Co-Immobilization of Tri-Enzymes for the Conversion of Hydroxymethylfurfural to 2,5-Diformylfuran


Abstract
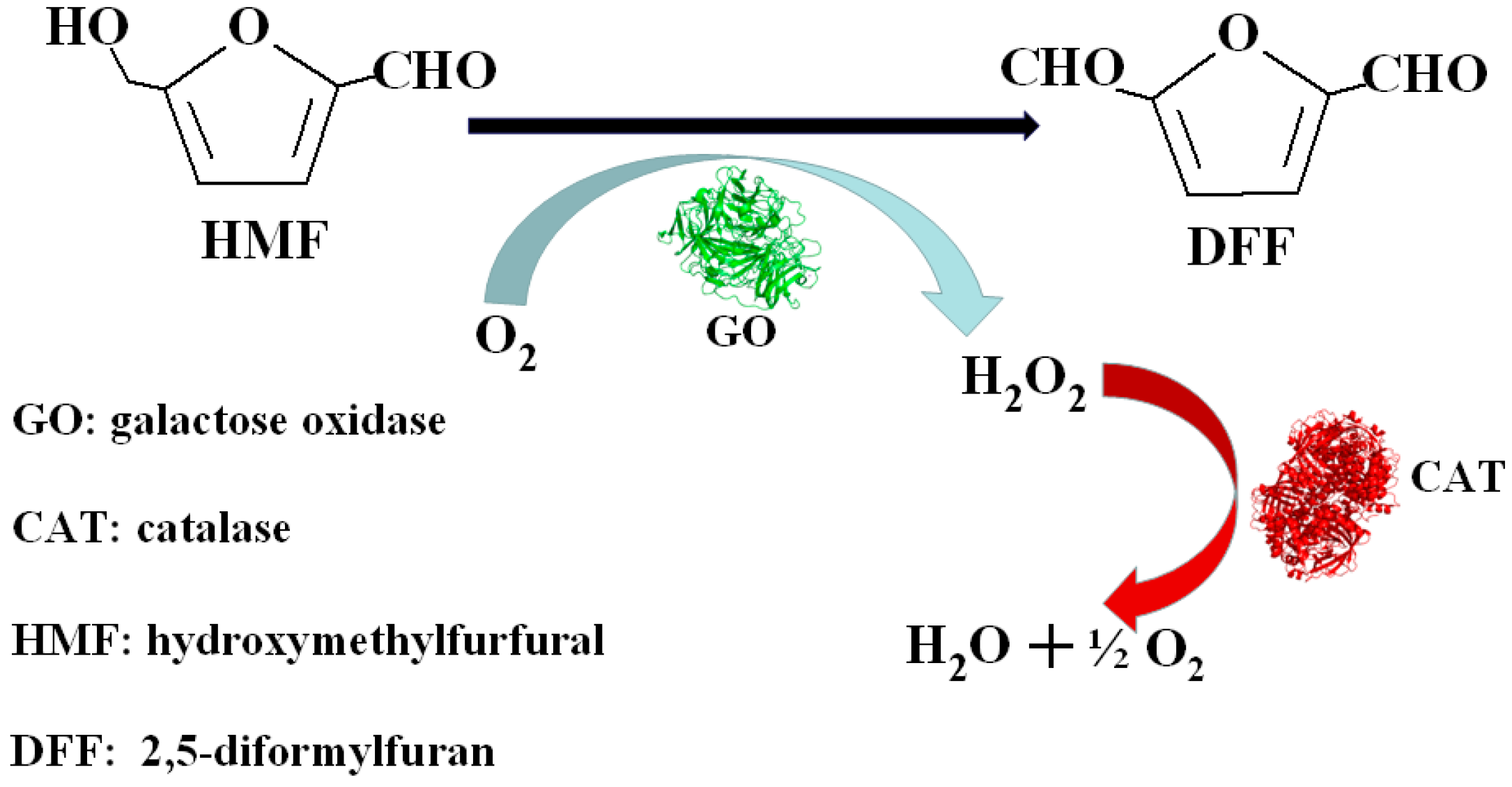
1. Introduction
2. Results and Discussion
2.1. SEM Images of Co-Immobilized Tri-Enzymes
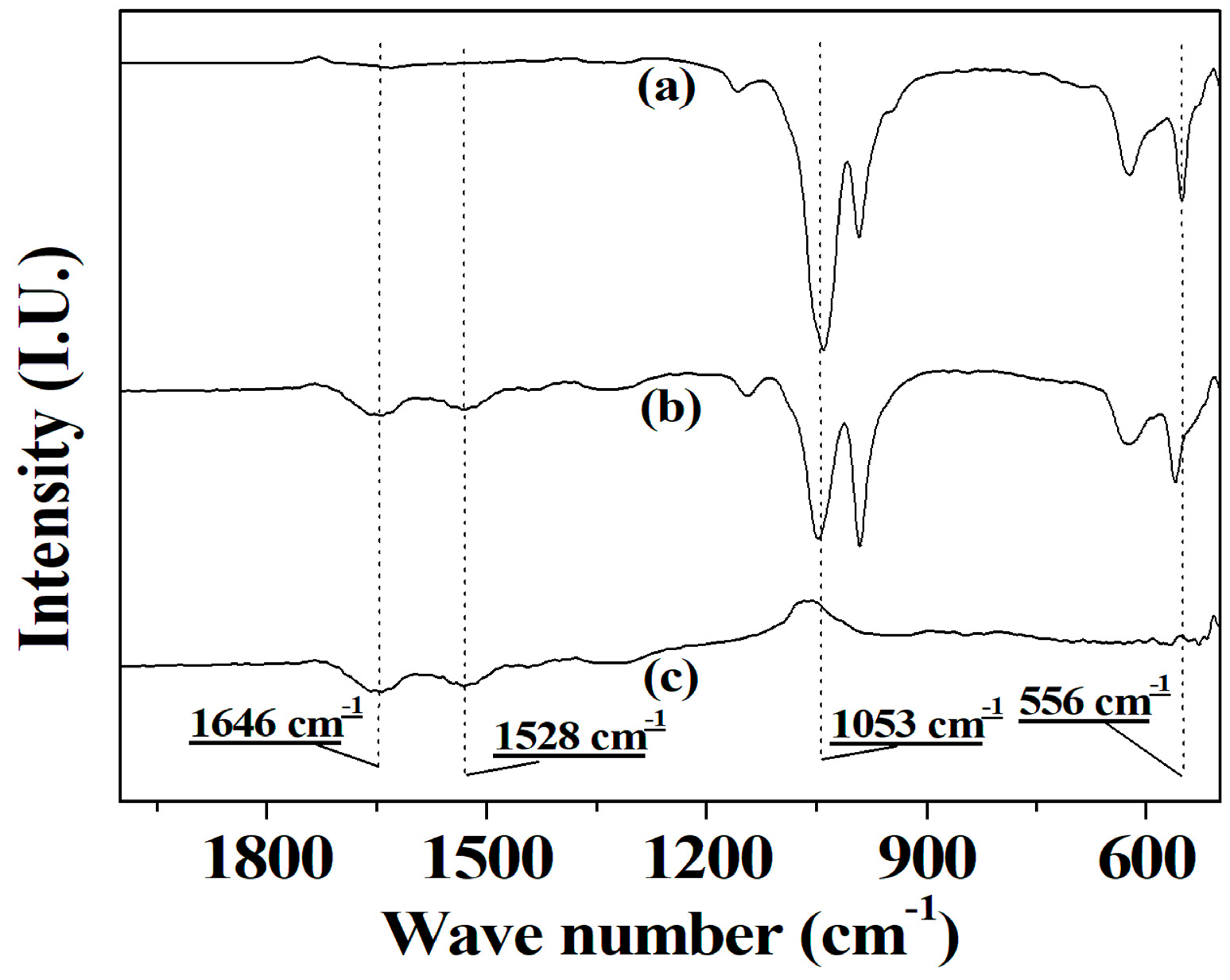
2.2. FTIR Spectra of Co-Immobilized Tri-Enzymes
2.3. Optimization of the Proportion of Enzymes
2.4. Comparison of Activity
2.5. Kinetic Parameters Analysis
2.6. Screening of Reaction Conditions
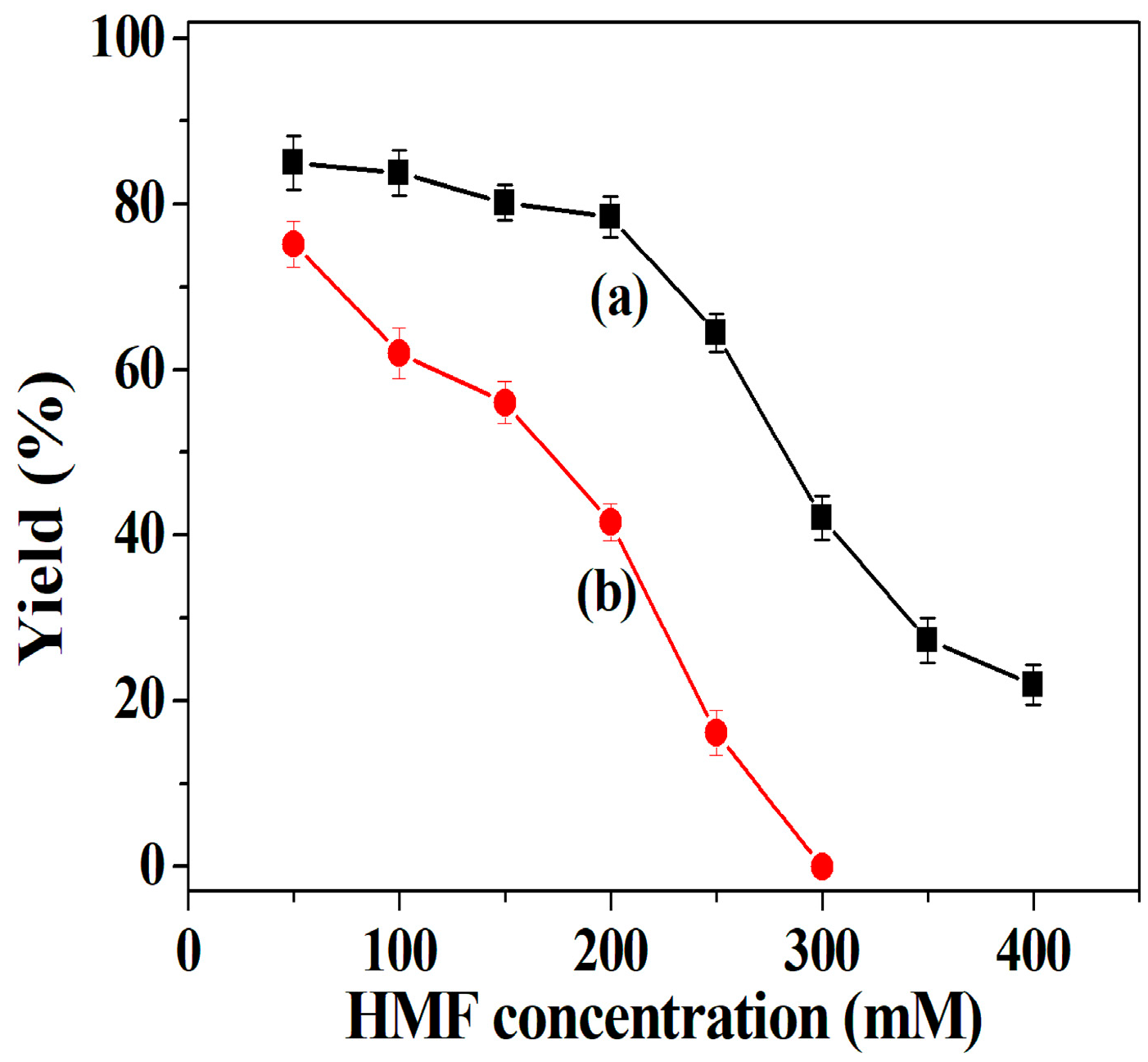
2.6.1. Effect of HMF Concentration
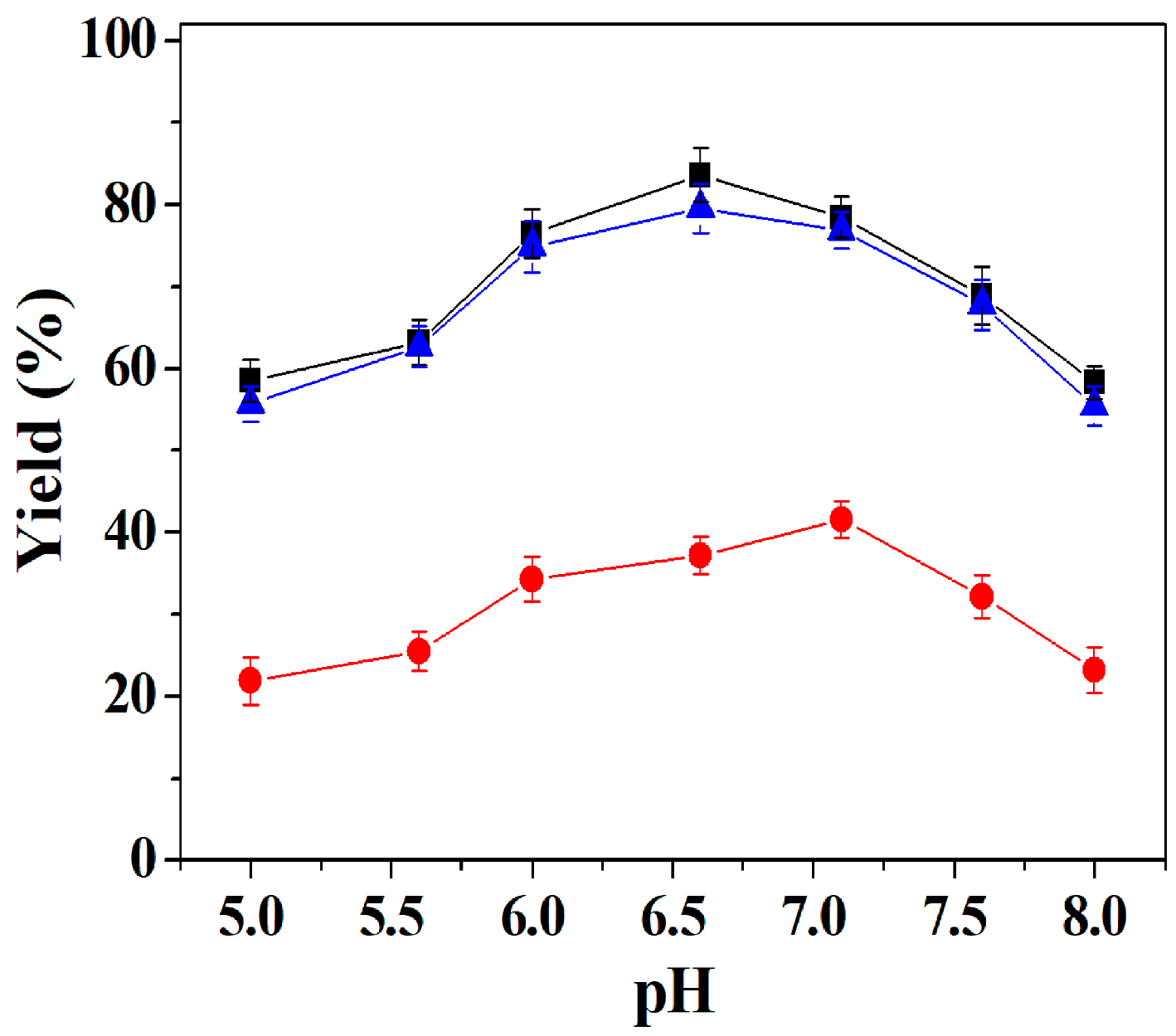
2.6.2. Effect of pH
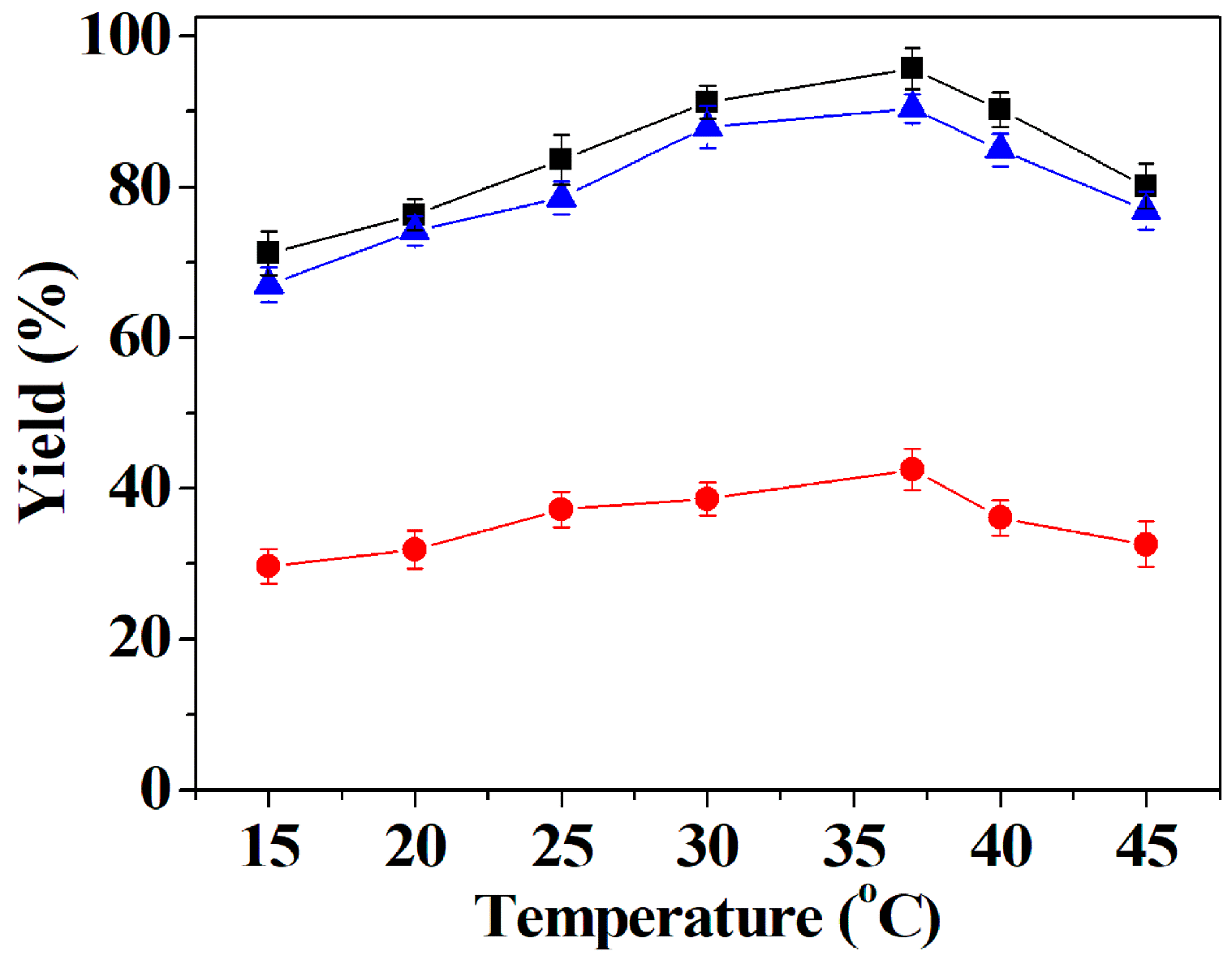
2.6.3. Effect of Temperature
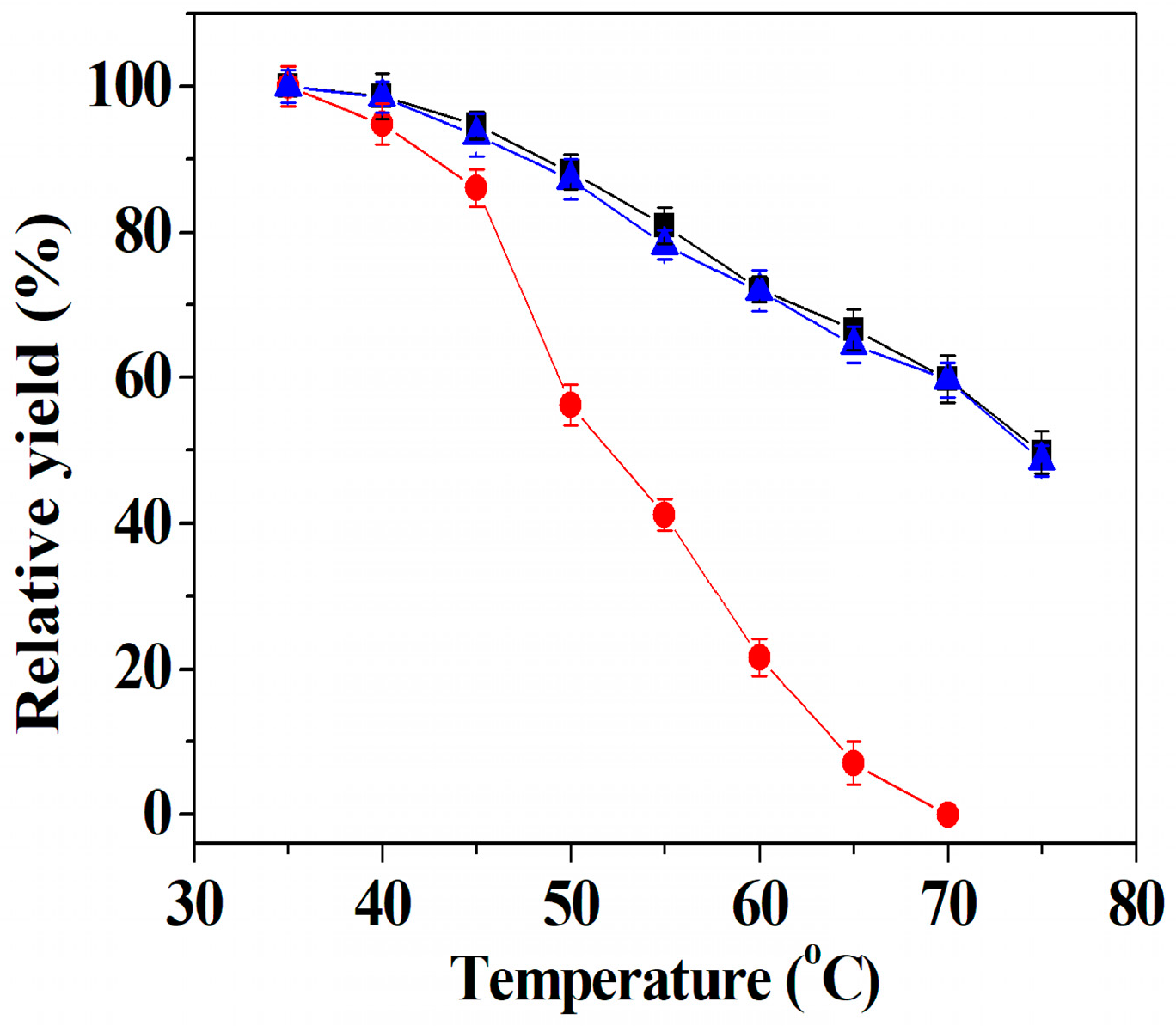
2.7. Thermal Stability
2.8. Comparison of DFF Yield
2.9. Reusability of Co-Immobilized Tri-Enzymes
2.10. Determination of the Level of H2O2 in Different Reaction Systems
3. Materials and Methods
3.1. Materials
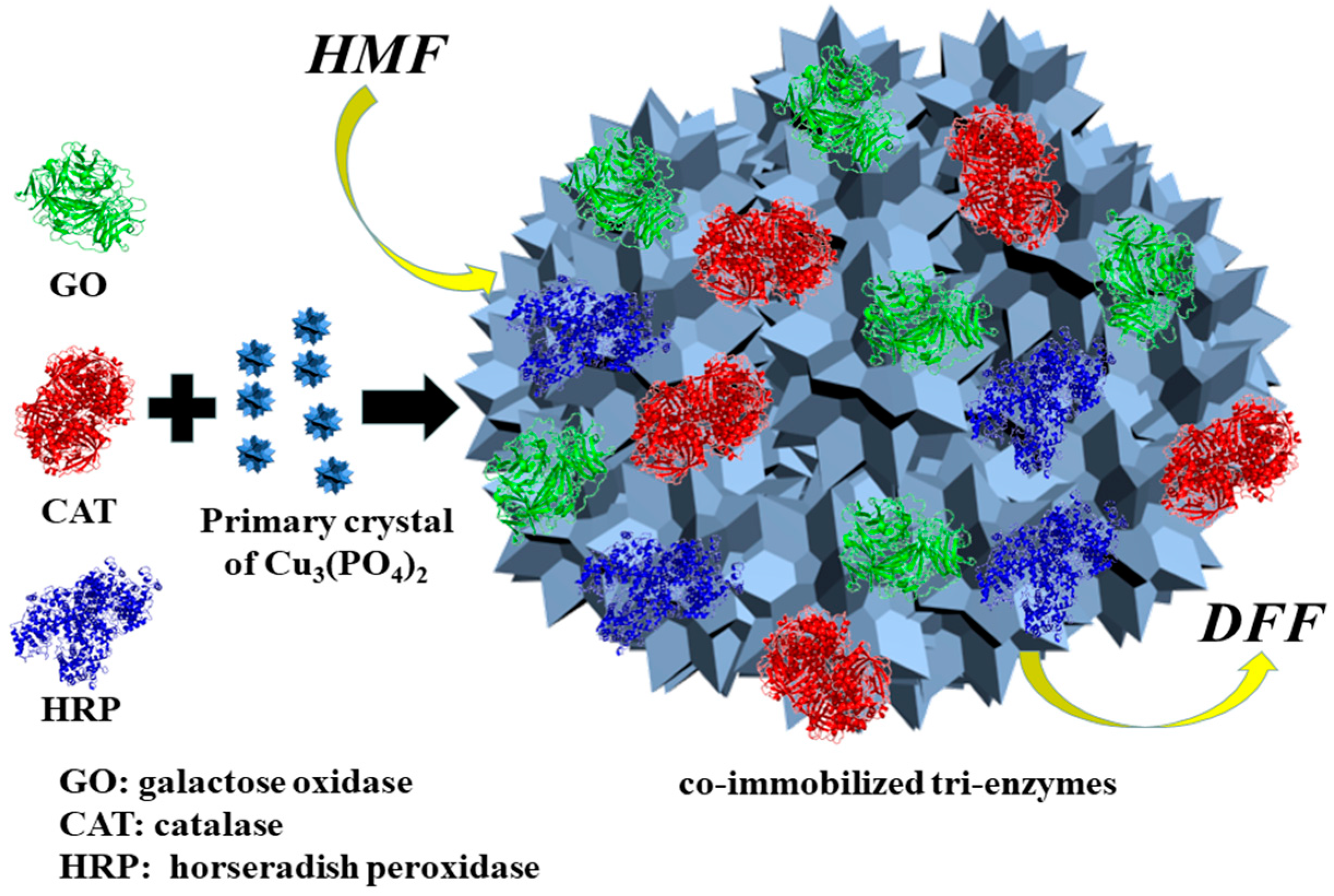
3.2. Co-Immobilization of Tri-Enzymes
3.3. Characteristics of Co-Immobilized Tri-Enzymes
3.4. Enzyme Assay
3.4.1. Galactose Oxidase
3.4.2. Catalase
3.4.3. Horseradish Peroxidase
3.5. Kinetic Analysis
3.6. Oxidation of HMF by Co-Immobilized Tri-Enzymes
3.7. HPLC Analysis
3.8. Thermal Stability
3.9. Reusability
3.10. Determination of the Level of H2O2 Produced during the Oxidation Reaction
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ordomsky, V.V.; Van Der Schaaf, J.; Schouten, J.C.; Nijhuis, T.A. Fructose Dehydration to 5-Hydroxymethylfurfural over Solid Acid Catalysts in a Biphasic System. ChemSusChem 2012, 5, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Nolte, M.W.; Shanks, B.H. Catalytic dehydration of C6carbohydrates for the production of hydroxymethylfurfural (HMF) as a versatile platform chemical. Green Chem. 2014, 16, 548–572. [Google Scholar] [CrossRef]
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—the US Department of Energy’s “Top 10” revisited. Green Chem. 2010, 12, 539. [Google Scholar] [CrossRef]
- Del Poeta, M.; Schell, W.A.; Dykstra, C.C.; Jones, S.; Tidwell, R.R.; Czarny, A.; Bajic, M.; Bajic, M.; Kumar, A.; Boykin, D.; et al. Structure-In Vitro Activity Relationships of Pentamidine Analogues and Dication-Substituted Bis-Benzimidazoles as New Antifungal Agents. Antimicrob. Agents Chemother. 1998, 42, 2495–2502. [Google Scholar] [CrossRef]
- Hopkins, K.T.; Wilson, W.D.; Bender, B.C.; McCurdy, D.R.; Hall, J.E.; Tidwell, R.R.; Kumar, A.; Bajic, M.; Boykin, D.W. Extended aromatic furan amidino derivatives as anti-Pneumocystis carinii agents. J. Med. Chem. 1998, 41, 3872–3878. [Google Scholar] [CrossRef] [PubMed]
- Richter, D.T.; Lash, T.D. Oxidation with dilute aqueous ferric chloride solutions greatly improves yields in the ‘4+ 1’synthesis of sapphyrins. Tetrahedron Lett. 1999, 40, 6735–6738. [Google Scholar] [CrossRef]
- Takimiya, K.; Otsubo, T.; Ogura, F.; Ashitaka, H.; Morita, K.; Suehiro, T. Pyranylidenemethyl- and Thiopyranylidenemethyl-substituted Furans, Thiophenes, and N-Methylpyrroles as Precursors of Organic Metals and Third-order Nonlinear Optical Materials. Chem. Lett. 1994, 23, 255–258. [Google Scholar] [CrossRef]
- Gandini, A.; Belgacem, N.M. Recent advances in the elaboration of polymeric materials derived from biomass components. Polym. Int. 1998, 47, 267–276. [Google Scholar] [CrossRef]
- Gandini, A. Furans in polymer chemistry. Prog. Polym. Sci. 1997, 22, 1203–1379. [Google Scholar] [CrossRef]
- Sheibley, D.W. Cross-Linked Polyvinyl Alcohol Films as Alkaline Battery Separators. J. Electrochem. Soc. 1983, 130, 255. [Google Scholar] [CrossRef]
- Daub, J.; Salbeck, J.; Knöchel, T.; Fischer, C.; Kunkely, H.; Rapp, K.M. Light-Sensitive Molecular Building Blocks with Electron Transfer Activity: Synthesis and Properties of a Photochemically Switchable, Dicyanovinyl-Substituted Furan. Angew. Chem. Int. Ed. 1989, 28, 1494–1496. [Google Scholar] [CrossRef]
- Van Putten, R.-J.; Van Der Waal, J.C.; De Jong, E.; Rasrendra, C.B.; Heeres, H.J.; De Vries, J.G. Hydroxymethylfurfural, A Versatile Platform Chemical Made from Renewable Resources. Chem. Rev. 2013, 113, 1499–1597. [Google Scholar] [CrossRef] [PubMed]
- Cottier, L.; Descotes, G.; Lewkowski, J.; Skowronski, R. ULTRASONICALLY ACCELERATED SYNTHESES OF FURAN-24-DICARBALDEHYDE FROM 5-HYDROXYMETHYL-2-FURFURAL. Org. Prep. Proced. Int. 1995, 27, 564–566. [Google Scholar] [CrossRef]
- Amarasekara, A.S.; Green, D.; McMillan, E. Efficient oxidation of 5-hydroxymethylfurfural to 2,5-diformylfuran using Mn(III)–salen catalysts. Catal. Commun. 2008, 9, 286–288. [Google Scholar] [CrossRef]
- Yoon, H.-J.; Choi, J.-W.; Jang, H.-S.; Cho, J.K.; Byun, J.-W.; Chung, W.-J.; Lee, S.-M.; Lee, Y.-S. Selective oxidation of 5-hydroxymethylfurfural to 2, 5-diformylfuran by polymer-supported IBX amide. Synlett 2011, 2011, 165–168. [Google Scholar]
- Hodgson, E.K.; Fridovich, I. Interaction of bovine erythrocyte superoxide dismutase with hydrogen peroxide. Inactivation of the enzyme. Biochem. 1975, 14, 5294–5299. [Google Scholar] [CrossRef] [PubMed]
- Navarro, O.C.; Canós, A.C.; Chornet, S.I. Chemicals from Biomass: Aerobic Oxidation of 5-Hydroxymethyl-2-Furaldehyde into Diformylfurane Catalyzed by Immobilized Vanadyl-Pyridine Complexes on Polymeric and Organofunctionalized Mesoporous Supports. Top. Catal. 2009, 52, 304–314. [Google Scholar] [CrossRef]
- Carlini, C.; Patrono, P.; Galletti, A.M.R.; Sbrana, G.; Zima, V. Selective oxidation of 5-hydroxymethyl-2-furaldehyde to furan-2,5-dicarboxaldehyde by catalytic systems based on vanadyl phosphate. Appl. Catal. A Gen. 2005, 289, 197–204. [Google Scholar] [CrossRef]
- Ma, J.; Du, Z.; Xu, J.; Chu, Q.; Pang, Y. Efficient Aerobic Oxidation of 5-Hydroxymethylfurfural to 2, 5-Diformylfuran, and Synthesis of a Fluorescent Material. ChemSusChem 2011, 4, 51–54. [Google Scholar] [CrossRef]
- De María, P.D.; Guajardo, N. Biocatalytic Valorization of Furans: Opportunities for Inherently Unstable Substrates. ChemSusChem 2017, 10, 4123–4134. [Google Scholar] [CrossRef]
- McKenna, S.M.; Herter, S.; Turner, N.J.; Leimkühler, S.; Carnell, A.J. Enzyme cascade reactions: synthesis of furandicarboxylic acid (FDCA) and carboxylic acids using oxidases in tandem. Green Chem. 2015, 17, 3271–3275. [Google Scholar] [CrossRef]
- Van Deurzen, M.P.; Van Rantwijk, F.; Sheldon, R.A. Chloroperoxidase-Catalyzed Oxidation of 5-Hydroxymethylfurfural. J. Carbohydr. Chem. 1997, 16, 299–309. [Google Scholar] [CrossRef]
- Qin, Y.-Z.; Li, Y.-M.; Zong, M.-H.; Wu, H.; Li, N. Enzyme-catalyzed selective oxidation of 5-hydroxymethylfurfural (HMF) and separation of HMF and 2,5-diformylfuran using deep eutectic solvents. Green Chem. 2015, 17, 3718–3722. [Google Scholar] [CrossRef]
- Whittaker, M.M.; Whittaker, J.W. The active site of galactose oxidase. J. Boil. Chem. 1988, 263, 6074–6080. [Google Scholar]
- Aebi, H. Catalase. In Methods of enzymatic analysis; Bergmeyer, H.U., Ed.; Elsevier: Amsterdam, The Netherlands, 1974; Volume 1974, pp. 673–684. [Google Scholar]
- Tressel, P.; Kosman, D.J. o,o-Dityrosine in native and horseradish peroxidase-activated galactose oxidase. Biochem. Biophys. Res. Commun. 1980, 92, 781–786. [Google Scholar] [CrossRef]
- Modig, T.; Liden, G.; Taherzadeh, M.J. Inhibition effects of furfural on alcohol dehydrogenase, aldehyde dehydrogenase and pyruvate dehydrogenase. Biochem. J. 2002, 363, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Taherzadeh, M.J.; Gustafsson, L.; Niklasson, C.; Liden, G.; Taherzadeh, M. Physiological effects of 5-hydroxymethylfurfural on Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2000, 53, 701–708. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Ge, J.; Lei, J.; Zare, R.N. Protein–inorganic hybrid nanoflowers. Nat. Nanotechnol. 2012, 7, 428–432. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of Different Enzyme Immobilization Strategies to Improve Enzyme Performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Arana-Peña, S.; Mendez-Sanchez, C.; Rios, N.S.; Ortiz, C.; Gonçalves, L.R.; Fernandez-Lafuente, R. New applications of glyoxyl-octyl agarose in lipases co-immobilization: Strategies to reuse the most stable lipase. Int. J. Boil. Macromol. 2019, 131, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Rueda, N.; Bartolome-Cabrero, R.; Peirce, S.; Virgen-Ortiz, J.J.; Tacias-Pascacio, V.G.; Fernandez-Lopez, L.; Russo, M.E.; Marzocchella, A.; Fernandez-Lafuente, R. Development of simple protocols to solve the problems of enzyme coimmobilization. Application to coimmobilize a lipase and a β-galactosidase. RSC Adv. 2016, 6, 61707–61715. [Google Scholar]
- Zaak, H.; Kornecki, J.F.; Siar, E.-H.; Fernandez-Lopez, L.; Corberán, V.C.; Sassi, M.; Fernandez-Lafuente, R. Coimmobilization of enzymes in bilayers using pei as a glue to reuse the most stable enzyme: Preventing pei release during inactivated enzyme desorption. Process. Biochem. 2017, 61, 95–101. [Google Scholar] [CrossRef]
- Rios, N.S.; Arana-Peña, S.; Mendez-Sanchez, C.; Ortiz, C.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Reuse of Lipase from Pseudomonas fluorescens via Its Step-by-Step Coimmobilization on Glyoxyl-Octyl Agarose Beads with Least Stable Lipases. Catalysts 2019, 9, 487. [Google Scholar] [CrossRef]
- López-Gallego, F.; Schmidt-Dannert, C. Multi-enzymatic synthesis. Curr. Opin. Chem. Boil. 2010, 14, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lafuente, R.; Rodríguez, V.; Guisán, J.M. The coimmobilization of d-amino acid oxidase and catalase enables the quantitative transformation of d-amino acids (d-phenylalanine) into α-keto acids (phenylpyruvic acid). Enzym. Microb. Technol. 1998, 23, 28–33. [Google Scholar] [CrossRef]
- Batencor, L.; Hidalgo, A.; Mateo, C.; Guisan, J.M.; Lopez-Gallego, F.; Fernandez-Lafuente, R. One-Pot Conversion of Cephalosporin C to 7-Aminocephalosporanic Acid in the Absence of Hydrogen Peroxide. Adv. Synth. Catal. 2005, 347, 1804–1810. [Google Scholar]
- Volpato, G.; Rodrigues, R.C.; Fernandez-Lafuente, R.; Rodrigues, R. Use of Enzymes in the Production of Semi-Synthetic Penicillins and Cephalosporins: Drawbacks and Perspectives. Curr. Med. Chem. 2010, 17, 3855–3873. [Google Scholar] [CrossRef]
- Orrego, A.H.; Lopez-Gallego, F.; Espaillat, A.; Cava, F.; Guisan, J.M.; Rocha-Martin, J. One-step Synthesis of α-Keto Acids from Racemic Amino Acids by A Versatile Immobilized Multienzyme Cell-free System. ChemCatChem 2018, 10, 3002–3011. [Google Scholar] [CrossRef]
- García-García, P.; Rocha-Martin, J.; Fernández-Lorente, G.; Guisan, J.M. Co-localization of oxidase and catalase inside a porous support to improve the elimination of hydrogen peroxide: Oxidation of biogenic amines by amino oxidase from Pisum sativum. Enzym. Microb. Technol. 2018, 115, 73–80. [Google Scholar] [CrossRef]
- Rocha-Martín, J.; Acosta, A.; Guisan, J.M.; López-Gallego, F.; Rocha-Martín, J.; Lopez-Gallego, F. Immobilizing Systems Biocatalysis for the Selective Oxidation of Glycerol Coupled to In Situ Cofactor Recycling and Hydrogen Peroxide Elimination. ChemCatChem 2015, 7, 1939–1947. [Google Scholar] [CrossRef]
- Hussain, F.; Arana-Peña, S.; Morellon-Sterling, R.; Barbosa, O.; Braham, S.A.; Kamal, S.; Fernandez-Lafuente, R. Further Stabilization of Alcalase Immobilized on Glyoxyl Supports: Amination Plus Modification with Glutaraldehyde. Molecules 2018, 23, 3188. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.T.R.; Fernandez-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.-S.; Kim, D.W.; Lee, S.; Kwak, C.H.; Bae, S.-T.; Noh, J.H.; Yoon, S.H.; Jung, H.S.; Kim, D.-W.; Hong, K.S. Synthesis of Cu2PO4OH Hierarchical Superstructures with Photocatalytic Activity in Visible Light. Adv. Funct. Mater. 2008, 18, 2154–2162. [Google Scholar] [CrossRef]
- Yang, W.-J.; Griffiths, P.R.; Byler, D.M.; Susi, H. Protein Conformation by Infrared Spectroscopy: Resolution Enhancement by Fourier Self-Deconvolution. Appl. Spectrosc. 1985, 39, 282–287. [Google Scholar] [CrossRef]
- Shaolin, M. Bioelectrochemical response of the polyaniline galactose oxidase electrode. J. Electroanal. Chem. 1994, 370, 135–139. [Google Scholar] [CrossRef]
- Kizilyar, N.; Akbulut, U.; Toppare, L.; Özden, M.; Yağci, Y. Immobilization of invertase in conducting polypyrrole/polytetrahydrofuran graft polymer matrices. Synth. Met. 1999, 104, 45–50. [Google Scholar] [CrossRef]
- Overbeeke, P.L.A.; Ottosson, J.; Hult, K.; Jongejan, J.A.; Duine, J.A. The Temperature Dependence of Enzymatic Kinetic Resolutions Reveals the Relative Contribution of Enthalpy and Entropy to Enzymatic Enantioselectivity. Biocatal. Biotransform. 1999, 17, 61–79. [Google Scholar] [CrossRef]
- Souza, L.T.D.A.; Moreno-Perez, S.; Lorente, G.F.; Cipolatti, E.P.; De Oliveira, D.; Resende, R.R.; Pessela, B.C. Immobilization of Moniliella spathulata R25L270 Lipase on Ionic, Hydrophobic and Covalent Supports: Functional Properties and Hydrolysis of Sardine Oil. Molecules 2017, 22, 1508. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, J.R.; Giordano, R.D.L.C.; Fernandez-Lafuente, R.; Tardioli, P.W. Evaluation of Strategies to Produce Highly Porous Cross-Linked Aggregates of Porcine Pancreas Lipase with Magnetic Properties. Molecules 2018, 23, 2993. [Google Scholar] [CrossRef]
- Verma, M.L.; Rao, N.M.; Tsuzuki, T.; Barrow, C.J.; Puri, M. Suitability of Recombinant Lipase Immobilised on Functionalised Magnetic Nanoparticles for Fish Oil Hydrolysis. Catalysts 2019, 9, 420. [Google Scholar] [CrossRef]
- Tosa, T.; Mori, T.; Fuse, N.; Chibata, I. Immobilization of papain. Enzymologia 1967, 32, 153. [Google Scholar] [PubMed]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-Enzymatic Cascade Reactions: Overview and Perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Turati, D.; Morais Júnior, W.; Terrasan, C.; Moreno-Perez, S.; Pessela, B.; Fernandez-Lorente, G.; Guisan, J.; Carmona, E. Immobilization of lipase from Penicillium sp. section gracilenta (CBMAI 1583) on different hydrophobic supports: Modulation of functional properties. Molecules 2017, 22, 339. [Google Scholar] [CrossRef] [PubMed]
- Molina-Gutiérrez, M.; Hakalin, N.L.S.; Rodríguez-Sánchez, L.; Alcaraz, L.; López, F.A.; Martínez, M.J.; Prieto, A. Effect of the Immobilization Strategy on the Efficiency and Recyclability of the Versatile Lipase from Ophiostoma piceae. Molecules 2019, 24, 1313. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, S.; Li, W.; Li, R.; Chen, S.; Ri, H.I.; Kim, T.M.; Kang, M.S.; Sun, L.; Sun, X.; et al. Improvement of Trehalose Production by Immobilized Trehalose Synthase from Thermus thermophilus HB27. Molecules 2018, 23, 1087. [Google Scholar] [CrossRef]
- Cohen, J.; Karav, S.; Barile, D.; de Moura Bell, J. Immobilization of an Endo-β-N-acetylglucosaminidase for the Release of Bioactive N-glycans. Catalysts 2018, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Li, X.; Li, F.; Yue, H.; He, C.; Xie, F.; Wang, Z. Enantioselective transesterification of (R, S)-2-pentanol catalyzed by a new flower-like nanobioreactor. RSC Adv. 2014, 4, 33998–34002. [Google Scholar] [CrossRef]
- Wu, Z.; Li, H.; Zhu, X.; Li, S.; Wang, Z.; Wang, L.; Li, Z.; Chen, G. Using Laccases in the Nanoflower to Synthesize Viniferin. Catalysts 2017, 7, 188. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lartillot, S.; Kedziora, P.; Athias, A. Purification and Characterization of a New Fungal Catalase. Prep. Biochem. 1988, 18, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A. [136] Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Kengen, S.W.M.; Van Der Oost, J.; De Vos, W.M. Molecular characterization of H2O2-forming NADH oxidases from Archaeoglobus fulgidus. JBIC J. Boil. Inorg. Chem. 2003, 270, 2885–2894. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit Ratio of GA/CAT/HRP Used a | Yield (%) | Concentration of H2O2 Produced from the Oxidation of HMF (μm) |
|---|---|---|
| 0.2/400/25 | 44.8 ± 1.8 | ND |
| 0.5/400/25 | 56.2 ± 2.8 | ND |
| 1.0/400/25 | 64.5 ± 2.3 | 4.9 ± 0.3 |
| 2.0/400/25 | 66.1 ± 2.2 | 40.7 ± 0.3 |
| 4.0/400/25 | 66.4 ± 1.9 | 92.9 ± 0.4 |
| Samples | Activity a (U mg−1) |
|---|---|
| Free galactose oxidase | 166.1 ± 3.2 |
| Galactose oxidase in co-immobilized tri-enzymes | 159.2 ± 2.7 |
| Independently immobilized galactose oxidase | 157.4 ± 2.1 |
| Free catalase | 2270.5 ± 29.4 |
| Catalase in co-immobilized tri-enzymes | 2245.3 ± 24.3 |
| Independently immobilized catalase | 2258.6 ± 28.7 |
| Free horseradish peroxidase | 3658.2 ± 34.2 |
| Horseradish peroxidase in co-immobilized tri-enzymes | 3583.9 ± 42.3 |
| Independently immobilized horseradish peroxidase | 3547.7 ± 36.1 |
| Parameter | Km (mM) | Vmax (µmol min−1 mg−1) |
|---|---|---|
| Free galactose oxidase | 17.6 ± 1.3 | 216.4 ± 5.7 |
| Galactose oxidase in co-immobilized tri-enzymes | 17.8 ± 1.6 | 207.3 ± 3.1 |
| Independently immobilized galactose oxidase | 17.4 ± 1.7 | 205.6 ± 4.7 |
| Free catalase | 26.9 ± 0.1 | 7218.6 ± 26.4 |
| Catalase in co-immobilized tri-enzymes | 25.2± 0.1 | 7015.4 ± 26.6 |
| Independently immobilized catalase | 26.3 ± 0.5 | 7183.8 ± 89.5 |
| Free horseradish peroxidase | 2.21 ± 0.04 | 4377.7 ± 28.4 |
| Horseradish peroxidase in co-immobilized tri-enzymes | 2.12 ± 0.03 | 4356.8 ± 25.2 |
| Independently immobilized horseradish peroxidase | 2.15 ± 0.06 | 4334.2 ± 31.8 |
| Samples | Yield (%) |
|---|---|
| Co-immobilized tri-enzymes a | 95.7 ± 2.7 |
| Immobilized enzymes independently b | 90.4 ± 1.9 |
| Free tri-enzymes c | 53.8 ± 2.3 |
| The nanoflower matrix | ND |
| Catalysts | The Concentration of H2O2 Produced from the Oxidation of HMF (μm) |
|---|---|
| Free tri-enzymes | 253.3 ± 3.7 |
| Independently immobilized tri-enzymes | 27.2 ± 2.2 |
| Co-immobilized tri-enzymes | 2.5 ± 0.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Shi, L.; Yu, X.; Zhang, S.; Chen, G. Co-Immobilization of Tri-Enzymes for the Conversion of Hydroxymethylfurfural to 2,5-Diformylfuran. Molecules 2019, 24, 3648. https://doi.org/10.3390/molecules24203648
Wu Z, Shi L, Yu X, Zhang S, Chen G. Co-Immobilization of Tri-Enzymes for the Conversion of Hydroxymethylfurfural to 2,5-Diformylfuran. Molecules. 2019; 24(20):3648. https://doi.org/10.3390/molecules24203648
Chicago/Turabian StyleWu, Zhuofu, Linjuan Shi, Xiaoxiao Yu, Sitong Zhang, and Guang Chen. 2019. "Co-Immobilization of Tri-Enzymes for the Conversion of Hydroxymethylfurfural to 2,5-Diformylfuran" Molecules 24, no. 20: 3648. https://doi.org/10.3390/molecules24203648
APA StyleWu, Z., Shi, L., Yu, X., Zhang, S., & Chen, G. (2019). Co-Immobilization of Tri-Enzymes for the Conversion of Hydroxymethylfurfural to 2,5-Diformylfuran. Molecules, 24(20), 3648. https://doi.org/10.3390/molecules24203648