org/10.3390/molecules24030414
APA StyleHong, E. Y., Kim, T. Y., Hong, G. U., Kang, H., Lee, J.-Y., Park, J. Y., Kim, S.-C., Kim, Y. H., Chung, M.-H., Kwon, Y.-I., & Ro, J. Y. (2019). Inhibitory Effects of Roseoside and Icariside E4 Isolated from a Natural Product Mixture (No-ap) on the Expression of Angiotensin II Receptor 1 and Oxidative Stress in Angiotensin II-Stimulated H9C2 Cells. Molecules, 24(3), 414. https://doi.org/10.3390/molecules24030414