
Curdlan (Alcaligenes faecalis) (1→3)-β-d-Glucan Oligosaccharides Drive M1 Phenotype Polarization in Murine Bone Marrow-Derived Macrophages via Activation of MAPKs and NF-κB Pathways

 ,
,
Abstract
1. Introduction
2. Results
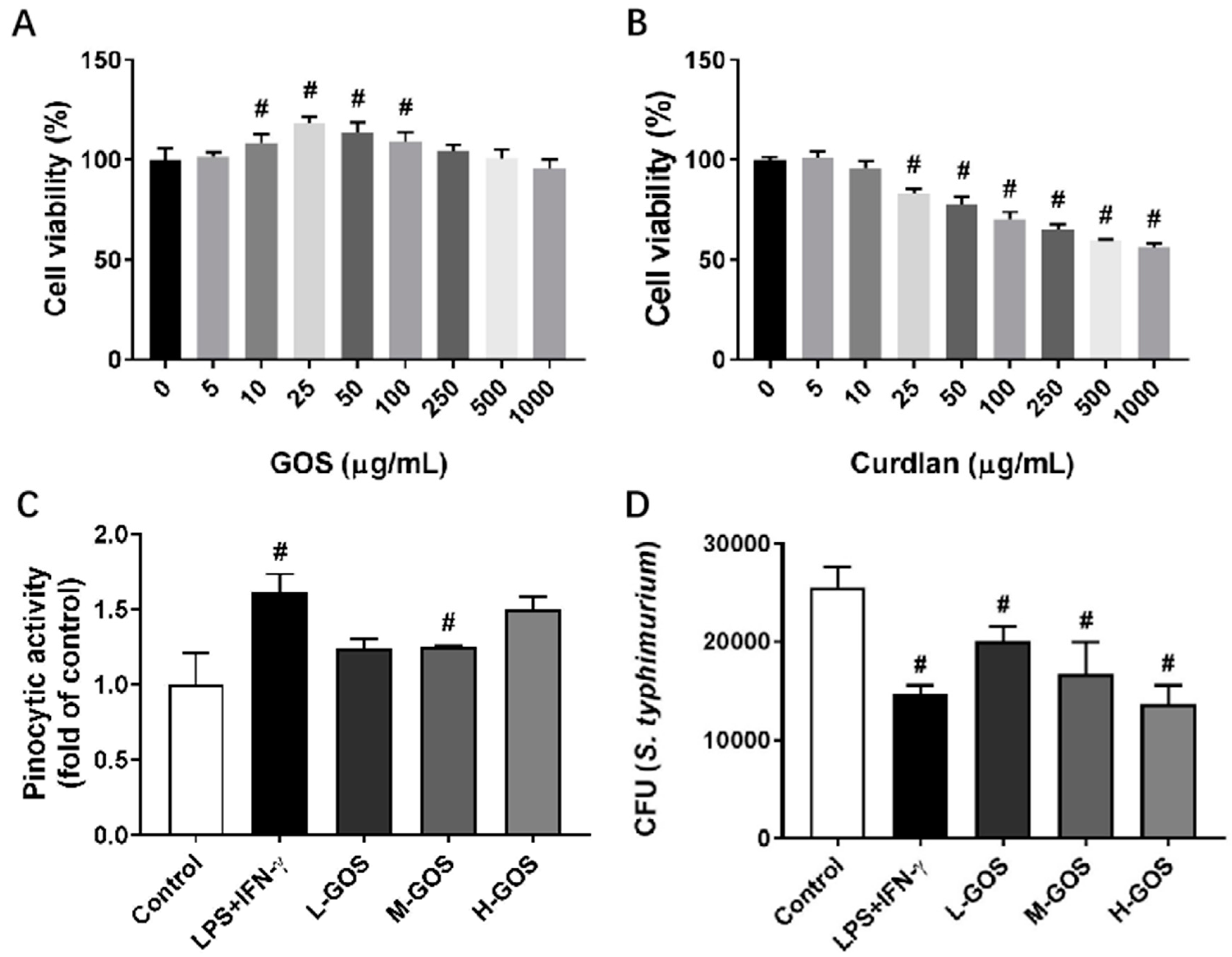
2.1. Proliferation, Pinocytosis, and Bactericidal Function of BMDMs
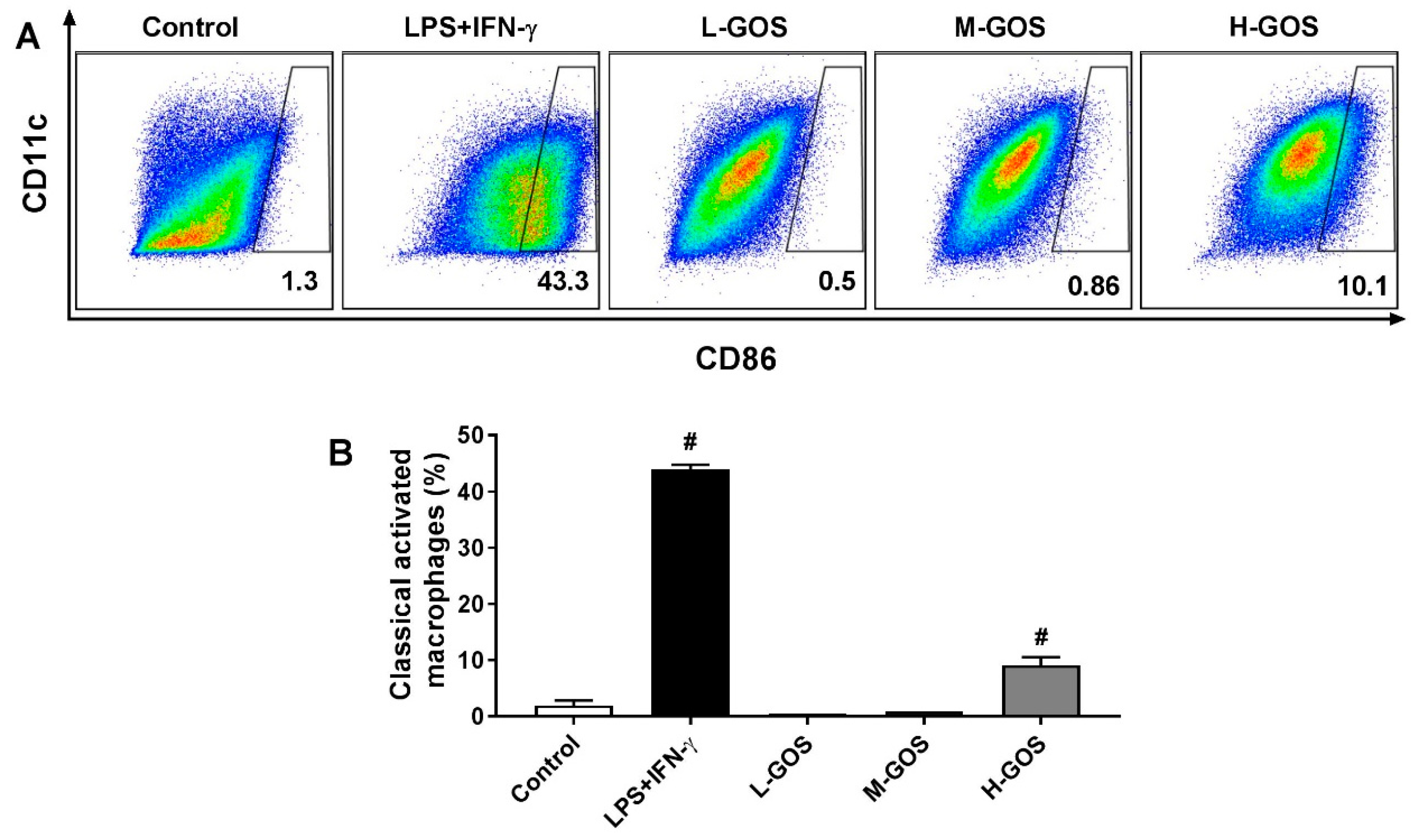
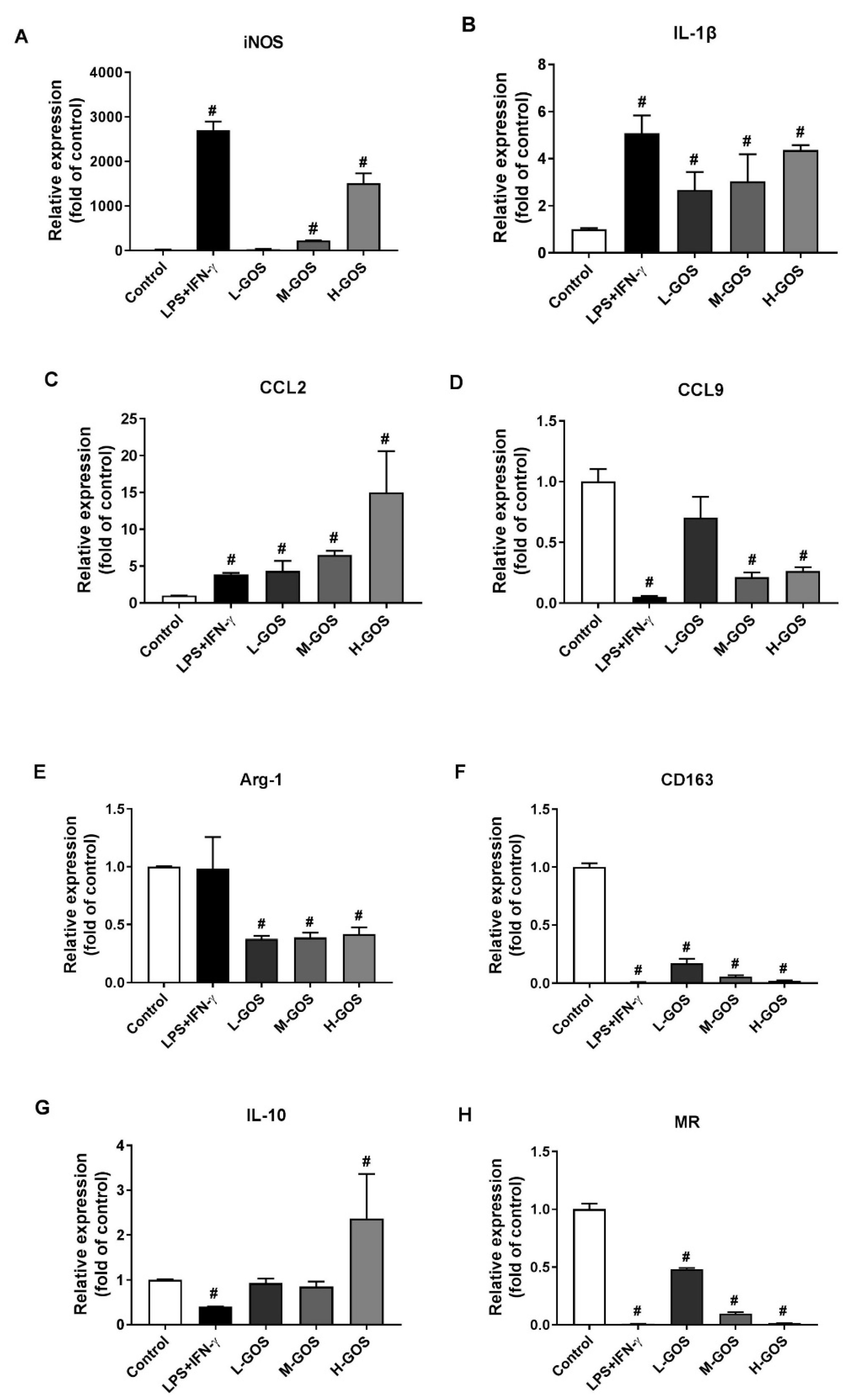
2.2. GOS Promoted M1 Phenotype Polarization of BMDMs
2.3. GOS Induced Marker Expression on BMDMs’ Surface
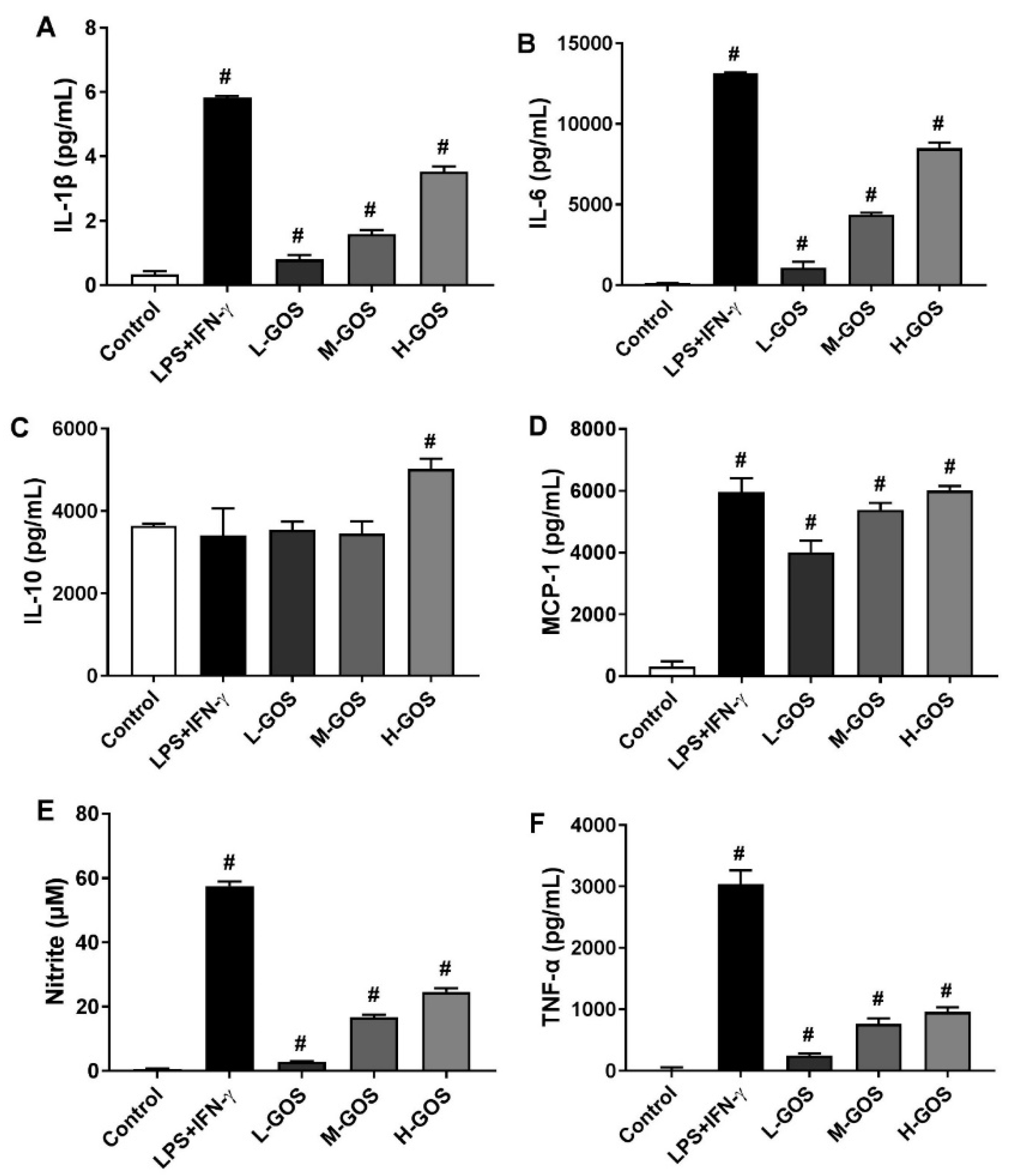
2.4. GOS Enhanced Cytokines and Nitrite Release of BMDMs
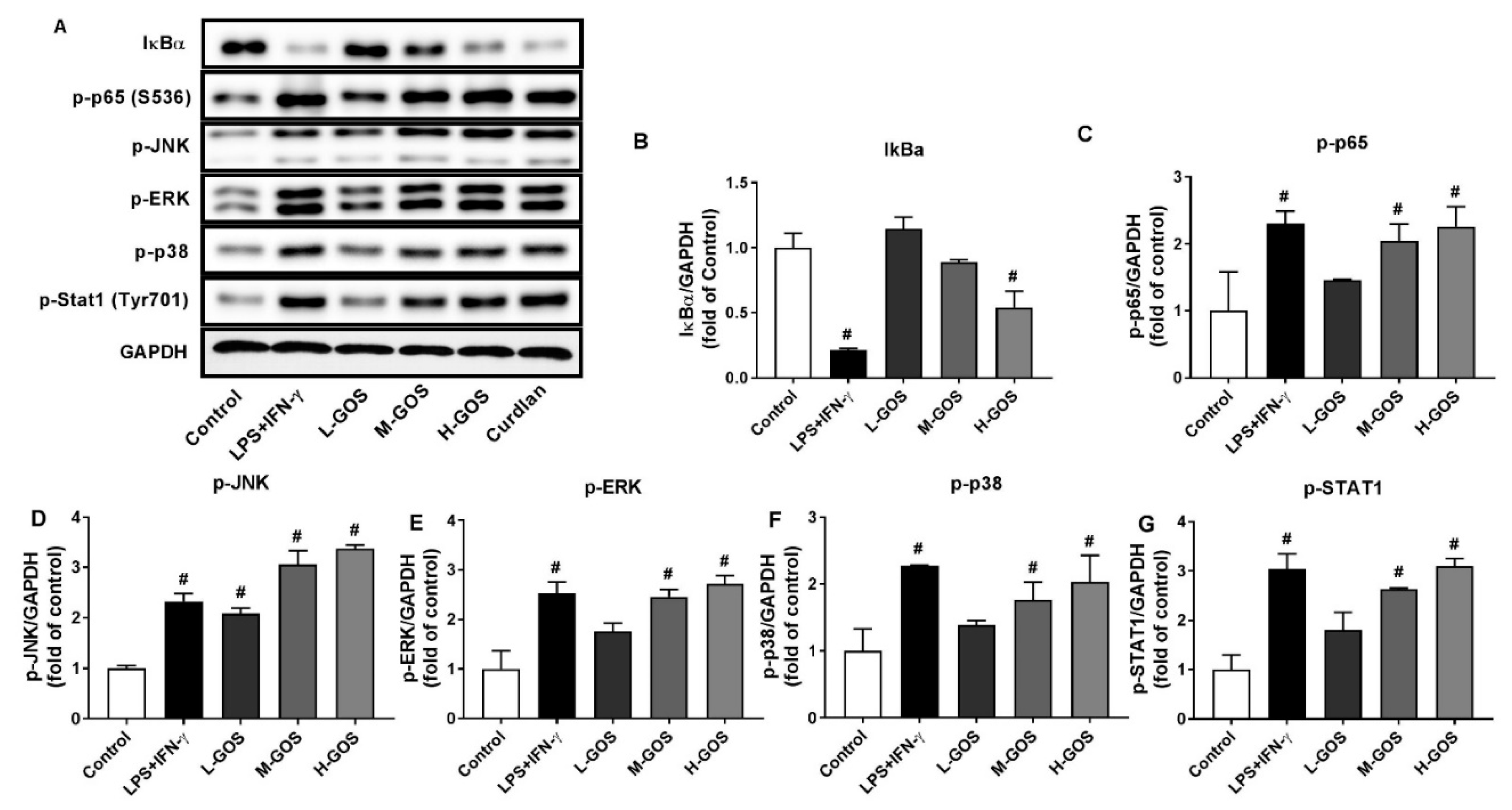
2.5. GOS Activated Mitogen-Activated Protein Kinases (MAPKs) and Nuclear Factor-Kappa B (NF-κB) Pathways in BMDMs
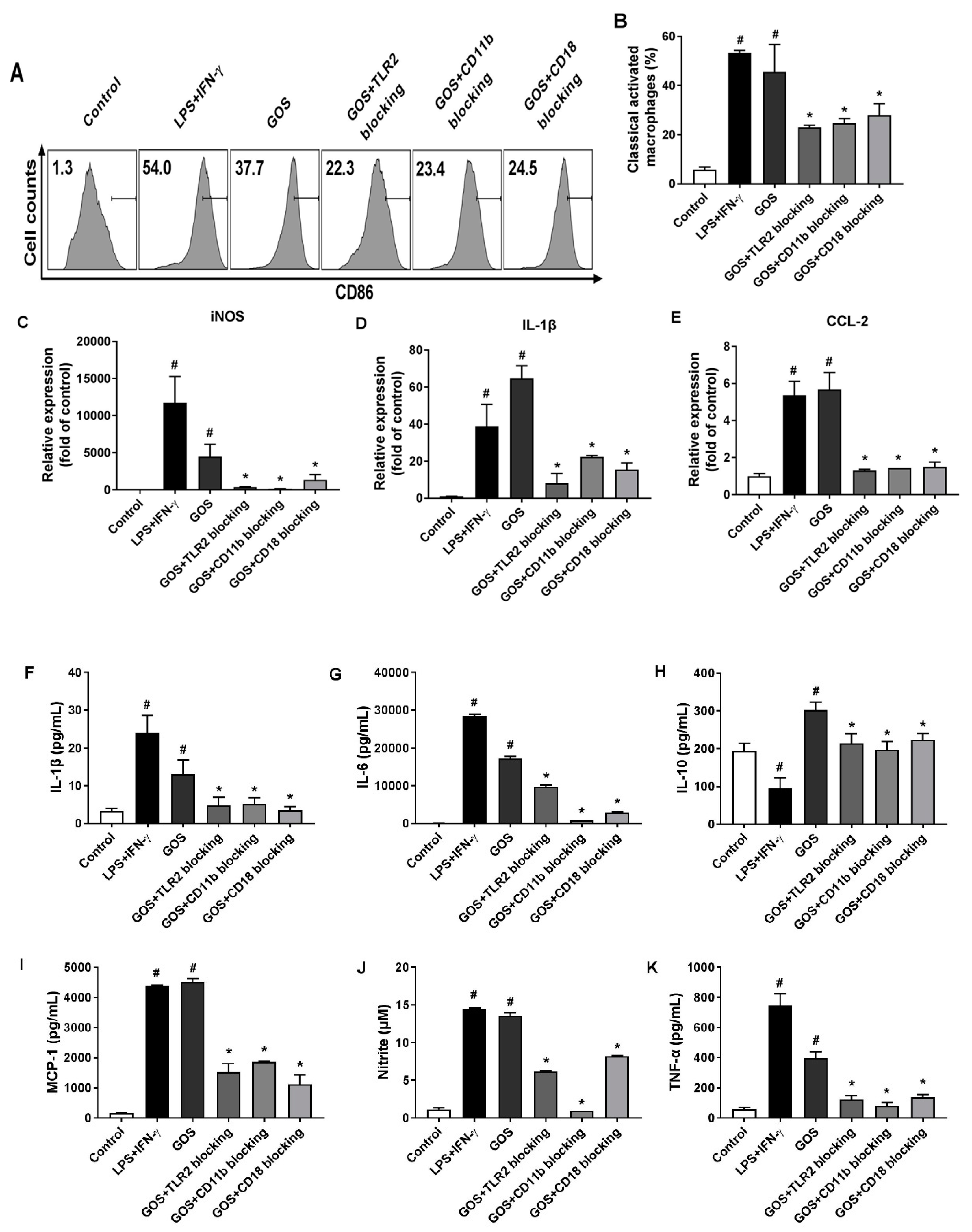
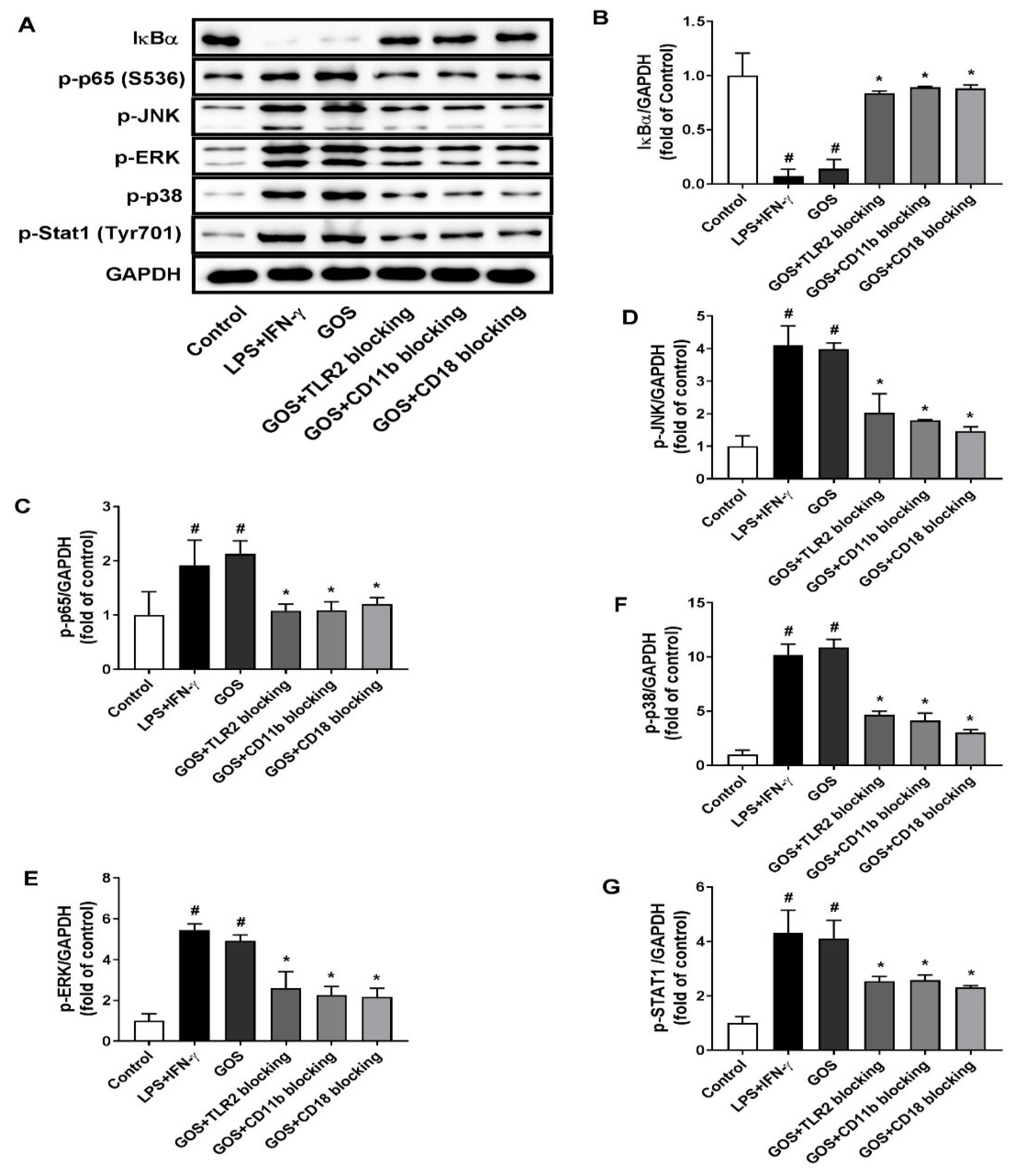
2.6. GOS Mediated M1 Phenotype Polarization was Suppressed after Receptor Blocking
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation and Differentiation of Bone Marrow Cells
4.3. Cytotoxicity Assay
4.4. Pinocytosis and Bacterial Killing Assays
4.5. Flow Cytometric Analysis
4.6. Nitrite and Cytokine Production Measurement
4.7. Quantitative Real Time-Polymerase Chain Reaction
4.8. Western Blot Assay
4.9. Receptor Blocking Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.H.; Smith, W.; Hao, D.J.; He, B.R.; Kong, L.B. M1 and M2 macrophage polarization and potentially therapeutic naturally occurring compounds. Int. Immunopharmacol. 2019, 70, 459–466. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Thapa, B.; Lee, K. Metabolic influence on macrophage polarization and pathogenesis. Bmb. Rep. 2019, 52, 360–372. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage polarization: Different gene signatures in M1(LPS+) vs. classically and M2(LPS-) vs. alternatively activated macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, Q.; Yang, T.; Ding, W.; Zhao, Y. Cellular metabolism and macrophage functional polarization. Int. Rev. Immunol. 2015, 34, 82–100. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Benoit, M.; Desnues, B.; Mege, J.L. Macrophage polarization in bacterial infections. J. Immunol. 2008, 181, 3733–3739. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Wang, L.X.; Zhang, S.X.; Wu, H.J.; Rong, X.L.; Guo, J. M2b macrophage polarization and its roles in diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef]
- Koh, Y.C.; Yang, G.L.; Lai, C.S.; Weerawatanakorn, M.; Pan, M.H. Chemopreventive effects of phytochemicals and medicines on M1/M2 polarized macrophage role in inflammation-related diseases. Int. J. Biol. Macromol. 2018, 19, 2208. [Google Scholar] [CrossRef]
- Ren, J.Y.; Hou, C.L.; Shi, C.C.; Lin, Z.H.; Liao, W.Z.; Yuan, E.D. A polysaccharide isolated and purified from Platycladus orientalis (L.) Franco leaves, characterization, bioactivity and its regulation on macrophage polarization. Carbohyd. Polym. 2019, 213, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.K.; Wang, Y.; Wu, Y.P.; Chen, H.L.; Zheng, S.S.; Li, Y.L.; Xu, X.; Li, W.F. Echinacea purpurea extract polarizes M1 macrophages in murine bone marrow-derived macrophages through the activation of JNK. J. Cell Biochem. 2017, 118, 2664–2671. [Google Scholar] [CrossRef] [PubMed]
- Minato, K.; Laan, L.C.; van Die, I.; Mizuno, M. Pleurotus citrinopileatus polysaccharide stimulates anti-inflammatory properties during monocyte-to-macrophage differentiation. Int. J. Biol. Macromol. 2019, 122, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.X.; Jing, Y.S.; Zhou, Q.Q.; Hu, X.J.; Zhu, J.H.; Guo, Z.Y. Structural elucidation and immunostimulatory activity of a new polysaccharide from Cordyceps militaris. Food Funct. 2018, 9, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Wu, W.J.; Ren, Y.; Li, X.F.; Tang, Y.Q.; Min, T.; Lai, F.R.; Wu, H. Structural characterization of a novel polysaccharide from Lepidium meyenii (Maca) and analysis of its regulatory function in macrophage polarization in vitro. J. Agric. Food Chem. 2017, 65, 1146–1157. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Z.P.; Bian, Z.X.; Han, Q.B. Astragalus polysaccharide RAP induces macrophage phenotype polarization to M1 via the Notch signaling pathway. Molecules 2019, 24, 2016. [Google Scholar] [CrossRef]
- Ma, Y.H.; Xing, Y.Y.; Mi, H.W.; Guo, Z.Q.; Lu, Y.Y.; Xi, T. Extraction, preliminary characterization and immunostimulatory activity in vitro of a polysaccharide isolated from Strongylocentrotus nudus eggs. Carbohyd. Polym. 2014, 111, 576–583. [Google Scholar] [CrossRef]
- Brown, G.D.; Herre, J.; Williams, D.L.; Willment, J.A.; Marshall, A.S.; Gordon, S. Dectin-1 mediates the biological effects of beta-glucans. J. Exp. Med. 2003, 197, 1119–1124. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. Beta-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef]
- Liu, M.; Luo, F.L.; Ding, C.L.; Albeituni, S.; Hu, X.L.; Ma, Y.F.; Cai, Y.H.; McNally, L.; Sanders, M.A.; Jain, D.; et al. Dectin-1 activation by a natural product beta-glucan converts immunosuppressive macrophages into an M1-like phenotype. J. Immunol. 2015, 195, 5055–5065. [Google Scholar] [CrossRef]
- Smith, A.J.; Graves, B.; Child, R.; Rice, P.J.; Ma, Z.C.; Lowman, D.W.; Ensley, H.E.; Ryter, K.T.; Evans, J.T.; Williams, D.L. Immunoregulatory activity of the natural product laminarin varies widely as a result of its physical properties. J. Immunol. 2018, 200, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.X.; Zhang, J.F. Bacterial glucans: Production, properties, and applications. Appl. Microbiol. Biot. 2016, 100, 9023–9036. [Google Scholar] [CrossRef] [PubMed]
- Funami, T.; Yada, H.; Nakao, Y. Curdlan properties for application in fat mimetic for meat products. J. Food Sci. 1998, 63, 283–287. [Google Scholar] [CrossRef]
- Funami, T.; Yotsusuka, F.; Nakao, Y. Thermo-irreversible characteristics of curdlan gels in a model reduced fat pork sausage. J. Food Sci. 1998, 63, 575–579. [Google Scholar] [CrossRef]
- Spicer, E.J.F.; Goldenthal, E.; Ikeda, T. A toxicological assessment of curdlan. Food Chem. Toxicol. 1999, 37, 455–479. [Google Scholar] [CrossRef]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary modulation of immune function by β-glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef]
- Barbosa-Lorenzi, V.C.; Peyda, S.; Scheynius, A.; Nilsson, G.; Lunderius-Andersson, C. Curdlan induces selective mast cell degranulation without concomitant release of LTC4, IL-6 or CCL2. Immunobiology 2017, 222, 647–650. [Google Scholar] [CrossRef]
- Rui, K.; Tian, J.; Tang, X.Y.; Ma, J.; Xu, P.; Tian, X.Y.; Wang, Y.G.; Xu, H.X.; Lu, L.W.; Wang, S.J. Curdlan blocks the immune suppression by myeloid-derived suppressor cells and reduces tumor burden. Immunol. Res. 2016, 64, 931–939. [Google Scholar] [CrossRef]
- Xiao, M.; Jiang, M.F.; Wu, K.; Yang, H.; Ni, X.W.; Yan, W.L.; Phillips, G.O.; Jiang, F.T. Investigation on curdlan dissociation by heating in water. Food Hydrocoll. 2017, 70, 57–64. [Google Scholar] [CrossRef]
- Shi, Y.Q.; Liu, J.; Yan, Q.J.; You, X.; Yang, S.Q.; Jiang, Z.Q. In vitro digestibility and prebiotic potential of curdlan (1→3)-beta-D-glucan oligosaccharides in Lactobacillus species. Carbohyd. Polym. 2018, 188, 17–26. [Google Scholar] [CrossRef]
- Hida, T.H.; Ishibashi, K.; Miura, N.N.; Adachi, Y.; Shirasu, Y.; Ohno, N. Cytokine induction by a linear 1,3-glucan, curdlan-oligo, in mouse leukocytes in vitro. Inflamm. Res. 2009, 58, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.Q.; Zhen, H.M.; Wang, N.N.; Yan, Q.J.; Jing, H.; Jiang, Z.Q. Curdlan oligosaccharides having higher immunostimulatory activity than curdlan in mice treated with cyclophosphamide. Carbohyd. Polym. 2019, 207, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Gissibl, A.; Care, A.; Parker, L.M.; Iqbal, S.; Hobba, G.; Nevalainen, H.; Nevalainen, H.; Sunna, A. Microwave pretreatment of paramylon enhances the enzymatic production of soluble β-1,3-glucans with immuno-stimulatory activity. Carbohyd. Polym. 2018, 196, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Mano, M.C.R.; Neri-Numa, I.A.; da Silva, J.B.; Paulino, B.N.; Pessoa, M.G.; Pastore, G.M. Oligosaccharide biotechnology: An approach of prebiotic revolution on the industry. Appl. Microbiol. Biot. 2018, 102, 17–37. [Google Scholar] [CrossRef]
- Xu, Q.; Chao, Y.L.; Wan, Q.B. Health benefit application of functional oligosaccharides. Carbohyd. Polym. 2009, 77, 435–441. [Google Scholar]
- Zhao, C.; Wu, Y.J.; Liu, X.Y.; Liu, B.; Cao, H.; Yu, H.; Sarker, S.D.; Nahar, L.; Xiao, J.B. Functional properties, structural studies and chemo-enzymatic synthesis of oligosaccharides. Trends Food Sci. Technol. 2017, 66, 135–145. [Google Scholar] [CrossRef]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF-κB activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef]
- Ji, J.; Hu, S.L.; Cui, Z.W.; Li, W.F. Probiotic Bacillus amyloliquefaciens mediate M1 macrophage polarization in mouse bone marrow-derived macrophages. Arch. Microbiol. 2013, 195, 349–356. [Google Scholar] [CrossRef]
- Mao, Y.L.; Wang, B.K.; Xu, X.; Du, W.; Li, W.F.; Wang, Y.M. Glycyrrhizic acid promotes M1 macrophage polarization in murine bone marrow-derived macrophages associated with the activation of JNK and NF-κB. Mediat. Inflamm. 2015, 2015, 372931. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-κB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.R.; Martinez-Pomares, L.; Stacey, M.; Lin, H.; Brown, G.D.; Gordon, S. Macrophage receptors and immune recognition. Annu. Rev. Immunol. 2005, 23, 901–944. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.H.; Gringhuis, S.I. Signaling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zou, S.W.; Xu, H.; Liu, Q.Y.; Song, J.H.; Xu, M.; Xu, X.J.; Zhang, L.N. The linear structure of beta-glucan from baker’s yeast and its activation of macrophage-like RAW264.7 cells. Carbohyd. Polym. 2016, 148, 61–68. [Google Scholar] [CrossRef]
- Zhou, P.; Chen, Z.Z.; Yan, Q.J.; Yang, S.Q.; Hilgenfeld, R.; Jiang, Z.Q. The structure of a glycoside hydrolase family 81 endo-beta-1,3-glucanase. Acta. Cryst. D 2013, 69, 2027–2038. [Google Scholar] [CrossRef]
- Zhang, M.M.; Wang, G.; Lai, F.R.; Wu, H. Structural characterization and immunomodulatory activity of a novel polysaccharide from Lepidium meyenii. J. Agric. Food Chem. 2016, 64, 1921–1931. [Google Scholar] [CrossRef]
- Schmolz, L.; Wallert, M.; Lorkowski, S. Optimized incubation regime for nitric oxide measurements in murine macrophages using the Griess assay. J. Immunol. Methods 2017, 449, 68–70. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds curdlan (1→3)-β-d-glucan oligosaccharides (GOS) are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Primer sequences (5′−3′) | |
|---|---|---|
| Forward | Reverse | |
| ARG-1 | TGAGAGACCACGGGGACCTG | GCACCACACTGACTCTTCCATTC |
| IL-10 | CTCTTACTGACTGGCATGAGGAT | GAGTCGGTTAGCAGTATGTTGT |
| CD163 | CCTCCTCATTGTCTTCCTCCTGTG | CATCCGCCTTTGAATCCATCTCTTG |
| CD206 | TCAGCTATTGGACGCGAGGCA | TCCGGGTTGCAAGTTGCCGT |
| IL-1β | GCAACTGTTCCTGAACTCAACT | ATCTTTTGGGGTCCGTCAACT |
| iNOS | CACCTTGGAGTTCACCCAGT | ACCACTCGTACTTGGGATGC |
| CCL-2 | TTAAAAACCTGGATCGGAACCAA | GCATTAGCTTCAGATTTACGGGT |
| CCL-9 | CCCTCTCCTTCCTCATTCTTACA | AGTCTTGAAAGCCCATGTGAAA |
| GAPDH | GTGCTGAGTATGTCGTGGAGTCTAC | GGCGGAGATGATGACCCTTTTGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Tang, J.; Li, X.; Yan, Q.; Ma, J.; Jiang, Z. Curdlan (Alcaligenes faecalis) (1→3)-β-d-Glucan Oligosaccharides Drive M1 Phenotype Polarization in Murine Bone Marrow-Derived Macrophages via Activation of MAPKs and NF-κB Pathways. Molecules 2019, 24, 4251. https://doi.org/10.3390/molecules24234251
Liu J, Tang J, Li X, Yan Q, Ma J, Jiang Z. Curdlan (Alcaligenes faecalis) (1→3)-β-d-Glucan Oligosaccharides Drive M1 Phenotype Polarization in Murine Bone Marrow-Derived Macrophages via Activation of MAPKs and NF-κB Pathways. Molecules. 2019; 24(23):4251. https://doi.org/10.3390/molecules24234251
Chicago/Turabian StyleLiu, Jun, Jiqing Tang, Xiuting Li, Qiaojuan Yan, Junwen Ma, and Zhengqiang Jiang. 2019. "Curdlan (Alcaligenes faecalis) (1→3)-β-d-Glucan Oligosaccharides Drive M1 Phenotype Polarization in Murine Bone Marrow-Derived Macrophages via Activation of MAPKs and NF-κB Pathways" Molecules 24, no. 23: 4251. https://doi.org/10.3390/molecules24234251
APA StyleLiu, J., Tang, J., Li, X., Yan, Q., Ma, J., & Jiang, Z. (2019). Curdlan (Alcaligenes faecalis) (1→3)-β-d-Glucan Oligosaccharides Drive M1 Phenotype Polarization in Murine Bone Marrow-Derived Macrophages via Activation of MAPKs and NF-κB Pathways. Molecules, 24(23), 4251. https://doi.org/10.3390/molecules24234251