
PI3K–AKT Signaling Activation and Icariin: The Potential Effects on the Perimenopausal Depression-Like Rat Model

 , ,
, ,
Abstract
:1. Introduction
2. Results
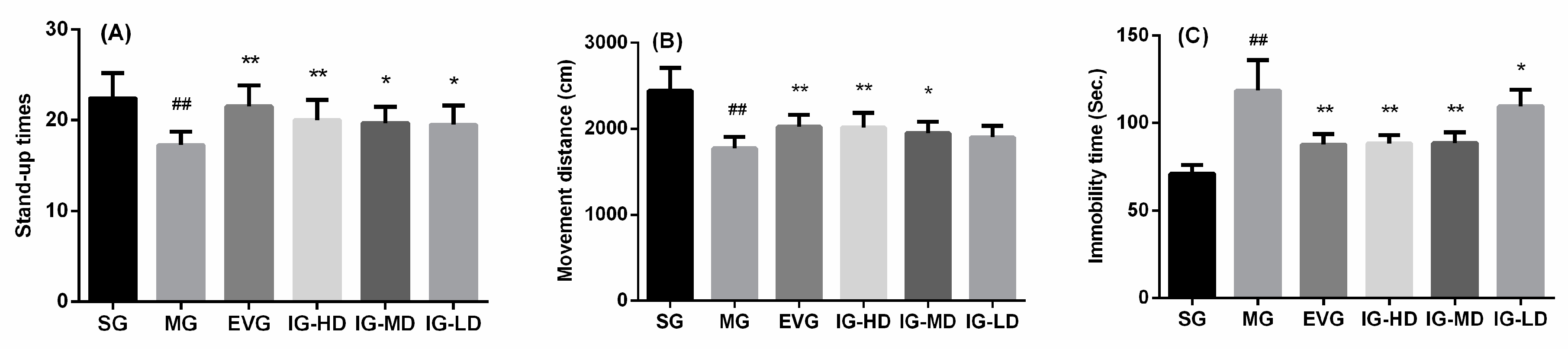
2.1. Effects of Icariin on Behavioral Changes in the Perimenopausal Depression Rats
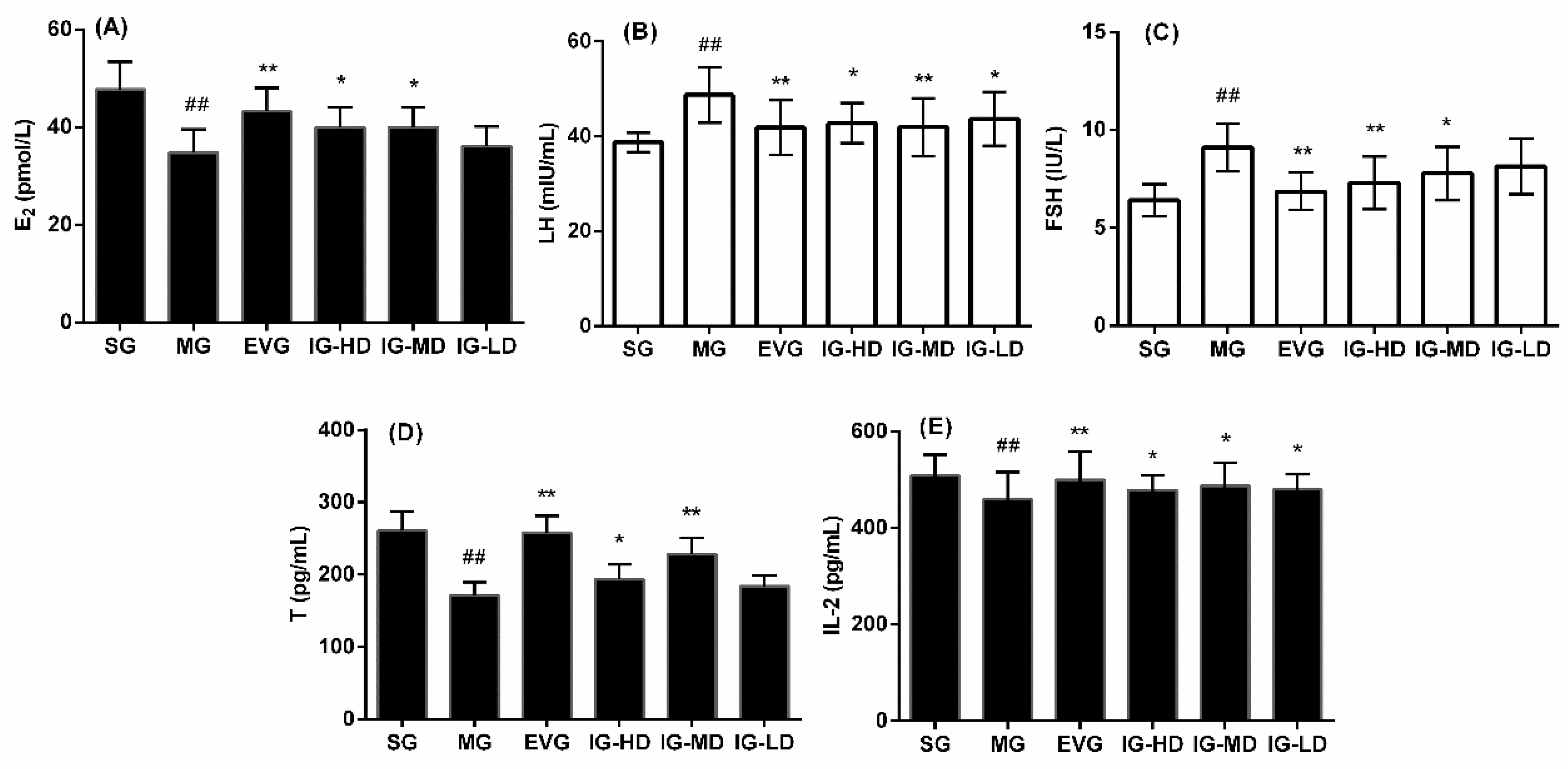
2.2. Effects of Icariin on Serum Biochemical Indicators in the Perimenopausal Depression Rats
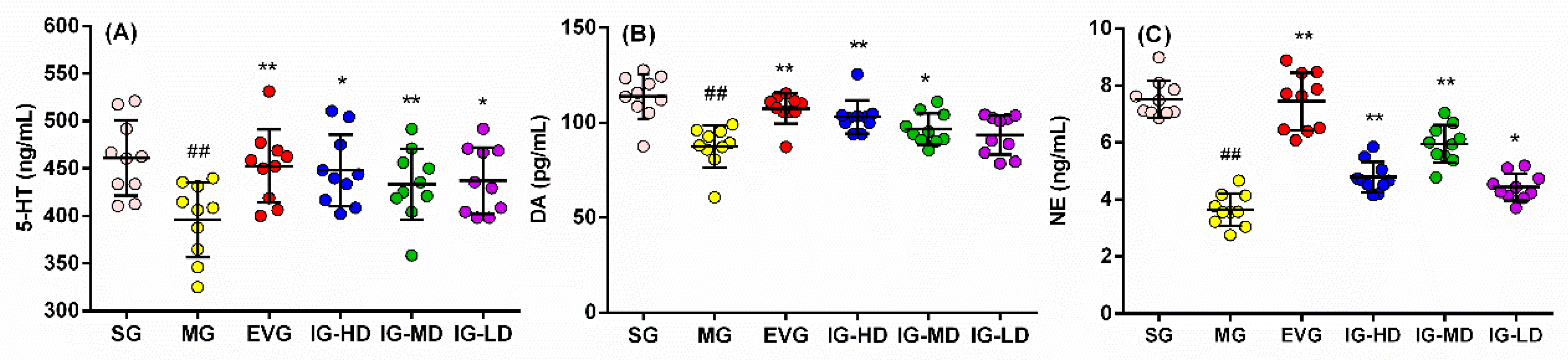
2.3. Effects of Icariin on Serum Biochemical Indicators in the Perimenopausal Depression Rats
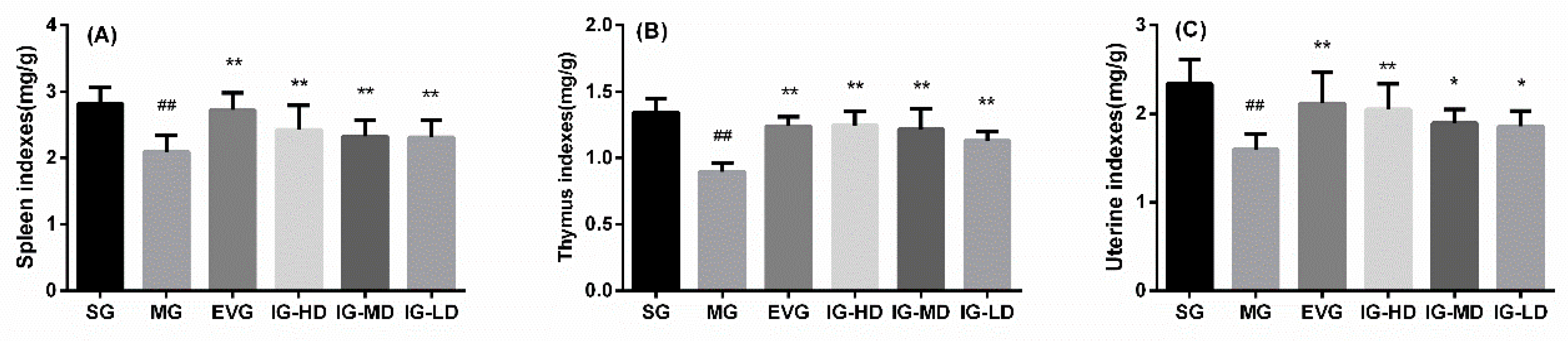
2.4. Effects of Icariin on Visceral Tissue in the Perimenopausal Depression Rats
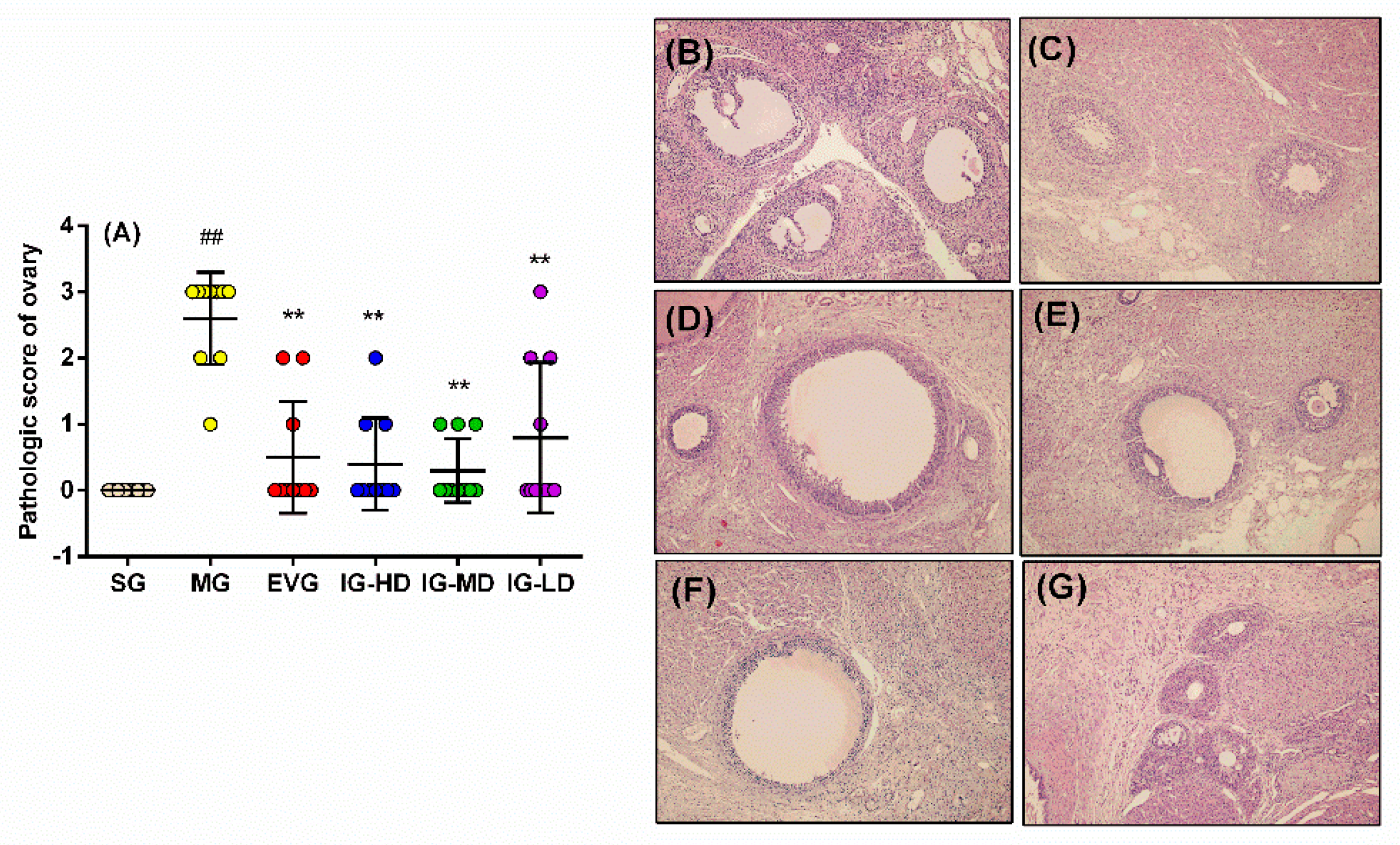
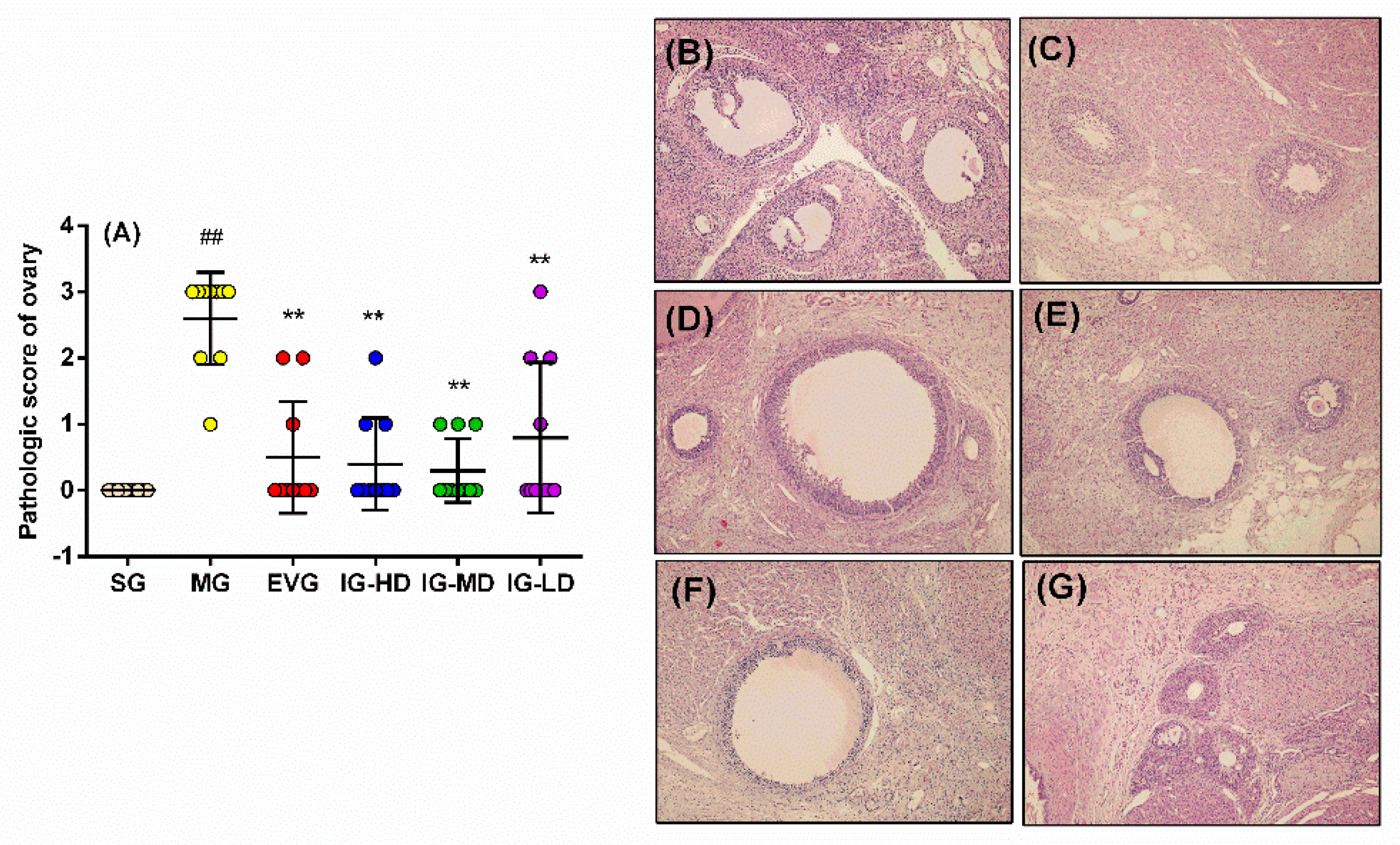
2.5. Effects of Icariin on the Pathological Changes of Ovary in the Perimenopausal Depression Rats
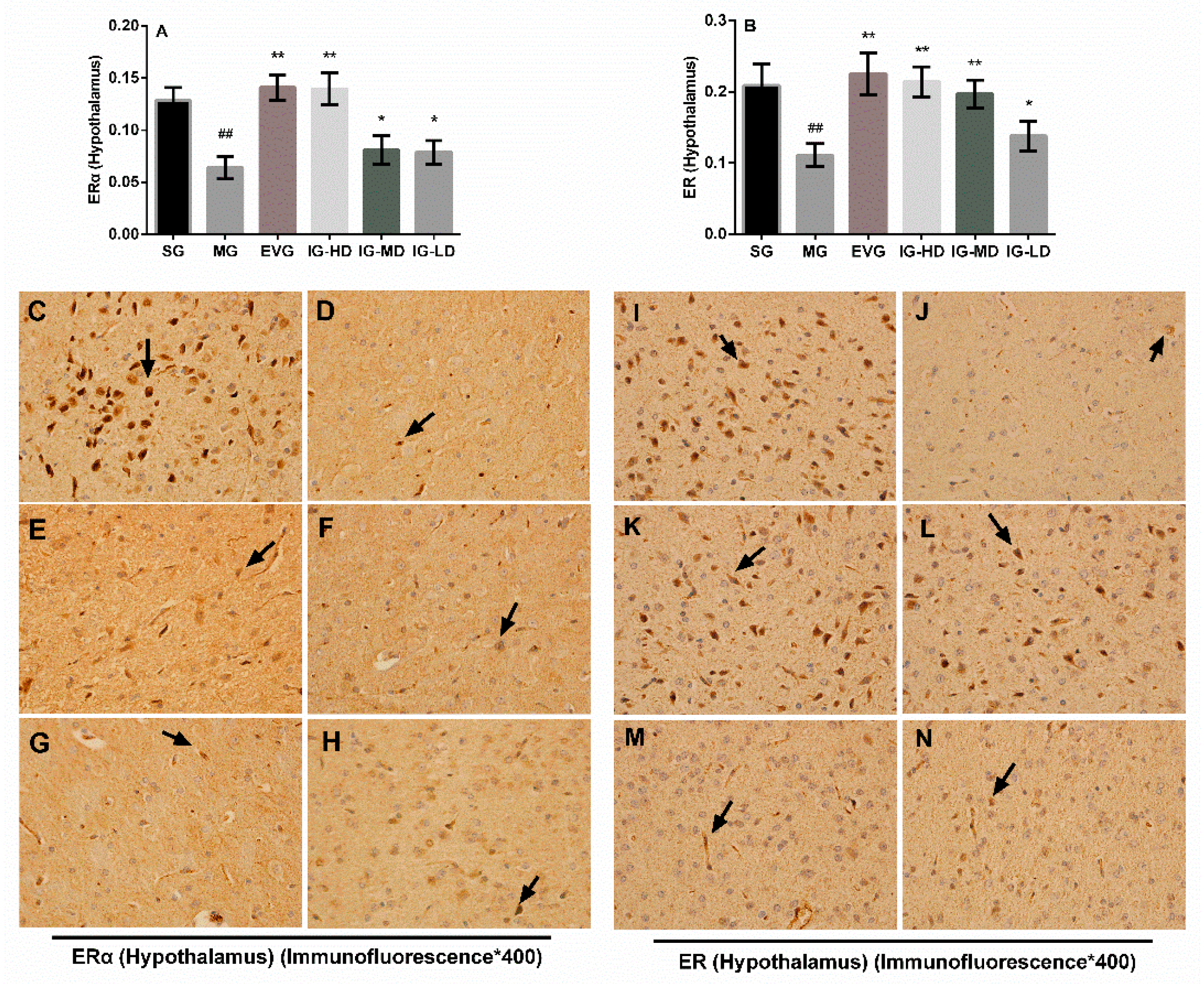
2.6. Effects of Icariin on the Expression of Hypothalamus ER and ERα in the Perimenopausal Depression Rats
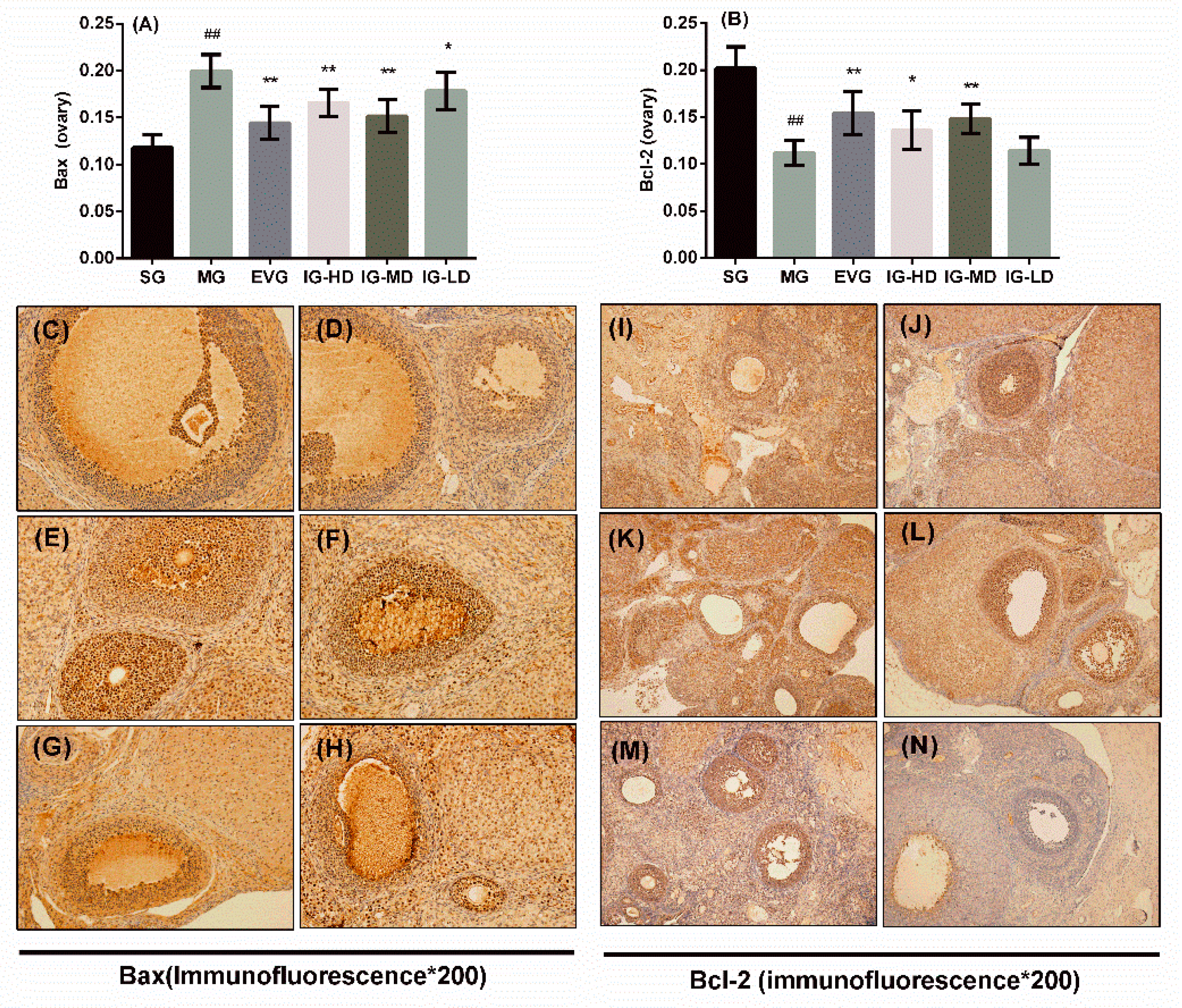
2.7. Effects of Icariin on the Expression of Ovary Bax and Bcl-2 in the Perimenopausal Depression Rats
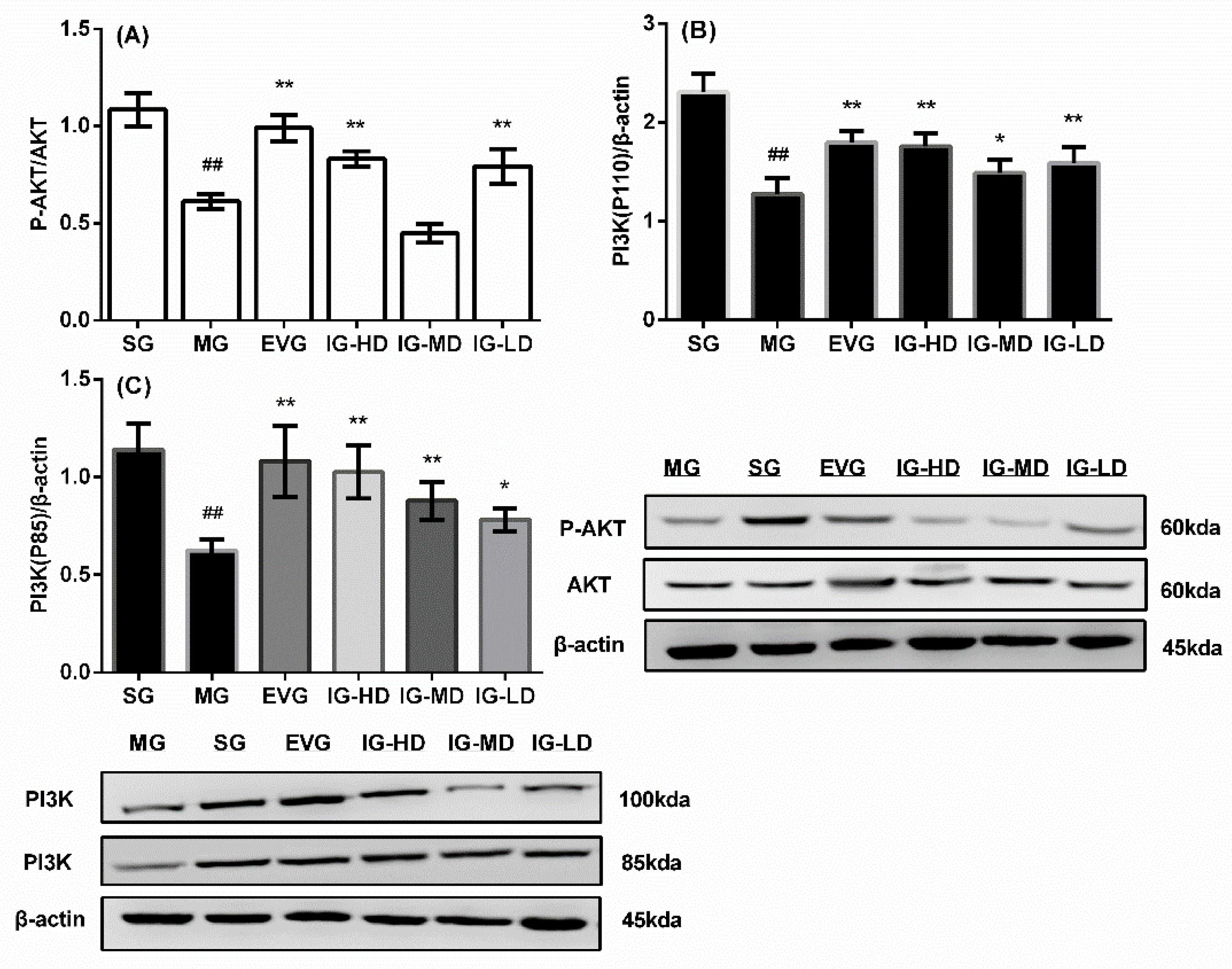
2.8. Effects of Icariin on the Expression of Ovaries p-AKT/AKT, PI3K (110 kd) and PI3K (85 kd) in the Perimenopausal Depression Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Chemicals
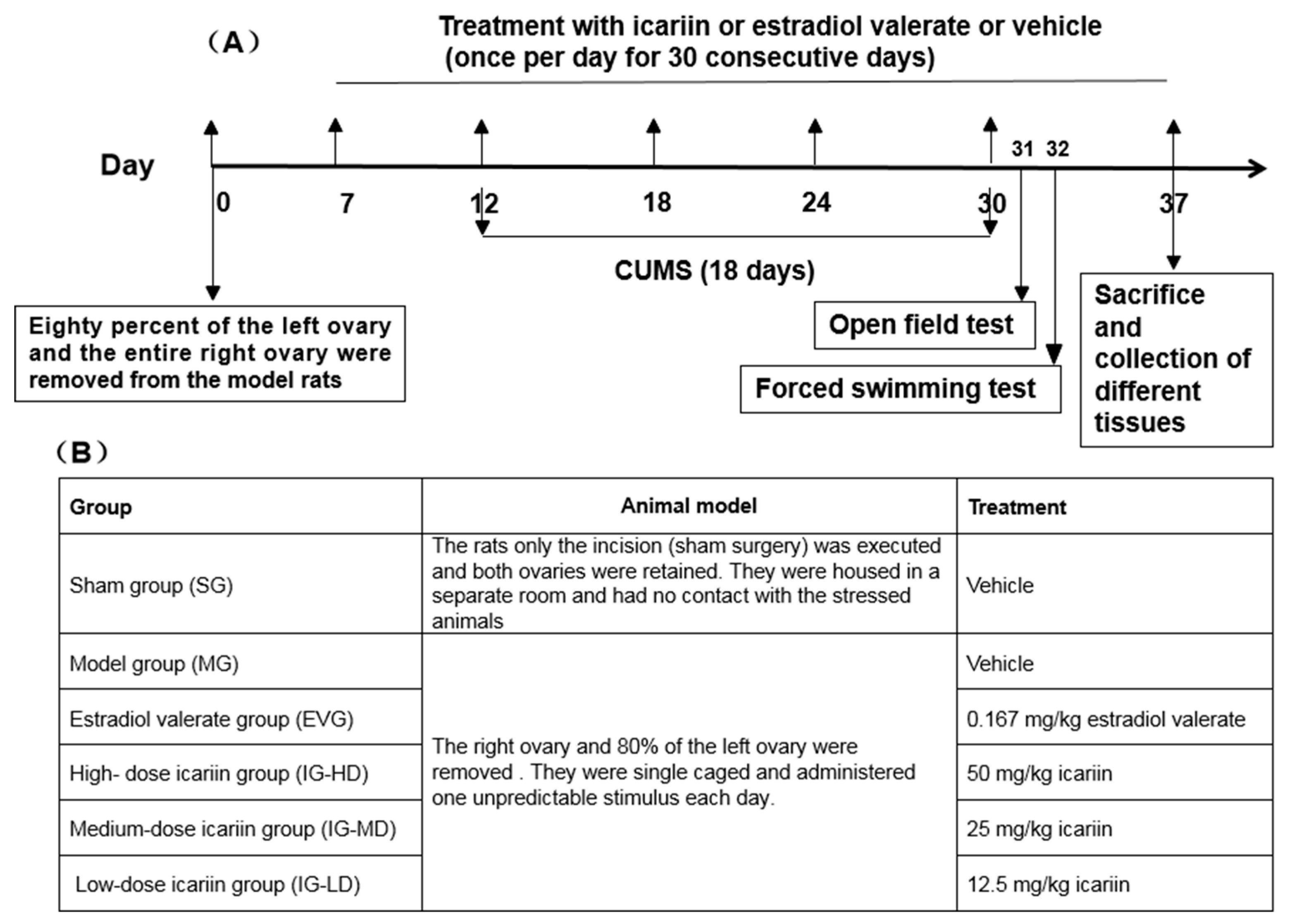
4.3. Experimental Design
4.4. Surgery
4.5. CUMS
4.6. Behavioral Assessments
4.7. Western Blot Analysis
4.8. Histopathological Examination and Immunofluorescence
4.9. Enzyme-Linked Immunosorbent Assay
4.10. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Natari, R.B.; Clavarino, A.M.; McGuire, T.M.; Dingle, K.D.; Hollingworth, S.A. The Bidirectional Relationship between Vasomotor Symptoms and Depression Across the Menopausal Transition: A Systematic Review of Longitudinal Studies. Menopause 2018, 25, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Maki, P.M.; Kornstein, S.G.; Joffe, H.; Bromberger, J.T.; Freeman, E.W.; Athappilly, G.; Bobo, W.V.; Rubin, L.H.; Koleva, H.K.; Cohen, L.S.; et al. Guidelines for the Evaluation and Treatment of Perimenopausal Depression: Summary and Recommendations. Menopause 2018, 25, 1069–1085. [Google Scholar] [CrossRef] [PubMed]
- Vivian-Taylor, J.; Hickey, M. Menopause and depression: Is there a link. Maturitas 2014, 79, 14–146. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.A.; Lovegreen, L.D. The Empty Nest Syndrome in Midlife Families: A Multimethod Exploration of Parental Gender Differences and Cultural Dynamics. J. Fam. Issues 2009, 30, 1651–1670. [Google Scholar] [CrossRef]
- Schmidt, P.J.; Murphy, J.H.; Haq, N.; Rubinow, D.R.; Danaceau, M.A. Stressful Life Events, Personal Losses, and Perimenopause-Related Depression. Arch. Womens Ment. Health 2004, 7, 19–26. [Google Scholar] [CrossRef]
- Strauss, J.R. The Reciprocal Relationship between Menopausal Symptoms and Depressive Symptoms: A 9-Year Longitudinal Study of American Women in Midlife. Maturitas 2011, 70, 302–306. [Google Scholar] [CrossRef]
- Dennerstein, L.; Guthrie, J.R.; Clark, M.; Lehert, P.; Henderson, V.W. A Population-Based Study of Depressed Mood in Middle-Aged, Australian-Born Women. Menopause 2004, 11, 563–568. [Google Scholar] [CrossRef]
- Callegari, C.; Buttarelli, M.; Cromi, A.; Diurni, M.; Salvaggio, F.; Bolis, P.F. Female Psychopathologic Profile during Menopausal Transition: A Preliminary Study. Maturitas 2007, 56, 447–451. [Google Scholar] [CrossRef]
- Gordon, J.L.; Peltier, A.; Grummisch, J.A.; Sykes Tottenham, L. Estradiol Fluctuation, Sensitivity to Stress, and Depressive Symptoms in the Menopause Transition: A Pilot Study. Front. Psychol. 2019, 10, 1319. [Google Scholar] [CrossRef]
- Albert, K.; Pruessner, J.; Newhouse, P. Estradiol Levels Modulate Brain Activity and Negative Responses to Psychosocial Stress Across the Menstrual Cycle. Psychoneuroendocrinology 2015, 59, 14–24. [Google Scholar] [CrossRef]
- Morgan, M.L.; Cook, I.A.; Rapkin, A.J.; Leuchter, A.F. Estrogen Augmentation of Antidepressants in Perimenopausal Depression: A Pilot Study. J. Clin. Psychiatry 2005, 66, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.L.; Cook, I.A.; Rapkin, A.J.; Leuchter, A.F. Neurophysiologic Changes during Estrogen Augmentation in Perimenopausal Depression. Maturitas 2007, 56, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Garay, R.P.; Zarate, C.A., Jr.; Charpeaud, T.; Citrome, L.; Correll, C.U.; Hameg, A.; Llorca, P.M. Investigational Drugs in Recent Clinical Trials for Treatment-Resistant Depression. Expert Rev. Neurother. 2017, 17, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Hugon-Rodin, J.; Amand, G.; Plu-Bureau, G. Safety of Estradiol Treatment in Perimenopausal Asymptomatic Women. JAMA Psychiatry 2018, 75, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Van Die, M.D.; Bone, K.M.; Burger, H.G.; Reece, J.E.; Teede, H.J. Effects of a Combination of Hypericum Perforatum and Vitex Agnus-Castus on PMS-Like Symptoms in Late-Perimenopausal Women: Findings from a Subpopulation Analysis. J. Altern. Complement. Med. 2009, 15, 1045–1048. [Google Scholar] [CrossRef]
- Cao, X.J.; Huang, X.C.; Wang, X. Effectiveness of Chinese Herbal Medicine Granules and Traditional Chinese Medicine-Based Psychotherapy for Perimenopausal Depression in Chinese Women: A Randomized Controlled Trial. Menopause 2019, 26, 1193–1203. [Google Scholar] [CrossRef]
- Tesch, B.J. Herbs Commonly Used by Women: An Evidence-Based Review. Am. J. Obstet. Gynecol. 2003, 188, S44–S55. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, D.; Yang, D.; Zhen, W.; Zhang, J.; Peng, S. The Effect of Icariin on Bone Metabolism and its Potential Clinical Application. Osteoporos. Int. 2018, 29, 535–544. [Google Scholar] [CrossRef]
- Xiao, H.H.; Fung, C.Y.; Mok, S.K.; Wong, K.C.; Ho, M.X.; Wang, X.L.; Yao, X.S.; Wong, M.S. Flavonoids from Herba Epimedii Selectively Activate Estrogen Receptor Alpha (ERA) and Stimulate ER-Dependent Osteoblastic Functions in UMR-106 Cells. J. Steroid Biochem. Mol. Biol. 2014, 143, 141–151. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, F.M.; Qiang, L.Q.; Zhang, D.M.; Kong, L.D. Icariin Attenuates Chronic Mild Stress-Induced Dysregulation of the LHPA Stress Circuit in Rats. Psychoneuroendocrinology 2010, 35, 72–83. [Google Scholar] [CrossRef]
- Gatza, M.L.; Lucas, J.E.; Barry, W.T.; Kim, J.W.; Wang, Q.; Crawford, M.D.; Datto, M.B.; Kelley, M.; Mathey-Prevot, B.; Potti, A.; et al. A Pathway-Based Classification of Human Breast Cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 6994–6999. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Loh, K.; Yap, Y.S. PI3K/Akt/mTOR Inhibitors in Breast Cancer. Cancer Biol. Med. 2015, 12, 342–354. [Google Scholar] [PubMed]
- Guo, X.Y. Study on PI3K/AKT-ENOS Mediated Icariin-Promoted Osteogenic Differentiation of Bone Marrow Stromal Cells and Osteoblasts. Gansu Agric. Univ. Beijing 2013, 84, 931–939. [Google Scholar]
- Ho, M.X.; Poon, C.C.; Wong, K.C.; Qiu, Z.C.; Wong, M.S. Icariin, But Not Genistein, Exerts Osteogenic and Anti-Apoptotic Effects in Osteoblastic Cells by Selective Activation of Non-Genomic ERα Signaling. Front. Pharm. 2018, 9, 474. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.J.; Han, B.; Wang, S.M.; Liang, S.W.; Zou, Z.J. Icariin Reverses Corticosterone-Induced Depression-Like Behavior, Decrease in Hippocampal Brain-Derived Neurotrophic Factor (BDNF) and Metabolic Network Disturbances Revealed by NMR-Based Metabonomics in Rats. J. Pharm. Biomed. Anal. 2016, 123, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Zweifel, J.E.; Obrien, W.H. A Meta-Analysis of the Effect of Hormone Replacement Therapy upon Depressed Mood. Psychoneuroendocrinology 1997, 22, 189–212. [Google Scholar] [CrossRef]
- Rubinow, D.R.; Johnson, S.L.; Schmidt, P.J.; Girdler, S.; Gaynes, B. Efficacy of Estradiol in Perimenopausal Depression: So Much Promise and so Few Answers. Depress. Anxiety 2015, 32, 539–549. [Google Scholar] [CrossRef]
- Schmidt, P.J.; Nieman, L.; Danaceau, M.A.; Tobin, M.B.; Roca, C.A.; Murphy, J.H.; Rubinow, D.R. Estrogen Replacement in Perimenopause-Related Depression: A Preliminary Report. Am. J. Obstet. Gynecol. 2000, 183, 414–420. [Google Scholar] [CrossRef]
- Whedon, J.M.; Kizhakke Veettil, A.; Rugo, N.A.; Kieffer, K.A. Bioidentical Estrogen for Menopausal Depressive Symptoms: A Systematic Review and Meta-Analysis. J. Womens Health (Larchmt) 2017, 26, 18–28. [Google Scholar] [CrossRef]
- Gordon, J.L.; Girdler, S.S. Hormone Replacement Therapy in the Treatment of Perimenopausal Depression. Curr. Psychiatry Rep. 2014, 16, 517. [Google Scholar] [CrossRef]
- Gordon, J.L.; Rubinow, D.R.; Eisenlohr-Moul, T.A.; Xia, K.; Schmidt, P.J.; Girdler, S.S. Efficacy of Transdermal Estradiol and Micronized Progesterone in the Prevention of Depressive Symptoms in the Menopause Transition: A Randomized Clinical Trial. JAMA Psychiatry 2018, 75, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.J.; Ben Dor, R.; Martinez, P.E.; Guerrieri, G.M.; Harsh, V.L.; Thompson, K.; Koziol, D.E.; Nieman, L.K.; Rubinow, D.R. Effects of Estradiol Withdrawal on Mood in Women with Past Perimenopausal Depression: A Randomized Clinical Trial. JAMA Psychiatry 2015, 72, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Marsh, W.K.; Bromberger, J.T.; Crawford, S.L.; Leung, K.; Kravitz, H.M.; Randolph, J.F.; Joffe, H.; Soares, C.N. Lifelong Estradiol Exposure and Risk of Depressive Symptoms during the Transition to Menopause and Postmenopause. Menopause 2017, 24, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, J.E.; Anderson, G.L.; Prentice, R.L.; LaCroix, A.Z.; Kooperberg, C.; Stefanick, M.L.; Jackson, R.D.; Beresford, S.A.; Howard, B.V.; Johnson, K.C.; et al. Risks and Benefits of Estrogen Plus Progestin in Healthy Postmenopausal Women: Principalresults from the Women’s Health Initiative Randomized Controlled Trial. JAMA 2002, 288, 321–333. [Google Scholar] [PubMed]
- Million Women Study Collaborators. Ovarian Cancer and Hormone Replacement Therapy in the Million Women Study. Lancet 2007, 369, 1703–1710. [Google Scholar] [CrossRef]
- Hill, D.A.; Crider, M.; Hill, S.R. Hormone Therapy and other Treatments for Symptoms of Menopause. Am. Fam. Physician 2016, 94, 884–889. [Google Scholar]
- Sarri, G.; Pedder, H.; Dias, S.; Guo, Y.; Lumsden, M.A. Vasomotor Symptoms Resulting from Natural Menopause: A Systematic Review and Network Meta-Analysis of Treatment Effects from the National Institute for Health and Care Excellence Guideline on Menopause. BJOG 2017, 124, 1514–1523. [Google Scholar] [CrossRef]
- Cao, L.H.; Miao, M.S. Analysis of Animal Models Based on Clinical Features of Perimenopausal Depression. China J. Tradit. Chin. Med. Pharm. 2016, 31, 1643–1647. [Google Scholar]
- Calmarza-Font, I.; Lagunas, N.; Garcia-Segura, L.M. Antidepressive and Anxiolytic Activity of Selective Estrogen Receptor Modulators in Ovariectomized Mice Submitted to Chronic Unpredictable Stress. Behav. Brain Res. 2012, 227, 287–290. [Google Scholar] [CrossRef]
- Koshkina, A.; Dudnichenko, T.; Baranenko, D.; Fedotova, J.; Drago, F. Effects of Vitamin D3 in Long-Term Ovariectomized Rats Subjected to Chronic Unpredictable Mild Stress: BDNF, NT-3, and NT-4 Implications. Nutrients 2019, 11, 1726. [Google Scholar] [CrossRef]
- Zhou, X.D.; Shi, D.D.; Zhang, Z.J. Ameliorative Effects of Radix Rehmanniae Extract on the Anxiety-And Depression-Like Symptoms in Ovariectomized Mice: A Behavioral and Molecular Study. Phytomedicine 2019, 63, 153012. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Chen, Y.X.; Dong, M. Treat Perimenopausal Sleep Disorders Based on Kidney Deficiency Blood Stasis and Heart-Kidney Imbalance. China J. Chin. Mater. Med. 2017, 42, 4455–4458. [Google Scholar]
- Vichinsartvichai, P.; Sirirat, S. Hematologic Parameters as the Predictors for Metabolic Syndrome in Perimenopausal and Postmenopausal Women Living in Urban Area: A Preliminary Report. Menopause. Prz. 2016, 15, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, J.; Yu, Y.; Yang, P.; Li, H.; Shen, Y.; Huang, X.; Liu, S. Evaluation of Basal Sex Hormone Levels for Activation of the Hypothalamic-Pituitary-Gonadal Axis. J. Pediatr. Endocrinol. Metab. 2018, 31, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Chen, D.R. Relationship between TCM syndrome differentiation in menopausal women and estradiol, follicle stimulating hormone and luteinizing hormone. China J. Tradit Chin. Med. Pharm. 2008, 23, 171–173. [Google Scholar]
- Wang, H. Clinical Syndrome Differentiation of Menopausal Syndrome. Beijing Univ. Chin. Med. 2003, 21, 93–95. [Google Scholar]
- Akyol, S.; Cinar, S.A.; Purisa, S.; Aydinli, K. Relationship between Lymphocytes, IL2 and the Hormones E2, LH, PRG and FSH in Menopausal and Postmenopausal Women. Am. J. Reprod Immunol. 2011, 66, 304–309. [Google Scholar] [CrossRef]
- Zhai, Z.M. Biregulation Effects of IL-2 on Immune Activation and Immune Tolerance. Chin. Pharmacol. Bull. 2013, 29, 319–322. [Google Scholar]
- Wang, Y.; Si, L.F.; Li, X.N.; Li, Z. Ginkgo Biloba Extract Enhances the Immune Function of Spleen and Thymus in Rats. Chin. J. Cell Mol. Immunol. 2015, 31, 792–5799. [Google Scholar]
- Han, P.; Song, Z.M. Effects of Modified Jieganjian on Neurotransmitter 5-HT and Blood Lipid Levels in Menopausal Depression Rats. Clin. J. Tradit. Chin. Med. 2014, 26, 407–409. [Google Scholar]
- Wang, H.B.; Li, X.H.; He, Y.W.; Xu, L.L.; Song, X.L.; Zheng, L. Effects of Reverse Acupuncture at Guanyuan Point on ACTH, NE and 5-HT Contents in Natural Menopausal Rats. J. BJ Univ. Chin. Med. 2008, 31, 826–829. [Google Scholar]
- Chen, B.; Li, J.; Xie, Y.; Ming, X.; Li, G.; Wang, J.; Li, M.; Li, X.; Xiong, L. Cang-Ai Volatile Oil Improves Depressive-Like Behaviors and Regulates DA and 5-HT Metabolism in the Brains of CUMS-Induced Rats. J. Ethnopharmacol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, Y.; Tsutsui-Kimura, I.; Sasamori, H.; Nebuka, M.; Nishitani, N.; Tanaka, K.F.; Yamanaka, A.; Yoshioka, M. Different Roles of Distinct Serotonergic Pathways in Anxiety-Like Behavior, Antidepressant-Like, and Anti-Impulsive Effects. Neuropharmacology 2019, 9, 107703. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Chen, S.J.; Liang, W.N.; Wang, M.; Li, C.F.; Wang, S.S.; Dong, S.Q.; Yi, L.T.; Li, C.D. Saikosaponin a Attenuates Perimenopausal Depression-Like Symptoms by Chronic Unpredictable Mild Stress. Neurosci. Lett. 2018, 662, 283–289. [Google Scholar] [CrossRef]
- Claudia Maria, P. Serotonin in Anxiety and Depression. J. Psychopharmacol. 2013, 27, 1083. [Google Scholar]
- Elzaiat, M.; Herman, L.; Legois, B.; Leger, T.; Todeschini, A.L.; Veitia, R.A. High-Throughput Exploration of the Network Dependent on AKT1 in Mouse Ovarian Granulosa Cells. Mol. Cell Proteomics. 2019, 8, 1307–1319. [Google Scholar] [CrossRef]
- Zhang, H.; Qin, F.; Liu, A.; Sun, Q.; Wang, Q.; Xie, S.; Lu, S.; Zhang, D.; Lu, Z. Electro-Acupuncture Attenuates the Mice Premature Ovarian Failure Via Mediating PI3K/AKT/mTOR Pathway. Life Sci. 2019, 217, 169–175. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Yu, B.; Chen, J.; Chen, J.; Zhao, Z.S.; Wang, J.L. Effects of Acupuncture on PI3K/Akt/mTOR Pathway in Rats with Premature Ovarian Failure. Chin. Acupunct. Moxibust 2015, 35, 53–58. [Google Scholar]
- Jiang, Y.F.; Wang, S.E. Estrogen Receptor ERα and PI3K/Akt Cell Signaling Pathways. Anat. Res. 2012, 34, 142–145. [Google Scholar]
- Zhang, F.; Feng, F.; Yang, P.; Li, Z.; You, J.; Xie, W.; Gao, X.; Yang, J. Four-And-A-Half-LIM Protein 1 Down-Regulates Estrogen Receptor α Activity Through Repression of AKT Phosphorylation in Human Breast Cancer Cell. Int. J. Biochem. Cell Biol. 2012, 44, 320–326. [Google Scholar] [CrossRef]
- Sun, M.; Paciga, J.E.; Feldman, R.I.; Yuan, Z.; Coppola, D.; Lu, Y.Y.; Shelley, S.A.; Nicosia, S.V.; Cheng, J.Q. Phosphatidylinositol-3-OH Kinase (PI3K)/AKT2, Activated in Breast Cancer, Regulates and is Induced by Estrogen Receptor Alpha (ERalpha) Via Interaction between ERalpha and PI3K. Cancer Res. 2001, 61, 5985–5991. [Google Scholar] [PubMed]
- Santen, R.J.; Song, R.X.; Zhang, Z.; Kumar, R.; Jeng, M.H.; Masamura, A.; Lawrence, J., Jr.; Berstein, L.; Yue, W. Long-Term Estradiol Deprivation in Breast Cancer Cells Up-Regulates Growth Factor Signaling and Enhances Estrogen Sensitivity. Endocr. Relat. Cancer 2005, 12, S61–S73. [Google Scholar] [CrossRef] [PubMed]
- Bratton, M.R.; Duong, B.N.; Elliott, S.; Weldon, C.B.; Beckman, B.S.; McLachlan, J.A.; Burow, M.E. Regulation of ER Alpha-Mediated Transcription of Bcl-2 by PI3K-AKT Crosstalk: Implications for Breast Cancer Cell Survival. Int. J. Oncol. 2010, 37, 541–550. [Google Scholar] [PubMed]
- Li, Z.C.; Yu, H.Y.; Wang, X.X.; Zhang, M.; Wang, J.P. Olmesartan Medoxomil Reverses Left Ventricle Hypertrophy and Reduces Inflammatory Cytokine IL-6 in the Renovascular Hypertensive Rats. Eur. Rev. Med. Pharm. Sci. 2013, 17, 3318–3322. [Google Scholar]
- Tian, S.; Miao, M.S.; Li, X.M.; Bai, M.; Wu, Y.Y.; Wei, Z.Z. Study on Neuroendocrine-Immune Function of Phenylethanoid Glycosides of Desertliving Cistanche Herb in Perimenopausal Rat Model. J. Ethnopharmacol. 2019, 238, 111884. [Google Scholar] [CrossRef]
- Fahim, A.T.; Abd El-Fattah, A.A.; Sadik NAH Ali, B.M. Resveratrol and Dimethyl Fumarate Ameliorate Testicular Dysfunction Caused by Chronic Unpredictable Mild Stress-Induced Depression in Rats. Arch. Biochem. Biophys. 2019, 665, 152–165. [Google Scholar] [CrossRef]
- Saad, M.A.; El-Sahar, A.E.; Sayed, R.H.; Elbaz, E.M.; Helmy, H.S.; Senousy, M.A. Venlafaxine Mitigates Depressive-Like Behavior in Ovariectomized Rats by Activating the EPO/EPOR/JAK2 Signaling Pathway and Increasing the Serum Estradiol Level. Neurotherapeutics 2019, 16, 404–415. [Google Scholar] [CrossRef]
- Chen, X.Q.; Li, C.F.; Chen, S.J.; Liang, W.N.; Wang, M.; Wang, S.S.; Dong, S.Q.; Yi, L.T.; Li, C.D. The Antidepressant-Like Effects of Chaihu Shugan San: Dependent on the Hippocampal BDNF-TrkB-ERK/Akt Signaling Activation in Perimenopausal Depression-Like Rats. Biomed. Pharmacother. 2018, 105, 45–52. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stressor | Details |
|---|---|
| Food deprivation | Rats were subjected to 24 h of food deprivation. Food was provided immediately after the end of the fasting period. |
| Water deprivation | Rats were subjected to 24 h of water deprivation. Water was provided immediately after the end of the fasting period. |
| Overnight illumination | Lights on for 24 h starting 20:30 |
| Wet sawdust bedding | 1 mL of water per 1 g of sawdust bedding for 20 h. Immediately after the stress, rats were removed from the cage and towel dried before being placed back in their home cage. |
| Heat environment | 45 °C for 5 min |
| Cold water swimming | Rats were placed for 5 min in a cylindrical clear plastic tank (46 cm high (H) × 20 cm diameter (D) filled with water (4 ± 1 °C) to a depth of 30 cm. Immediately after the swim, rats were removed from the tank and towel dried before being placed back in their home cage. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.-H.; Qiao, J.-Y.; Huang, H.-Y.; Fang, X.-Y.; Zhang, R.; Miao, M.-S.; Li, X.-M. PI3K–AKT Signaling Activation and Icariin: The Potential Effects on the Perimenopausal Depression-Like Rat Model. Molecules 2019, 24, 3700. https://doi.org/10.3390/molecules24203700
Cao L-H, Qiao J-Y, Huang H-Y, Fang X-Y, Zhang R, Miao M-S, Li X-M. PI3K–AKT Signaling Activation and Icariin: The Potential Effects on the Perimenopausal Depression-Like Rat Model. Molecules. 2019; 24(20):3700. https://doi.org/10.3390/molecules24203700
Chicago/Turabian StyleCao, Li-Hua, Jing-Yi Qiao, Hui-Yuan Huang, Xiao-Yan Fang, Rui Zhang, Ming-San Miao, and Xiu-Min Li. 2019. "PI3K–AKT Signaling Activation and Icariin: The Potential Effects on the Perimenopausal Depression-Like Rat Model" Molecules 24, no. 20: 3700. https://doi.org/10.3390/molecules24203700
APA StyleCao, L.-H., Qiao, J.-Y., Huang, H.-Y., Fang, X.-Y., Zhang, R., Miao, M.-S., & Li, X.-M. (2019). PI3K–AKT Signaling Activation and Icariin: The Potential Effects on the Perimenopausal Depression-Like Rat Model. Molecules, 24(20), 3700. https://doi.org/10.3390/molecules24203700