Widely Targeted Metabolomics Analysis Reveals the Effect of Flooding Stress on the Synthesis of Flavonoids in Chrysanthemum morifolium

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
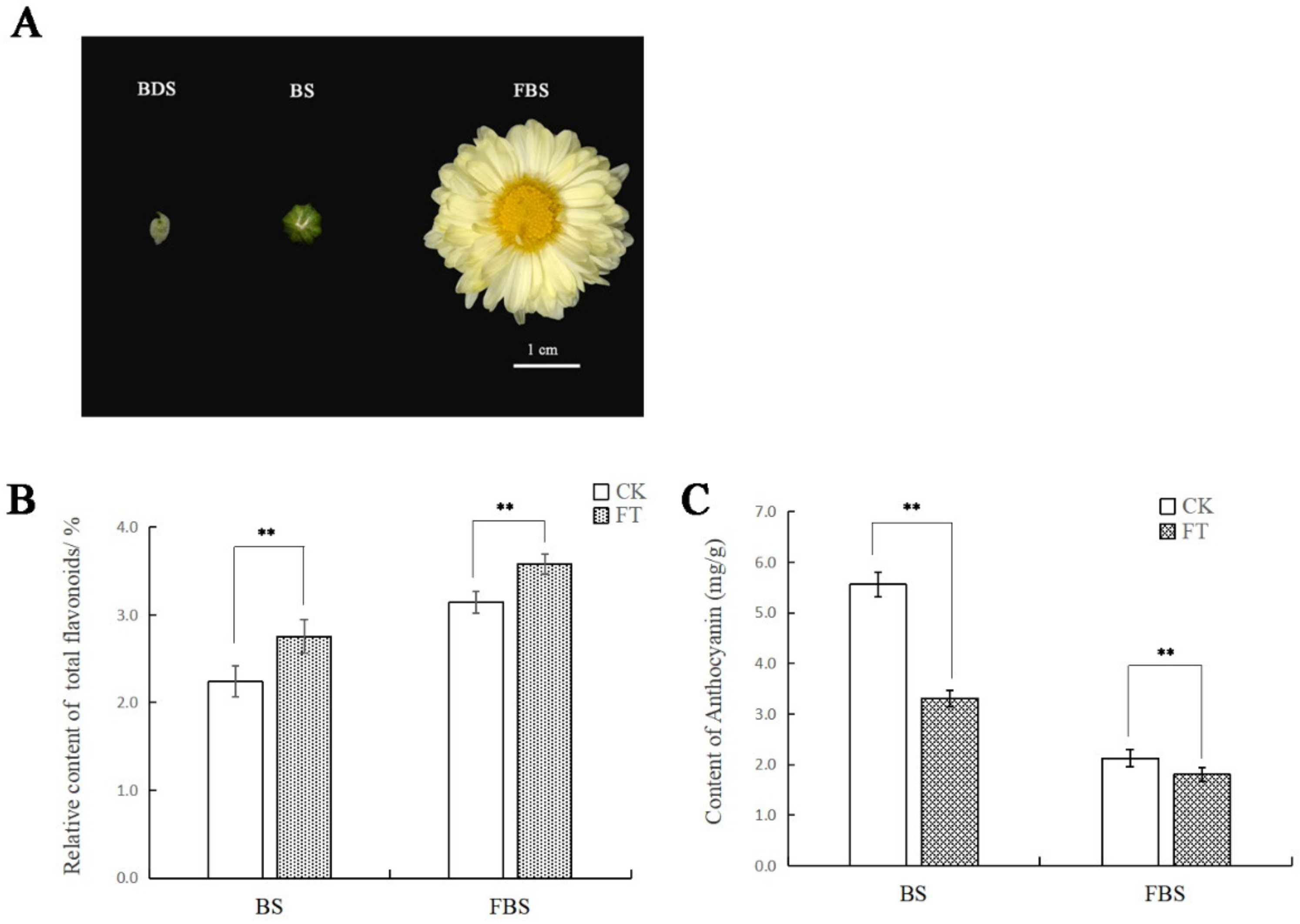
2.1. Differences in Total Flavonoids and Anthocyanins Contents in “Hangju” After Flooding Stress
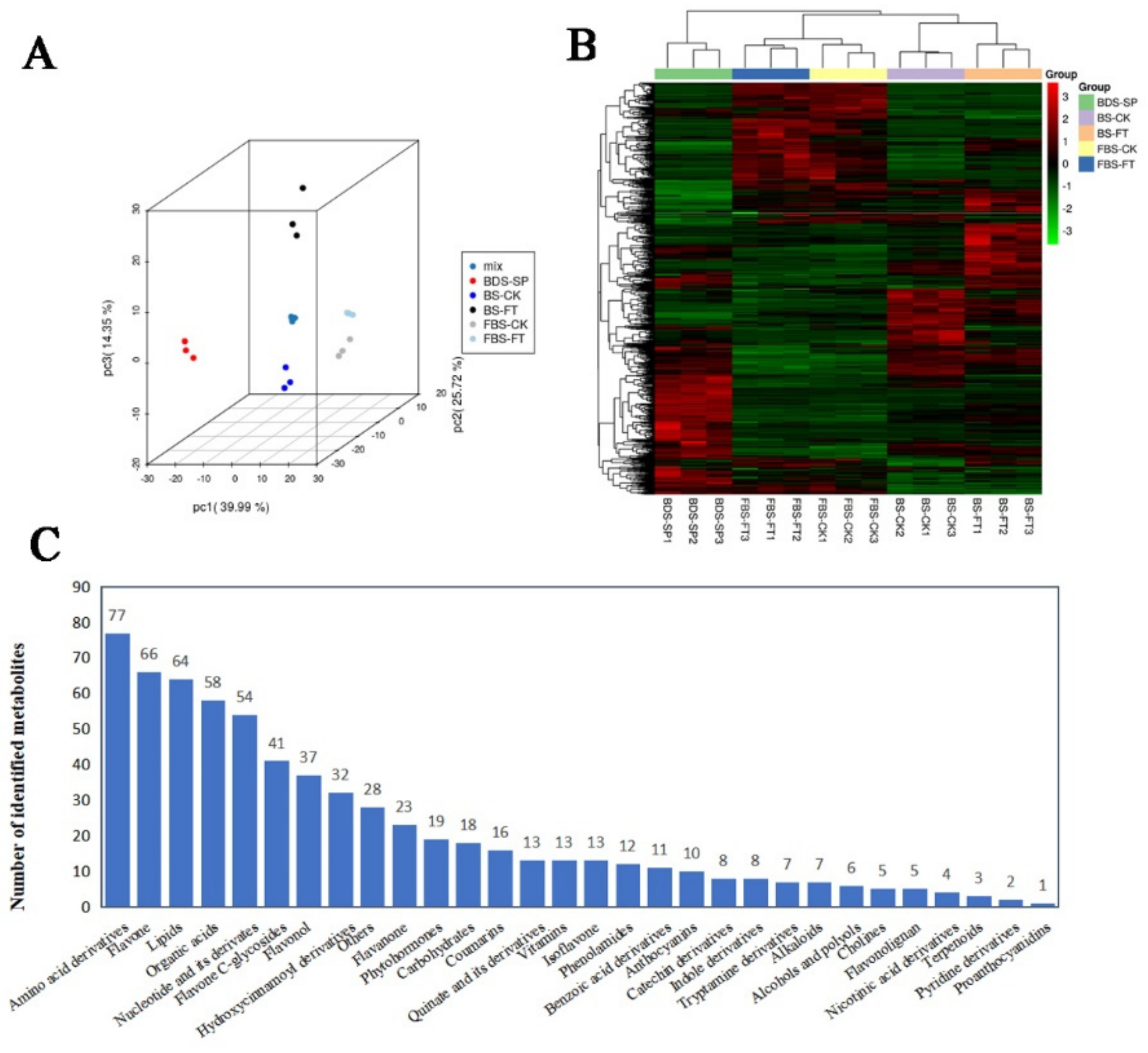
2.2. Metabolite Profiling of “Hangju” in Different Growth Stages After Flooding Stress Treatment
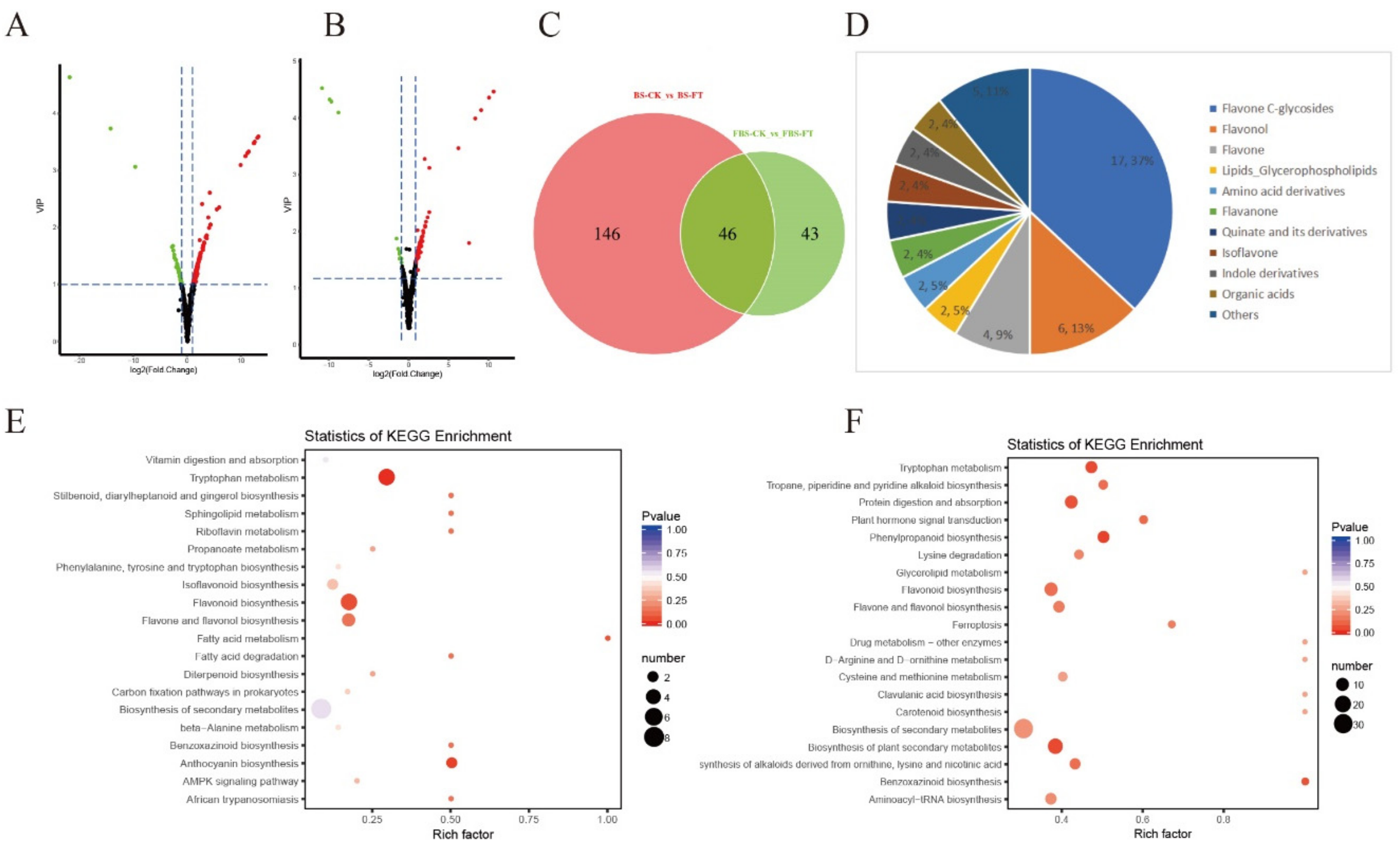
2.3. Analysis of Differential Metabolites in “Hangju” After Flooding Stress
2.4. Analysis of Differential Metabolites in “Hangju” After Flooding Stress
3. Discussion
3.1. Metabolome Analysis to Evaluate the Quality of Medicinal Plants
3.2. Relationship between Flooding Stress and Medicinal Quality of “Hangju”
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. Flooding Treatment and Sampling Method
4.3. Measurement of Total Flavonoids
4.4. Measurement of Anthocyanins
4.5. Analysis of “Hangju” Metabolomics Based on LC-MS Data
4.6. Real-Time Quantitative PCR
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Da Silva, J.A.T. Chrysanthemum: Advances in tissue culture, cryopreservation, postharvest technology, genetics and transgenic biotechnology. Biotechnol. Adv. 2003, 21, 715–766. [Google Scholar] [CrossRef]
- Van Der Ploeg, A.; Heuvelink, E. The influence of temperature on growth and development of chrysanthemum cultivars. J. Hortic. Sci. Biotechnol. 2015, 81, 174–182. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M. Identification of the phenolic components of chrysanthemum flower (Chrysanthemum morifolium Ramat). Food Chem. 2010, 120, 319–326. [Google Scholar] [CrossRef]
- Miao, J. Growth and accumulation of bioactive compounds in medicinal Chrysanthemum morifolium Ramat. cv. ’Chuju’ under different colored shade polyethylene. J. Med. Plants Res. 2012, 6. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Editorial Committee. Pharmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2015; pp. 310–311. [Google Scholar]
- Ukiya, M.; Akihisa, T.; Tokuda, H.; Suzuki, H.; Mukainaka, T.; Ichiishi, E.; Yasukawa, K.; Kasahara, Y.; Nishino, H. Constituents of Compositae plants III. Anti-tumor promoting effects and cytotoxic activity against human cancer cell lines of triterpene diols and triols from edible chrysanthemum flowers. Cancer Lett. 2002, 177, 7–12. [Google Scholar] [CrossRef]
- Kim, I.S.; Koppula, S.; Park, P.-J.; Kim, E.H.; Kim, C.G.; Choi, W.S.; Lee, K.H.; Choi, D.-K. Chrysanthemum morifolium Ramat (CM) extract protects human neuroblastoma SH-SY5Y cells against MPP+-induced cytotoxicity. J. Ethnopharmacol. 2009, 126, 447–454. [Google Scholar] [CrossRef]
- Tsuji-Naito, K.; Saeki, H.; Hamano, M. Inhibitory effects of Chrysanthemum species extracts on formation of advanced glycation end products. Food Chem. 2009, 116, 854–859. [Google Scholar] [CrossRef]
- Lii, C.K.; Lei, Y.P.; Yao, H.T.; Hsieh, Y.S.; Tsai, C.W.; Liu, K.L.; Chen, H.W. Chrysanthemum morifolium Ramat. reduces the oxidized LDL-induced expression of intercellular adhesion molecule-1 and E-selectin in human umbilical vein endothelial cells. J. Ethnopharmacol. 2010, 128, 213–220. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, H.J.; Lee, Y.S. A new anti-HIV flavonoid from glucoronide from Chrysanthemum marifolium. Planta Med. 2003, 69, 859–861. [Google Scholar]
- Shao, Q.-S.; Guo, Q.-S.; Deng, Y.-M.; Guo, H.-P. A comparative analysis of genetic diversity in medicinal Chrysanthemum morifolium based on morphology, ISSR and SRAP markers. Biochem. Syst. Ecol. 2010, 38, 1160–1169. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, J.; He, L.; Sun, Y.; Cai, H. Molecular structure and the second introns variation of gene F3 ’ H of two medicinal Chrysanthemum morifolium populations. Biochem. Syst. Ecol. 2013, 51, 251–258. [Google Scholar] [CrossRef]
- Huang, L.; Guo, L.; Ma, C.; Gao, W.; Yuan, Q. Top-geoherbs of traditional Chinese medicine: Common traits, quality characteristics and formation. Front. Med. 2011, 5, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhu, Z.; Guo, Q.; Mao, P. Variation in major flavonoids glycosides and caffeoylquinic acids during florescence of three Chrysanthemum morifolium Ramat cv. ’Hangju’ genotypes. Biochem. Syst. Ecol. 2013, 47, 74–79. [Google Scholar] [CrossRef]
- Ueyama, Y.; Suzuki, K.; Fukuchi-Mizutani, M.; Fukui, Y.; Miyazaki, K.; Ohkawa, H.; Kusumi, T.; Tanaka, Y. Molecular and biochemical characterization of torenia flavonoid 3′-hydroxylase and flavone synthase II and modification of flower color by modulating the expression of these genes. Plant. Sci. 2002, 163, 253–263. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, B.; Singh, K. Functional characterization of flavanone 3-hydroxylase gene from Phyllanthus emblica (L.). J. Plant. Biochem. Biotechnol. 2015, 24, 453–460. [Google Scholar] [CrossRef]
- Pawlikowska-Pawlega, B.; Gruszecki, W.I.; Misiak, L.; Paduch, R.; Piersiak, T.; Zarzyka, B.; Pawelec, J.; Gawron, A. Modification of membranes by quercetin, a naturally occurring flavonoid, via its incorporation in the polar head group. Biochim. Biophys. Acta. 2007, 1768, 2195–2204. [Google Scholar] [CrossRef]
- Netzel, M.; Strass, G.; Kaul, C.; Bitsch, I.; Dietrich, H.; Bitsch, R. In vivo antioxidative capacity of a composite berry juice. Food Res. Int. 2002, 35, 213–216. [Google Scholar] [CrossRef]
- Ashida, H.; Fukuda, I.; Yamashita, T.; Kanazawa, K. Flavones and flavonols at dietary levels inhibit a transformation of aryl hydrocarbon receptor induced by dioxin. Febs. Lett. 2000, 476, 213–217. [Google Scholar] [CrossRef]
- Deavours, B.E.; Dixon, R.A. Metabolic engineering of isoflavonoid biosynthesis in alfalfa. Plant. Physiol. 2005, 138, 2245–2259. [Google Scholar] [CrossRef]
- Huang, L.-Q.; Guo, L.-P. Secondary metabolites accumulating and geoherbs formation under enviromental stress. China J. Chin. Mater. Med. 2007, 32, 277–280. [Google Scholar]
- Zou, Q.-J.; Wang, T.; Guo, Q.-S.; Xiao, Y.-M.; Wu, L.-W. Cloning and expression analysis of F3’H and quantification of downstream products in Chrysanthemum morifolium under flooding stress. China J. Chin. Mater. Med. 2018, 43, 52–57. [Google Scholar] [CrossRef]
- Jackson, M.B.; Colmer, T.D. Response and adaptation by plants to flooding stress. Ann. Bot 2005, 96, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant. 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-Discovery Approach for Sample Matching of a Nerve-Agent Precursor Using Liquid Chromatography-Mass Spectrometry, XCMS, and Chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Brennan, L. The nutritional metabolomics crossroads: How to ensure success for dietary biomarkers. Am. J. Clin. Nutr. 2017, 105, 293–294. [Google Scholar] [CrossRef]
- Mallick, S.; Singh, S.K.; Sarkar, C.; Saha, B.; Bhadra, R. Human placental lipid induces melanogenesis by increasing the expression of tyrosinase and its related proteins in vitro. Pigment. Cell Res. 2005, 18, 25–33. [Google Scholar] [CrossRef]
- Kanu, P.J.; Zhu, K.; Kanu, J.B.; Zhou, H.; Qian, H.; Zhu, K. Biologically active components and nutraceuticals in sesame and related products: A review and prospect. Trends Food Sci. Technol. 2007, 18, 599–608. [Google Scholar] [CrossRef]
- Bose, U.; Hewavitharana, A.K.; Ng, Y.K.; Shaw, P.N.; Fuerst, J.A.; Hodson, M.P. LC-MS-Based Metabolomics Study of Marine Bacterial Secondary Metabolite and Antibiotic Production in Salinispora arenicola. Mar. Drugs 2015, 13, 249–266. [Google Scholar] [CrossRef]
- Saito, K.; Matsuda, F. Metabolomics for Functional Genomics, Systems Biology, and Biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef]
- Duan, L.-X.; Chen, T.-L.; Li, M.; Chen, M.; Zhou, Y.-Q.; Cui, G.-H.; Zhao, A.-H.; Jia, W.; Huang, L.-Q.; Qi, X. Use of the Metabolomics Approach to Characterize Chinese Medicinal Material Huangqi. Mol. Plant. 2012, 5, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Kueger, S.; Steinhauser, D.; Willmitzer, L.; Giavalisco, P. High-resolution plant metabolomics: From mass spectral features to metabolites and from whole-cell analysis to subcellular metabolite distributions. Plant. J. 2012, 70, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, L.; Huang, X.; Wang, X.; Yang, R.; Mao, J.; Wang, X.; Wang, X.; Zhang, Q.; Li, P. Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines. Molecules 2018, 23, 1180. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, C.; Zhou, C.; Li, J.; Zhang, Y. Molecular characterization of the C-glucosylation for puerarin biosynthesis in Pueraria lobata. Plant. J. 2017, 90, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Wang, B.; He, G.; Wang, Y.; Tang, X.; Wang, S.; Ma, Y.; Fu, C.; Chai, G.; Zhou, G. Metabolomics Integrated with Transcriptomics Reveals Redirection of the Phenylpropanoids Metabolic Flux in Ginkgo biloba. J. Agric. Food Chem 2019, 67, 3284–3291. [Google Scholar] [CrossRef]
- Mattson, M.P. Hormesis defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. Hormesis as a biological hypothesis. Environ. Health Perspect. 1998, 106, 357–362. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, X.; Yang, G.; Huang, L.; Ma, J. Hormesis and its application in medicinal plant growing. China J. Chin. Mater. Med. 2011, 36, 525–529. [Google Scholar]
- Liu, S.P.; Bi, Y.T.; Tian, W.; Xue, Y.H. Comparative Physiological Responses of Two Buxus Species on Flooding Stress. Appl. Mech. Mater. 2013, 295–298, 2314–2317. [Google Scholar] [CrossRef]
- Guo, L.-P.; Huang, L.-Q.; Yan, H.; Lv, D.-M.; Jiang, Y.-X. Habitat characteristics for the growth of Atractylodes lancea based on GIS. China J. Chin. Mater. Med. 2005, 30, 565–569. [Google Scholar]
- Cipollini, M.L.; Paulk, E.; Cipollini, D.F. Effect of nitrogen and water treatment on leaf chemistry in horsenettle (Solanum carolinense), and relationship to herbivory by flea beetles (Epitrix spp.) and tobacco hornworm (Manduca sexta). J. Chem. Ecol. 2002, 28, 2377–2398. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant. Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Halliwell, B.; Rafter, J.; Jenner, A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: Direct or indirect effects? Antioxidant or not? Am. J. Clin. Nutr. 2005, 81, 268S–276S. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, I.M.; Zhang, S.M.; Blumberg, J.B.; Buring, J.E.; Sesso, H.D. Dietary intake of selected flavonols, flavones, and flavonoid-rich foods and risk of cancer in middle-aged and older women. Am. J. Clin. Nutr. 2009, 89, 905–912. [Google Scholar] [CrossRef]
- Jan, A.T.; Kamli, M.R.; Murtaza, I.; Singh, J.B.; Ali, A.; Haq, Q.M.R. Dietary Flavonoid Quercetin and Associated Health BenefitsAn Overview. Food Rev. Int. 2010, 26, 302–317. [Google Scholar] [CrossRef]
- Wang, T.; Guo, Q.-s.; Mao, P.-F. Flavonoid accumulation during florescence in three Chrysanthemum morifolium Ramat cv. ’Hangju’ genotypes. Biochem. Syst. Ecol. 2014, 55, 79–83. [Google Scholar] [CrossRef]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant. Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef]
- Wang, Y.; Lysoe, E.; Armarego-Marriott, T.; Erban, A.; Paruch, L.; van Eerde, A.; Bock, R.; Liu-Clarke, J. Transcriptome and metabolome analyses provide insights into root and root-released organic anion responses to phosphorus deficiency in oat. J. Exp. Bot. 2018, 69, 3759–3771. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Zou, Q.; Guo, Q.; Yang, F.; Wu, L.; Zhang, W. Widely Targeted Metabolomics Analysis Reveals the Effect of Flooding Stress on the Synthesis of Flavonoids in Chrysanthemum morifolium. Molecules 2019, 24, 3695. https://doi.org/10.3390/molecules24203695
Wang T, Zou Q, Guo Q, Yang F, Wu L, Zhang W. Widely Targeted Metabolomics Analysis Reveals the Effect of Flooding Stress on the Synthesis of Flavonoids in Chrysanthemum morifolium. Molecules. 2019; 24(20):3695. https://doi.org/10.3390/molecules24203695
Chicago/Turabian StyleWang, Tao, Qingjun Zou, Qiaosheng Guo, Feng Yang, Liwei Wu, and Wenyan Zhang. 2019. "Widely Targeted Metabolomics Analysis Reveals the Effect of Flooding Stress on the Synthesis of Flavonoids in Chrysanthemum morifolium" Molecules 24, no. 20: 3695. https://doi.org/10.3390/molecules24203695
APA StyleWang, T., Zou, Q., Guo, Q., Yang, F., Wu, L., & Zhang, W. (2019). Widely Targeted Metabolomics Analysis Reveals the Effect of Flooding Stress on the Synthesis of Flavonoids in Chrysanthemum morifolium. Molecules, 24(20), 3695. https://doi.org/10.3390/molecules24203695