Simultaneous Use of Stimulatory Agents to Enhance the Production and Hypoglycaemic Activity of Polysaccharides from Inonotus obliquus by Submerged Fermentation

 and
and
Abstract
:1. Introduction
2. Results
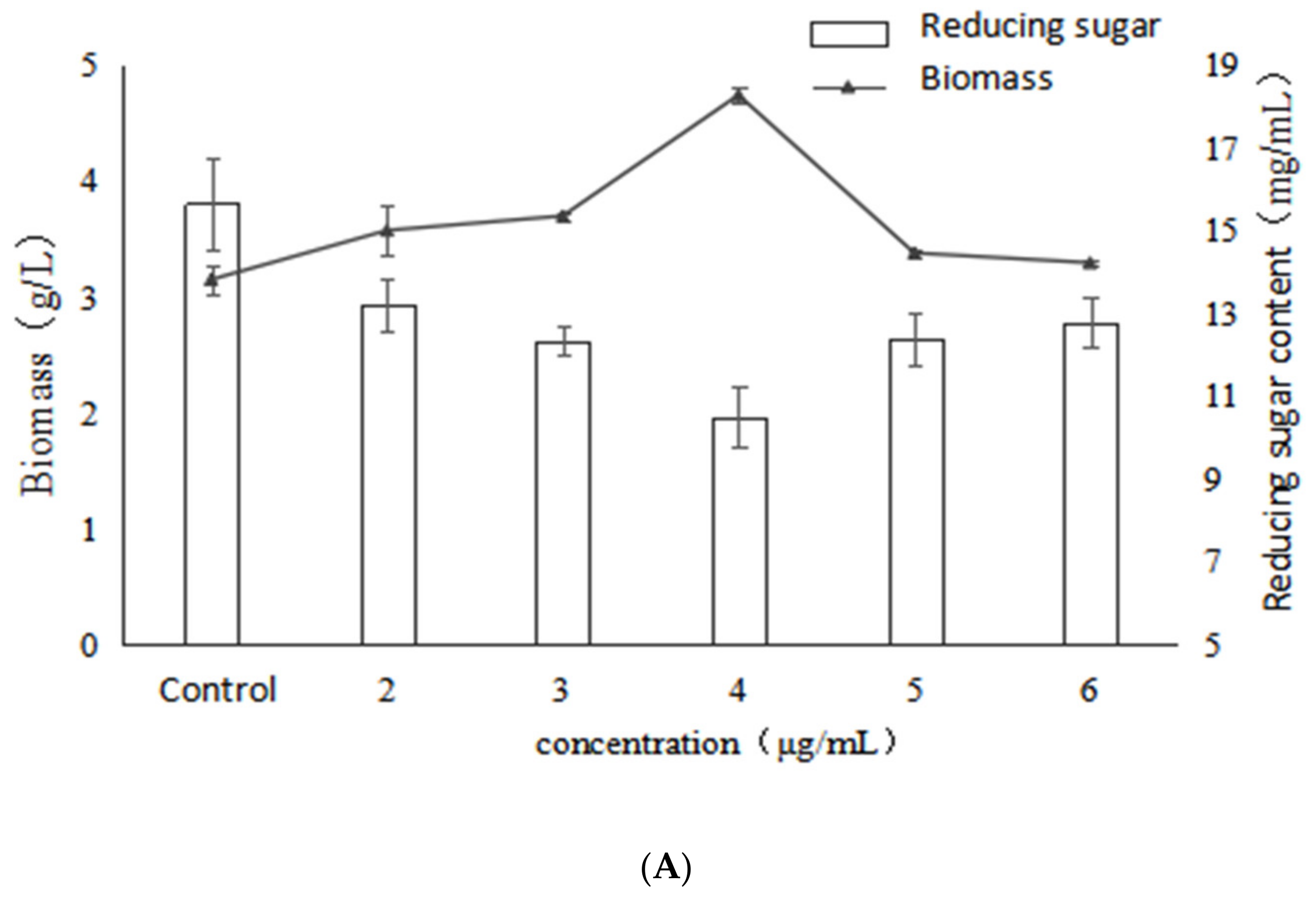
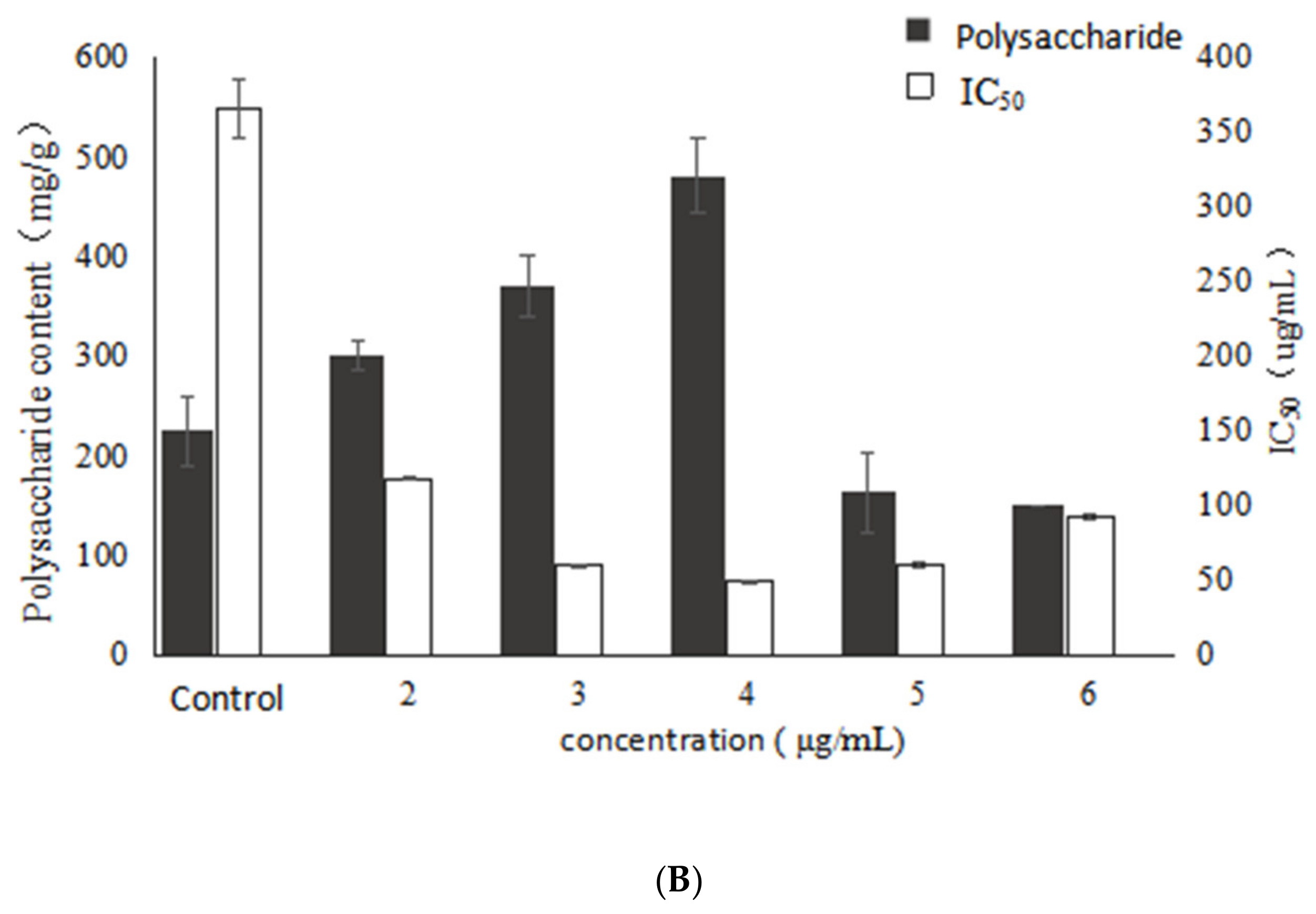
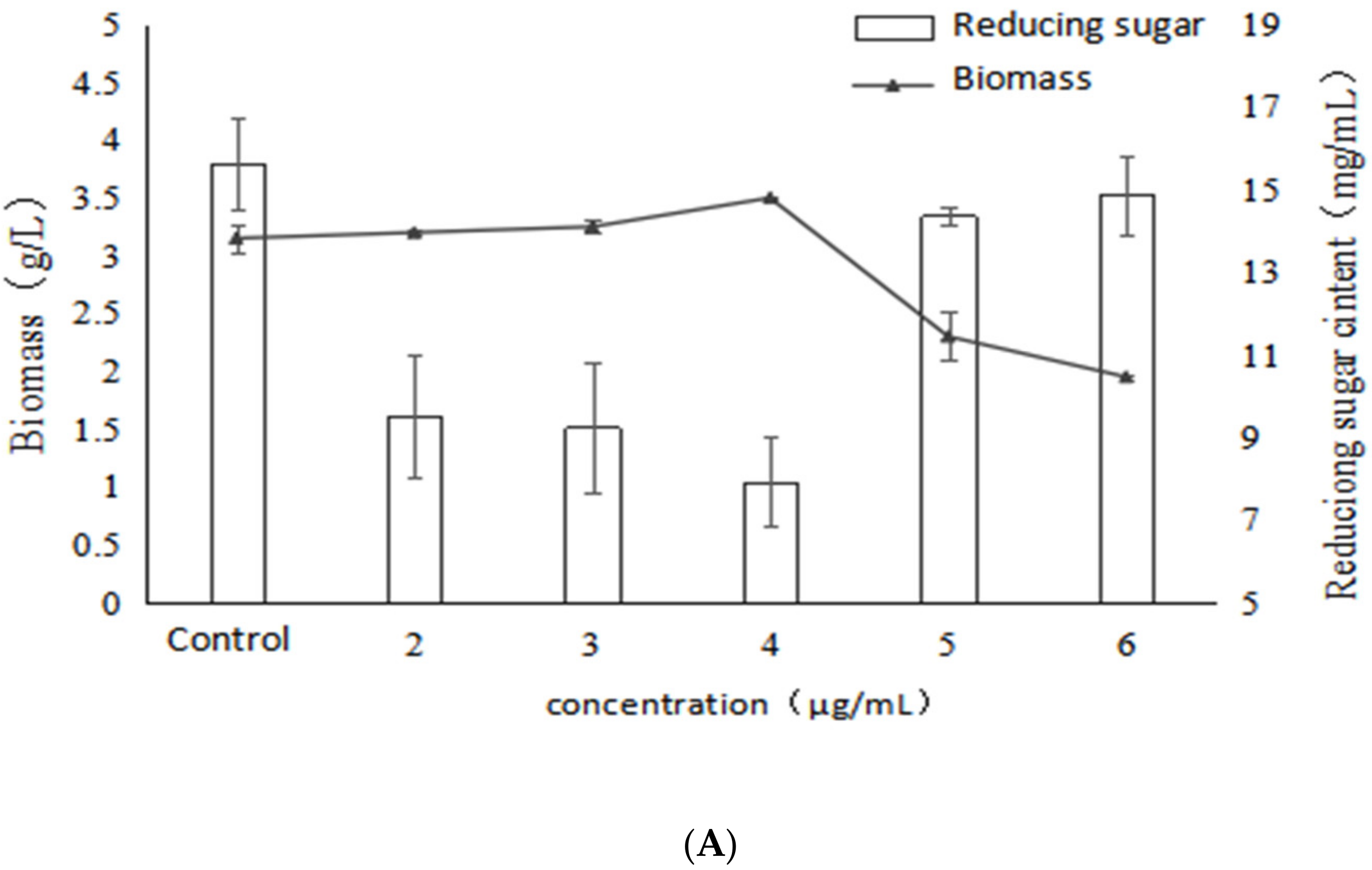
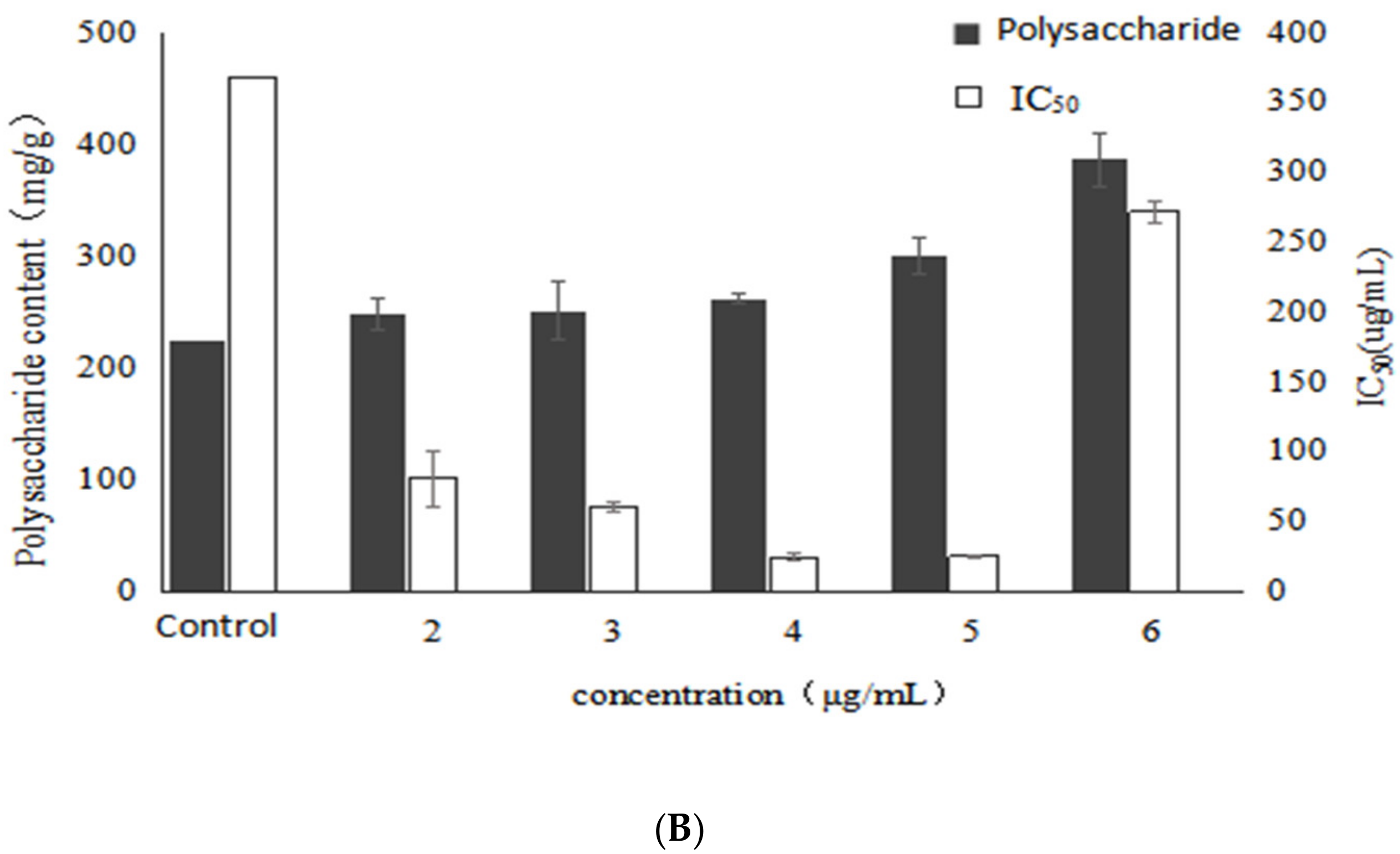
2.1. Effect of VB6 on Bioactive Exopolysaccharides by Submerged Fermentation of Inonotus Obliquus
2.2. Effect of VB1 on Bioactive Polysaccharides by Submerged Fermentation of Inonotus Obliquus
2.3. Effect of Betulin on Bioactive Polysaccharides by Submerged Fermentation of Inonotus Obliquus
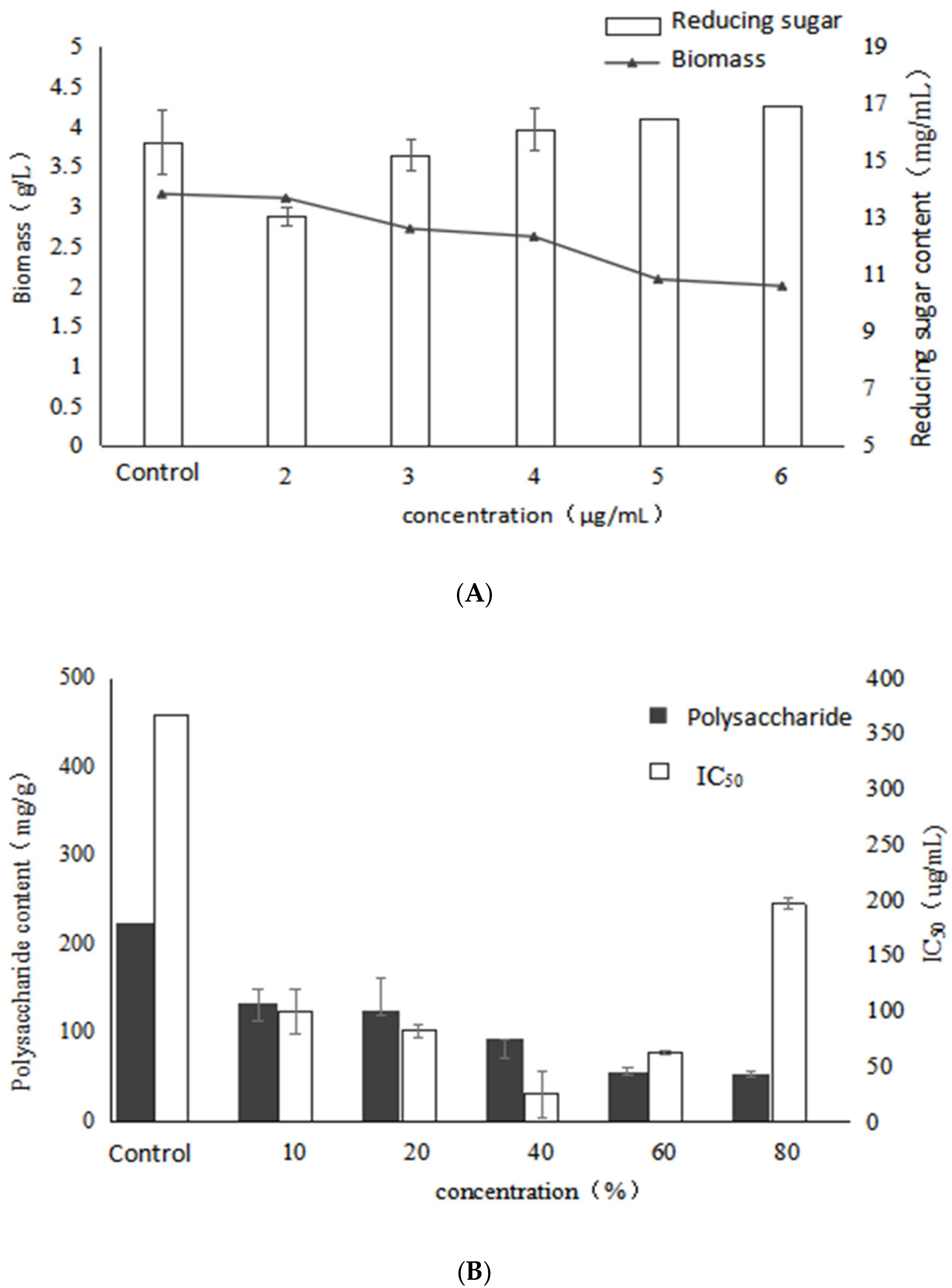
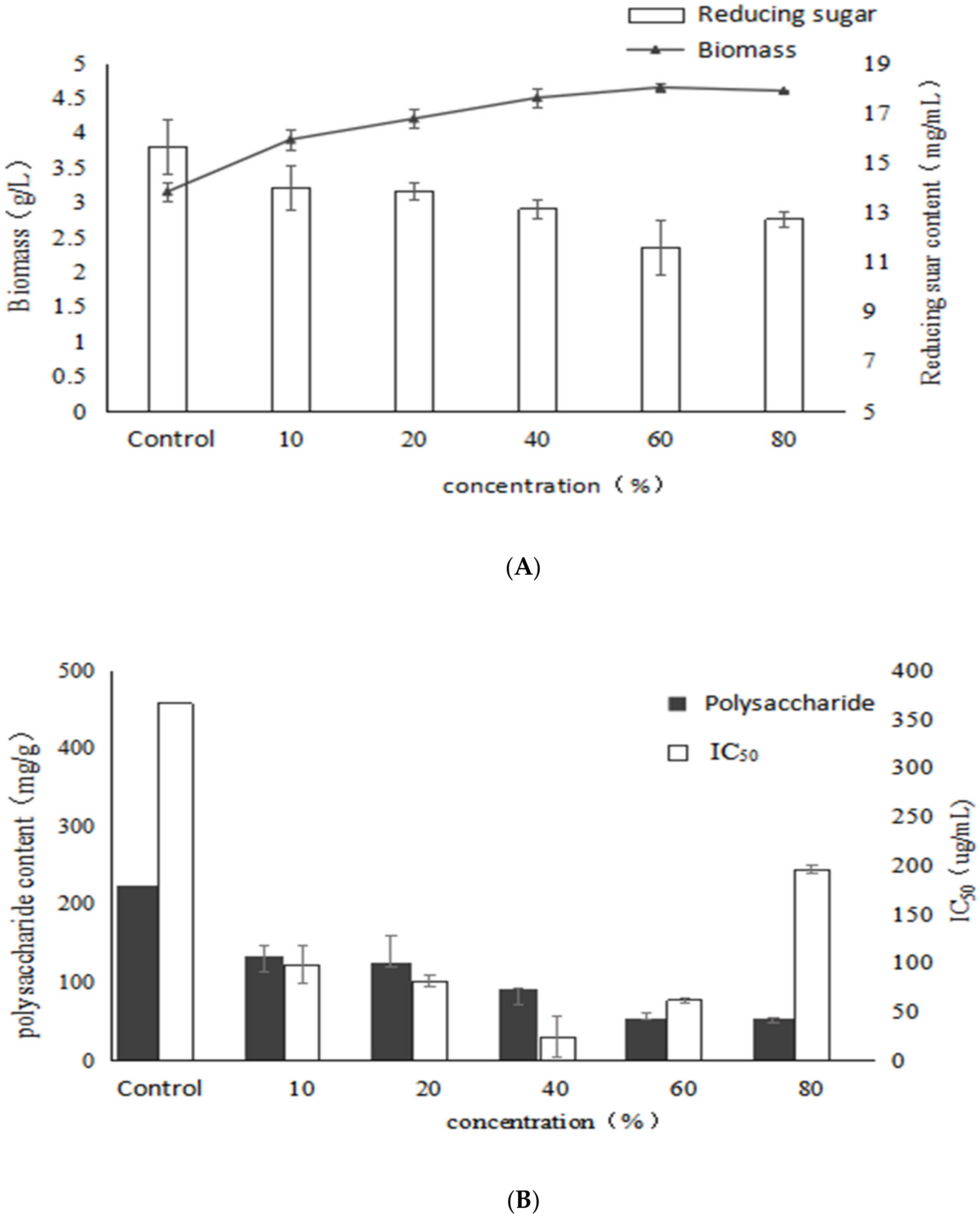
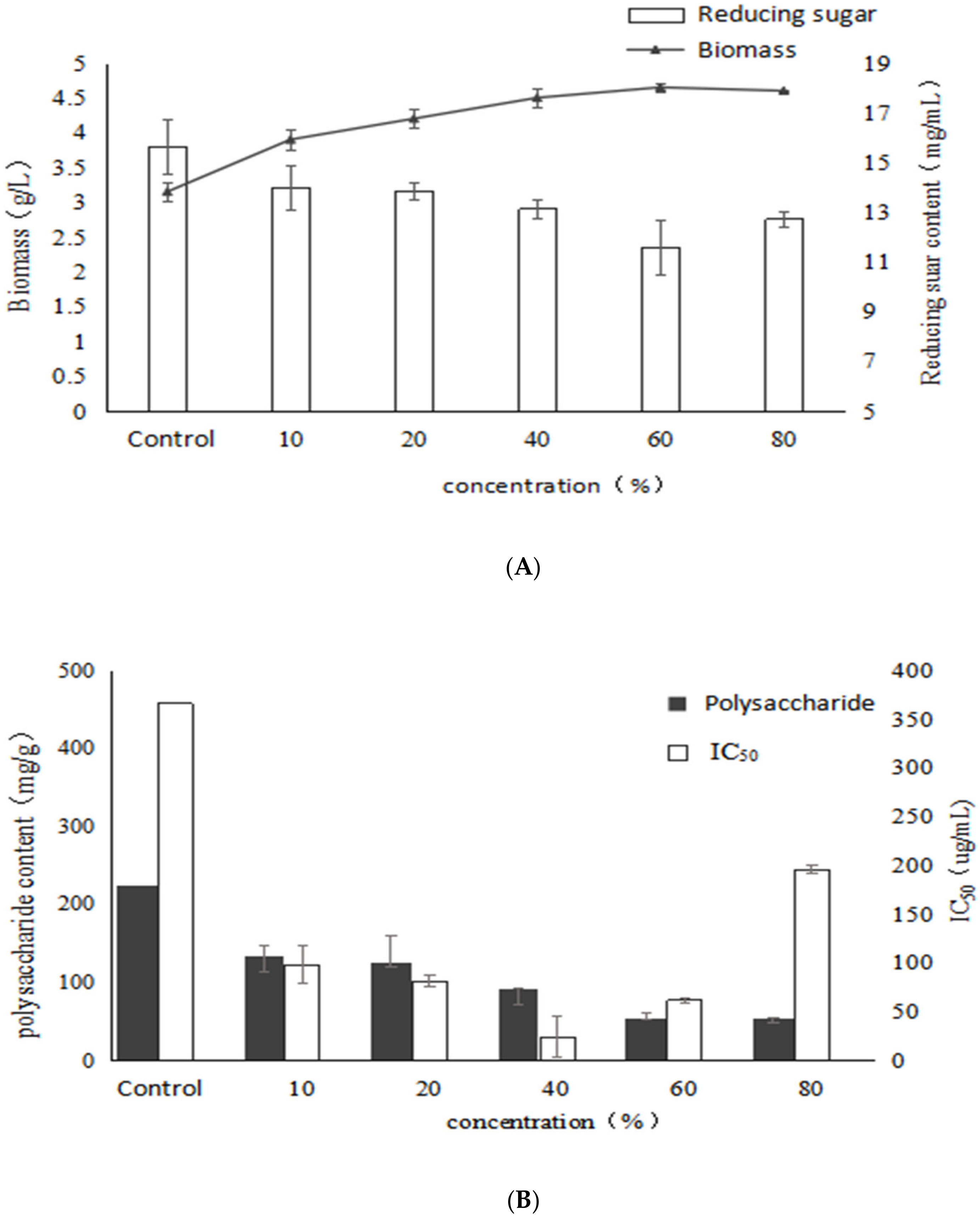
2.4. Effect of Birch Extract on Bioactive Polysaccharides by Submerged Fermentation of Inonotus Obliquus
3. Discussion
4. Materials and Methods
4.1. Materials and Regents
4.2. Submerged Fermentation
4.3. Measurement of Mycelial Biomass
4.4. Polysaccharide Content and Reducing Sugar Content in Fermentation Broth
4.5. Assay of α-Glucosidase Inhibitory Activity
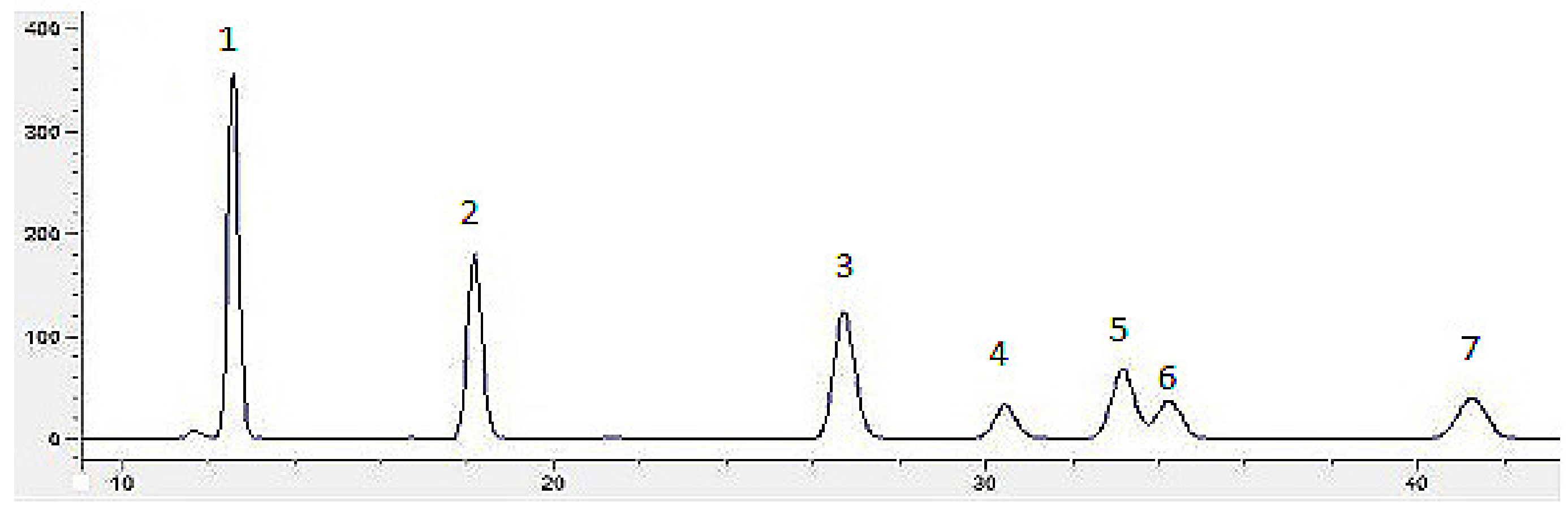
4.6. Monosaccharide Composition Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Park, Y.; Won, J.; Kim, Y.; Choi, J.; Park, H.; Lee, K. In vivo and in vitro anti-inflammatory and anti-nociceptive effects of the methanol extract of Inonotus obliquus. J. Ethnopharmacol. 2005, 101, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Hui, J.; Kou, W.; Xin, R.; Jia, F.; Wang, N. Identification of Inonotus obliquus and analysis of antioxidation and antitumor activities of polysaccharides. Curr. Microbiol. 2008, 57, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Xueming, L.; Haixia, C.; Peng, D.; Lingling, F.; Xin, Z. Phytochemical characteristics and hypoglycaemic activity of fraction from mushroom Inonotus obliquus. J. Sci. Food Agric. 2010, 90, 276–280. [Google Scholar]
- Ma, L.; Chen, H.; Dong, P.; Lu, X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom Inonotus obliquus. Food Chem. 2013, 139, 503–508. [Google Scholar] [CrossRef]
- Xu, X.; Wu, Y.; Chen, H. Comparative antioxidative characteristics of polysaccharide-enriched extracts from natural sclerotia and cultured mycelia in submerged fermentation of Inonotus obliquus. Food Chem. 2011, 127, 74–79. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Li, S.; Li, W.; Yuan, G.; Pan, Y.; Chen, H. Anti-diabetic effects of Inonotus obliquus polysaccharides in streptozotocin-induced type 2 diabetic mice and potential mechanism via PI3K-Akt signal pathway. Biomed. Pharmacother. 2017, 95, 1669–1677. [Google Scholar] [CrossRef]
- Zhang, B.; Cheung, P.C.K. Use of stimulatory agents to enhance the production of bioactive exopolysaccharide from pleurotus tuber-regium by submerged fermentation. J. Agr. Food Chem. 2011, 59, 1210–1216. [Google Scholar] [CrossRef]
- Huang, S.; Ding, S.; Fan, L. Antioxidant activities of five polysaccharides from Inonotus obliquus. Int. J. Biol. Macromol. 2012, 50, 1183–1187. [Google Scholar] [CrossRef]
- Liu, P.; Xue, J.; Tong, S.; Dong, W.; Wu, P. Structure Characterization and Hypoglycaemic Activities of Two Polysaccharides from Inonotus obliquus. Molecules 2018, 23, 1948. [Google Scholar] [CrossRef]
- Chen, H.; Yan, M.; Zhu, J.; Xu, X. Enhancement of exo-polysaccharide production and antioxidant activity in submerged cultures of Inonotus obliquus by lignocellulose decomposition. J. Ind. Microbiol. Biotechnol. 2011, 38, 291–298. [Google Scholar] [CrossRef]
- Park, J.; Kim, S.; Hwang, H.; Cho, Y.; Yun, J. Stimulatory effect of plant oils and fatty acids on the exo-biopolymer production in Cordyceps militaris. Enzyme Microb. Technol. 2002, 31, 250–255. [Google Scholar] [CrossRef]
- Xiao, J.; Xiao, D.; Xiong, Q.; Liang, Z.; Zhong, J. Nutritional requirements for the hyperproduction of bioactive exopolysaccharides by submerged fermentation of the edible medicinal fungus Cordyceps taii. Biochem. Eng. J. 2010, 49, 241–249. [Google Scholar] [CrossRef]
- Shih, I.; Tsai, K.; Hsieh, C. Effects of culture conditions on the mycelial growth and bioactive metabolite production in submerged culture of Cordyceps militaris. Biochem. Eng. J. 2007, 33, 193–201. [Google Scholar] [CrossRef]
- Bai, Y.; Feng, Y.; Mao, D.; Xu, C. Optimization for betulin production from mycelial culture of Inonotus obliquus by orthogonal design and evaluation of its antioxidant activity. J. Taiwan Inst. Chem. 2012, 43, 663–669. [Google Scholar] [CrossRef]
- Jonathan, S.G.; Fasidi, I.O. Studies on phytohormones, vitamins and mineral element requirements of Lentinus subnudus (Berk) and schizophyllum commune (Fr. Ex. Fr) from Nigeria. Food Chem. 2001, 75, 303–307. [Google Scholar] [CrossRef]
- Xu, C.; Shu, C. Medium Optimization for Producing Bioactive Exopolysaccharides by Agaricus brasiliensis S. Wasser et al. (=A. blazei Murrill ss. Heinem) in Submerged Culture. Food Technol. Biotechnol. 2007, 45, 327–333. [Google Scholar]
- Cong, W.; Weiwei, L.; Zhongqin, C.; Xudong, G.; Guoqi, Y.; Yuxiang, P.; Haixia, C. Effects of simulated gastrointestinal digestion in vitro on the chemical properties, antioxidant activity, α-amylase and α-glucosidase inhibitory activity of polysaccharides from Inonotus obliquus. Food Res. Int. 2018, 103, 280–288. [Google Scholar]
- Xu, X.; Shen, M.; Quan, L. Stimulatory Agents Simultaneously Improving the Production and Antioxidant Activity of Polyphenols from Inonotus obliquus by Submerged Fermentation. Appl. Biochem. Biotechnol. 2015, 176, 1237–1250. [Google Scholar] [CrossRef]
- Wang, L.; Lu, Z.; Geng, Y.; Zhang, X.; Xu, G.; Shi, J.; Xu, Z. Stimulated production of steroids in Inonotus obliquus by host factors from birch. J. Biosci. Bioeng. 2014, 118, 728–731. [Google Scholar] [CrossRef]
- Liu, G.; Xiao, H.; Wang, X.; Zhao, Y.; Zhang, Y.; Ren, G. Stimulated Production of Triterpenoids of Ganoderma lucidum by an Ether Extract from the Medicinal Insect, Catharsius molossus, and Identification of the Key Stimulating Active Components. Appl. Biochem. Biotechnol. 2011, 165, 87–97. [Google Scholar] [CrossRef]
- Zhao, G.; Yan, W.; Cao, D. Simultaneous determination of betulin and betulinic acid in white birch bark using RP-HPLC. J. Pharm. Biomed. 2007, 43, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.O.; Han, S.B.; Lee, H.W.; Ahn, H.J.; Yoon, Y.D.; Jung, J.K.; Kim, H.M.; Shin, C.S. Immuno-stimulating effect of the endo-polysaccharide produced by submerged culture of Inonotus obliquus. Life Sci. 2005, 77, 2438–2456. [Google Scholar] [CrossRef] [PubMed]
- Babitskaia, V.G.; Shcherba, V.V.; Ikonnikova, N.V. Melanin complex of the fungus Inonotus obliquus. Appl. Biochem. Microbiol. 2000, 36, 439. [Google Scholar]
- Xu, X.; Zhao, W.; Shen, M. Antioxidant activity of liquid cultured Inonotus obliquus polyphenols using tween-20 as a stimulatory agent: Correlation of the activity and the phenolic profiles. J. Taiwan Inst. Chem. 2016, 69, 41–47. [Google Scholar] [CrossRef]
- Xu, X.; Hu, Y.; Zhu, L. The capability of Inonotus obliquus for lignocellulosic biomass degradation in peanut shell and for simultaneous production of bioactive polysaccharides and polyphenols in submerged fermentation. J. Taiwan Inst. Chem. 2014, 45, 2851–2858. [Google Scholar] [CrossRef]
- Jonathan, S.G.; Fasidi, I.O. Effect of carbon, nitrogen and mineral sources on growth of Psathyerella atroumbonata (Pegler), a Nigerian edible mushroom. Food Chem. 2001, 72, 479–483. [Google Scholar] [CrossRef]
- Xue, J.; Tong, S.; Wang, Z.; Liu, P. Chemical characterization and hypoglycaemic activities in vitro of two polysaccharides from Inonotus obliquus by submerged culture. Molecules 2018, 23, 3261. [Google Scholar] [CrossRef]
- Zhang, A.; Shen, Y.; Cen, M.; Hong, X.; Shao, Q.; Chen, Y.; Zheng, B. Polysaccharide and crocin contents, and antioxidant activity of saffron from different origins. Ind. Crop. Prod. 2019, 133, 111–117. [Google Scholar] [CrossRef]
- Yang, F.; Ke, Y.; Kuo, S. Effect of fatty acids on the mycelial growth and polysaccharide formation by Ganoderma lucidum in shake flask cultures. Enzyme Microb. Technol. 2000, 27, 295–301. [Google Scholar] [CrossRef]
- Kim, J.; Hyun, T.K.; Kim, M. The inhibitory effects of ethanol extracts from sorghum, foxtail millet and proso millet on α-glucosidase and α-amylase activities. Food. Chem. 2011, 124, 1647–1651. [Google Scholar] [CrossRef]
- Malinowska, E.; Klimaszewska, M.; Strączek, T.; Schneider, K.; Kapusta, C.; Podsadni, P.; Łapienis, G.; Dawidowski, M.; Kleps, J.; Górska, S.; et al. Selenized polysaccharides—Biosynthesis and structural analysis. Carbohydr. Polym. 2018, 198, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jiang, W.; Lu, J.; Yu, Y.; Wu, B. Analysis of the monosaccharide composition of water-soluble polysaccharides from Sargassum fusiforme by high performance liquid chromatography/electrospray ionisation mass spectrometry. Food Chem. 2014, 145, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Quan, L.; Shen, M. Effect of chemicals on production, composition and antioxidant activity of polysaccharides of Inonotus obliquus. Int. J. Biol. Macromol. 2015, 77, 143–150. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations (μg/mL) | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|
| Mannose (mol%) | 18.83 | 18.40 | 21.66 | 26.79 | 28.63 |
| Rhamnose (mol%) | 19.45 | 23.94 | 27.93 | 26.25 | 24.26 |
| Glucose (mol%) | 33.84 | 25.01 | 15.14 | 12.94 | 12.57 |
| Galactose (mol%) | - | - | - | - | - |
| Xylose (mol%) | - | - | - | - | - |
| Arabinose (mol%) | - | - | - | - | - |
| Fucose (mol%) | 27.88 | 32.65 | 35.27 | 34.02 | 34.54 |
| Concentrations (μg/mL) | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|
| Mannose (mol%) | 13.18 | 20.76 | 19.84 | 19.13 | 18.61 |
| Rhamnose (mol%) | 17.07 | 27.13 | 17.07 | 19.26 | 20.84 |
| Glucose (mol%) | 25.01 | 26.08 | 31.97 | 32.44 | 32.68 |
| Galactose (mol%) | 19.61 | - | - | - | - |
| Xylose (mol%) | - | - | - | - | - |
| Arabinose (mol%) | - | - | - | - | - |
| Fucose (mol%) | 25.13 | 26.03 | 31.12 | 29.17 | 27.87 |
| Concentrations (μg/mL) | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|
| Mannose (mol%) | 10.09 | 13.23 | 13.67 | 12.39 | 11.10 |
| Rhamnose (mol%) | 23.30 | 22.63 | 22.78 | 19.46 | 13.47 |
| Glucose (mol%) | 18.35 | 27.17 | 28.67 | 29.25 | 44.63 |
| Galactose (mol%) | - | - | - | - | - |
| Xylose (mol%) | - | - | - | - | - |
| Arabinose (mol%) | 14.50 | 15.71 | 13.05 | 10.98 | 15.34 |
| Fucose (mol%) | 33.76 | 21.26 | 21.84 | 26.92 | 15.46 |
| Concentrations (%) | 10 | 20 | 40 | 60 | 80 |
|---|---|---|---|---|---|
| Mannose (mol%) | 15.80 | 10.39 | 13.45 | 13.03 | 21.68 |
| Rhamnose (mol%) | 11.25 | 15.31 | 15.83 | 16.78 | 20.01 |
| Glucose (mol%) | 54.89 | 26.99 | 27.08 | 23.79 | 35.14 |
| Galactose (mol%) | - | 17.00 | 17.32 | 17.91 | - |
| Xylose (mol%) | - | 13.62 | 14.11 | 12.04 | - |
| Arabinose (mol%) | 18.07 | 15.99 | 12.21 | 16.44 | 23.09 |
| Fucose (mol%) | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Zhao, Z.; Zhou, X.; Hu, J.; Xue, J.; Liu, X.; Zhang, J.; Liu, P.; Tong, S. Simultaneous Use of Stimulatory Agents to Enhance the Production and Hypoglycaemic Activity of Polysaccharides from Inonotus obliquus by Submerged Fermentation. Molecules 2019, 24, 4400. https://doi.org/10.3390/molecules24234400
Wang M, Zhao Z, Zhou X, Hu J, Xue J, Liu X, Zhang J, Liu P, Tong S. Simultaneous Use of Stimulatory Agents to Enhance the Production and Hypoglycaemic Activity of Polysaccharides from Inonotus obliquus by Submerged Fermentation. Molecules. 2019; 24(23):4400. https://doi.org/10.3390/molecules24234400
Chicago/Turabian StyleWang, Mengya, Zhezhen Zhao, Xia Zhou, Jinrong Hu, Jiao Xue, Xiao Liu, Jingsheng Zhang, Ping Liu, and Shisheng Tong. 2019. "Simultaneous Use of Stimulatory Agents to Enhance the Production and Hypoglycaemic Activity of Polysaccharides from Inonotus obliquus by Submerged Fermentation" Molecules 24, no. 23: 4400. https://doi.org/10.3390/molecules24234400
APA StyleWang, M., Zhao, Z., Zhou, X., Hu, J., Xue, J., Liu, X., Zhang, J., Liu, P., & Tong, S. (2019). Simultaneous Use of Stimulatory Agents to Enhance the Production and Hypoglycaemic Activity of Polysaccharides from Inonotus obliquus by Submerged Fermentation. Molecules, 24(23), 4400. https://doi.org/10.3390/molecules24234400