
Biochemical Basis of Anti-Cancer-Effects of Phloretin—A Natural Dihydrochalcone


Abstract
1. Introduction
2. Anti-Inflammatory and Anticancer Effects of Phloretin—Evidence from In Vivo Studies
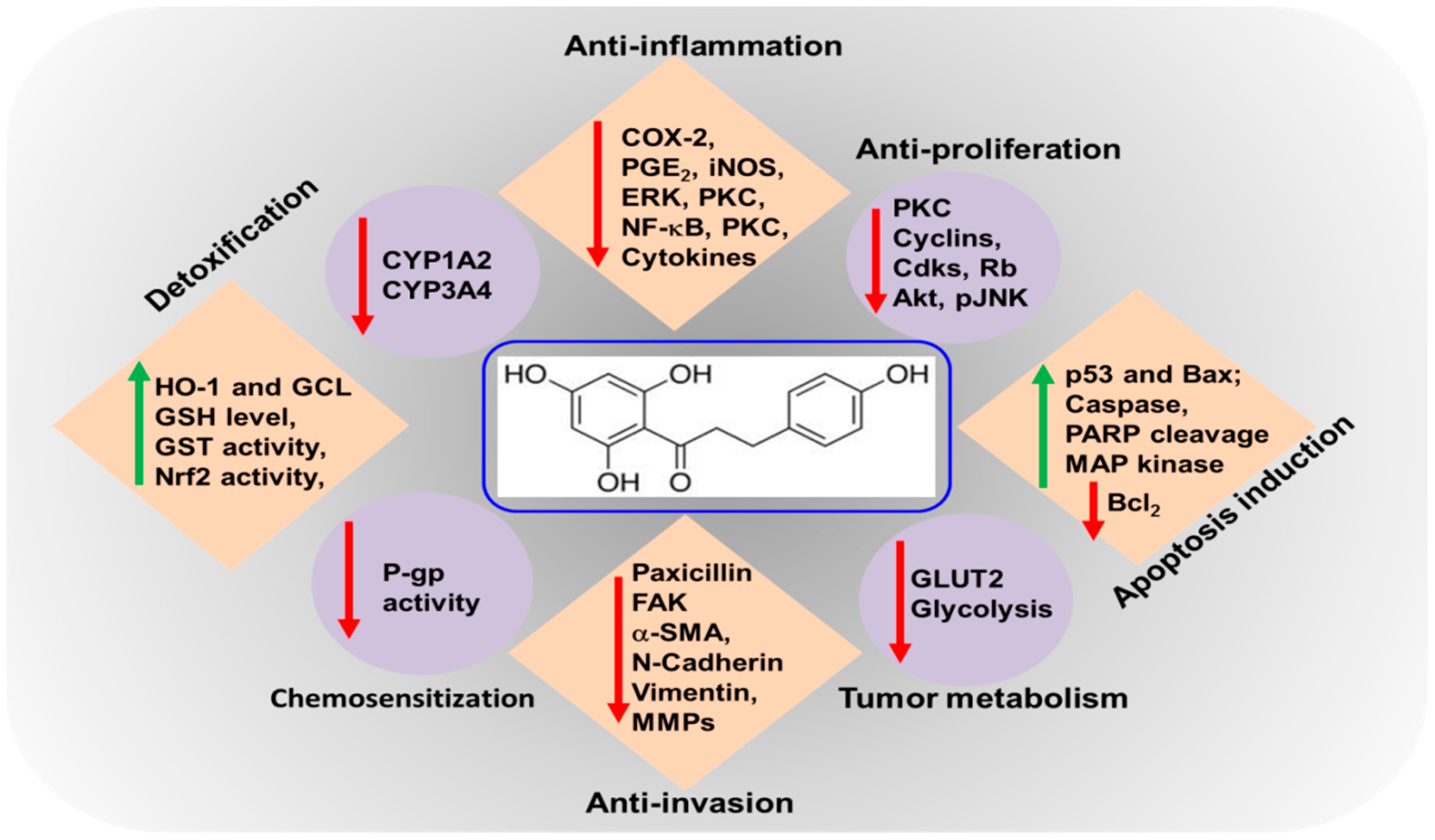
3. The Biochemistry behind the Anti-Inflammatory and Anti-Cancer Effects of Phloretin
3.1. Inhibitory Effects of Phloretin on Inflammatory Markers
3.2. Modulating Biotransformation of Putative Carcinogens
3.3. Antiproliferative and Apoptosis Inducing Effects of Phloretin
3.4. Blockade of Tumor Cell Migration and Invasion
3.5. Inhibition of Glucose Uptake by Phloretin
3.6. Phloretin as a Potential Cancer Immunotherapy Agent
3.7. Phloretin Alleviates Chemotherapy Resistance
4. Pharmacokinetics and Toxicity Profile of Phloretin
5. Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Mann, J.R.; Backlund, M.G.; DuBois, R.N. Mechanisms of disease: Inflammatory mediators and cancer prevention. Nat. Clin. Pract. Oncol. 2005, 2, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Baer-Dubowska, W. Cancer chemopreventive agents-drugs for the 21st century? Acta Pol. Pharm. 2006, 63, 369–373. [Google Scholar] [PubMed]
- Sporn, M.B. Approaches to prevention of epithelial cancer during the preneoplastic period. Cancer Res. 1976, 36, 2699–2702. [Google Scholar] [PubMed]
- Moolgavkar, S.H. The multistage theory of carcinogenesis and the age distribution of cancer in man. J. Natl. Cancer Inst. 1978, 61, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Tsuda, H.; Ohshima, Y.; Nomoto, H.; Fujita, K.; Matsuda, E.; Iigo, M.; Takasuka, N.; Moore, M.A. Cancer prevention by natural compounds. Drug Metab. Pharmacokinet. 2004, 19, 245–263. [Google Scholar] [CrossRef] [PubMed]
- De Flora, S.; Ferguson, L.R. Overview of mechanisms of cancer chemopreventive agents. Mutat. Res. 2005, 591, 8–15. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.J. Breaking the relay in deregulated cellular signal transduction as a rationale for chemoprevention with anti-inflammatory phytochemicals. Mutat. Res. 2005, 591, 123–146. [Google Scholar] [CrossRef]
- Gerhauser, C. Cancer chemopreventive potential of apples, apple juice, and apple components. Planta Med. 2008, 74, 1608–1624. [Google Scholar] [CrossRef]
- Zessner, H.; Pan, L.; Will, F.; Klimo, K.; Knauft, J.; Niewohner, R.; Hummer, W.; Owen, R.; Richling, E.; Frank, N.; et al. Fractionation of polyphenol-enriched apple juice extracts to identify constituents with cancer chemopreventive potential. Mol. Nutr. Food Res. 2008, 52, S28–S44. [Google Scholar] [CrossRef]
- Fini, L.; Piazzi, G.; Daoud, Y.; Selgrad, M.; Maegawa, S.; Garcia, M.; Fogliano, V.; Romano, M.; Graziani, G.; Vitaglione, P.; et al. Chemoprevention of intestinal polyps in ApcMin/+ mice fed with western or balanced diets by drinking annurca apple polyphenol extract. Cancer Prev. Res. 2011, 4, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Watanabe, S.; Miyake, N.; Kohno, H.; Osawa, T. Dihydrochalcones: Evaluation as novel radical scavenging antioxidants. J. Agric. Food Chem 2003, 51, 3309–3312. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Ono, T.; Umeda, M. Inhibition of biological actions of 12-O-tetradecanoylphorbol-13-acetate by inhibitors of protein kinase C. Jpn. J. Cancer Res. 1986, 77, 1107–1113. [Google Scholar] [PubMed]
- Park, S.Y.; Kim, E.J.; Shin, H.K.; Kwon, D.Y.; Kim, M.S.; Surh, Y.J.; Park, J.H. Induction of apoptosis in HT-29 colon cancer cells by phloretin. J. Med. Food 2007, 10, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.C.; Tsai, C.Y.; Wang, Y.J.; Wei, P.L.; Lee, C.H.; Chen, J.H.; Wu, C.H.; Ho, Y.S. Apple polyphenol phloretin potentiates the anti-cancer actions of paclitaxel through induction of apoptosis in human hep G2 cells. Mol. Carcinog. 2009, 48, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Huang, W.C.; Liou, C.J. Evaluation of the anti-inflammatory effects of phloretin and phlorizin in lipopolysaccharide-stimulated mouse macrophages. Food Chem. 2012, 134, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Ho, Y.S.; Tsai, C.Y.; Wang, Y.J.; Tseng, H.; Wei, P.L.; Lee, C.H.; Liu, R.S.; Lin, S.Y. In vitro and in vivo study of phloretin-induced apoptosis in human liver cancer cells involving inhibition of type II glucose transporter. Int. J. Cancer 2009, 124, 2210–2219. [Google Scholar] [CrossRef]
- Min, J.; Huang, K.; Tang, H.; Ding, X.; Qi, C.; Qin, X.; Xu, Z. Phloretin induces apoptosis of non-small cell lung carcinoma A549 cells via JNK1/2 and p38 MAPK pathways. Oncol. Rep. 2015, 34, 2871–2879. [Google Scholar] [CrossRef]
- Shin, J.W.; Kundu, J.K.; Surh, Y.J. Phloretin inhibits phorbol ester-induced tumor promotion and expression of cyclooxygenase-2 in mouse skin: Extracellular signal-regulated kinase and nuclear factor-kappaB as potential targets. J. Med. Food 2012, 15, 253–257. [Google Scholar] [CrossRef]
- Anand, M.A.; Suresh, K. Biochemical profiling and chemopreventive activity of phloretin on 7,12-Dimethylbenz (a) anthracene induced oral carcinogenesis in male golden Syrian hamsters. Toxicol. Int. 2014, 21, 179–185. [Google Scholar]
- Oresajo, C.; Stephens, T.; Hino, P.D.; Law, R.M.; Yatskayer, M.; Foltis, P.; Pillai, S.; Pinnell, S.R. Protective effects of a topical antioxidant mixture containing vitamin C, ferulic acid, and phloretin against ultraviolet-induced photodamage in human skin. J. Cosmet. Dermatol. 2008, 7, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Surh, Y.J. Emerging avenues linking inflammation and cancer. Free Radic. Biol. Med. 2012, 52, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Eaden, J.; Abrams, K.; Ekbom, A.; Jackson, E.; Mayberry, J. Colorectal cancer prevention in ulcerative colitis: A case-control study. Aliment. Pharmacol. Ther. 2000, 14, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Regmi, S.C.; Kim, J.A.; Cho, M.H.; Yun, H.; Lee, C.S.; Lee, J. Apple flavonoid phloretin inhibits Escherichia coli O157:H7 biofilm formation and ameliorates colon inflammation in rats. Infect. Immun. 2011, 79, 4819–4827. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.T.; Tu, S.H.; Yang, P.S.; Hsu, S.P.; Lee, W.H.; Ho, C.T.; Wu, C.H.; Lai, Y.H.; Chen, M.Y.; Chen, L.C. Apple Polyphenol Phloretin Inhibits Colorectal Cancer Cell Growth via Inhibition of the Type 2 Glucose Transporter and Activation of p53-Mediated Signaling. J. Agric. Food Chem. 2016, 64, 6826–6837. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Fang, L.W.; Liou, C.J. Phloretin Attenuates Allergic Airway Inflammation and Oxidative Stress in Asthmatic Mice. Front. Immunol. 2017, 8, 134. [Google Scholar] [CrossRef]
- Wang, H.; Yang, T.; Wang, T.; Hao, N.; Shen, Y.; Wu, Y.; Yuan, Z.; Chen, L.; Wen, F. Phloretin attenuates mucus hypersecretion and airway inflammation induced by cigarette smoke. Int. Immunopharmacol. 2018, 55, 112–119. [Google Scholar] [CrossRef]
- Wu, K.H.; Ho, C.T.; Chen, Z.F.; Chen, L.C.; Whang-Peng, J.; Lin, T.N.; Ho, Y.S. The apple polyphenol phloretin inhibits breast cancer cell migration and proliferation via inhibition of signals by type 2 glucose transporter. J. Food Drug Anal. 2018, 26, 221–231. [Google Scholar] [CrossRef]
- Rezk, B.M.; Haenen, G.R.; van der Vijgh, W.J.; Bast, A. The antioxidant activity of phloretin: The disclosure of a new antioxidant pharmacophore in flavonoids. Biochem. Biophys. Res. Commun. 2002, 295, 9–13. [Google Scholar] [CrossRef]
- Mendes, R.A.; BLS, E.S.; Takeara, R.; Freitas, R.G.; Brown, A.; de Souza, G.L.C. Probing the antioxidant potential of phloretin and phlorizin through a computational investigation. J. Mol. Model. 2018, 24, 101. [Google Scholar] [CrossRef]
- Schaefer, S.; Baum, M.; Eisenbrand, G.; Dietrich, H.; Will, F.; Janzowski, C. Polyphenolic apple juice extracts and their major constituents reduce oxidative damage in human colon cell lines. Mol. Nutr. Food Res. 2006, 50, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Lii, C.K.; Lin, A.H.; Yeh, Y.W.; Yao, H.T.; Li, C.C.; Liu, K.L.; Chen, H.W. Induction of glutathione synthesis and heme oxygenase 1 by the flavonoids butein and phloretin is mediated through the ERK/Nrf2 pathway and protects against oxidative stress. Free Radic. Biol. Med. 2011, 51, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Durai, P.; Jeon, D.; Jung, I.D.; Lee, S.J.; Park, Y.M.; Kim, Y. Phloretin as a Potent Natural TLR2/1 Inhibitor Suppresses TLR2-Induced Inflammation. Nutrients 2018, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Lai, C.L.; Liang, Y.T.; Hung, H.C.; Liu, H.C.; Liou, C.J. Phloretin attenuates LPS-induced acute lung injury in mice via modulation of the NF-kappaB and MAPK pathways. Int. Immunopharmacol. 2016, 40, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Wu, S.J.; Tu, R.S.; Lai, Y.R.; Liou, C.J. Phloretin inhibits interleukin-1beta-induced COX-2 and ICAM-1 expression through inhibition of MAPK, Akt, and NF-kappaB signaling in human lung epithelial cells. Food Funct. 2015, 6, 1960–1967. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Dai, Y.W.; Peng, H.L.; Kang, C.W.; Kuo, C.Y.; Liou, C.J. Phloretin ameliorates chemokines and ICAM-1 expression via blocking of the NF-kappaB pathway in the TNF-alpha-induced HaCaT human keratinocytes. Int. Immunopharmacol. 2015, 27, 32–37. [Google Scholar] [CrossRef]
- Sekhon-Loodu, S.; Ziaullah; Rupasinghe, H.P. Docosahexaenoic acid ester of phloridzin inhibit lipopolysaccharide-induced inflammation in THP-1 differentiated macrophages. Int. Immunopharmacol. 2015, 25, 199–206. [Google Scholar] [CrossRef]
- Chung, M.J.; Sohng, J.K.; Choi, D.J.; Park, Y.I. Inhibitory effect of phloretin and biochanin A on IgE-mediated allergic responses in rat basophilic leukemia RBL-2H3 cells. Life Sci. 2013, 93, 401–408. [Google Scholar] [CrossRef]
- Xu, M.; Gu, W.; Shen, Z.; Wang, F. Anti-cancer Activity of Phloretin Against Human Gastric Cancer Cell Lines Involves Apoptosis, Cell Cycle Arrest, and Inhibition of Cell Invasion and JNK Signalling Pathway. Med. Sci. Monit. 2018, 24, 6551–6558. [Google Scholar] [CrossRef]
- Duan, H.; Wang, R.; Yan, X.; Liu, H.; Zhang, Y.; Mu, D.; Han, J.; Li, X. Phloretin induces apoptosis of human esophageal cancer via a mitochondria-dependent pathway. Oncol. Lett. 2017, 14, 6763–6768. [Google Scholar] [CrossRef]
- Ma, L.; Wang, R.; Nan, Y.; Li, W.; Wang, Q.; Jin, F. Phloretin exhibits an anti-cancer effect and enhances the anti-cancer ability of cisplatin on non-small cell lung cancer cell lines by regulating expression of apoptotic pathways and matrix metalloproteinases. Int. J. Oncol. 2016, 48, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Abkin, S.V.; Ostroumova, O.S.; Komarova, E.Y.; Meshalkina, D.A.; Shevtsov, M.A.; Margulis, B.A.; Guzhova, I.V. Phloretin increases the anti-tumor efficacy of intratumorally delivered heat-shock protein 70 kDa (HSP70) in a murine model of melanoma. Cancer Immunol. Immunother. 2016, 65, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Xing, Y.F.; Zhou, Z.; Yao, Y. Dihydrochalcone Compounds Isolated from Crabapple Leaves Showed Anti-cancer Effects on Human Cancer Cell Lines. Molecules 2015, 20, 21193–21203. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Oshima, M.; Inaba, K.; Taketo, M.M. Hyperplastic gastric tumors induced by activated macrophages in COX-2/mPGES-1 transgenic mice. EMBO J. 2004, 23, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Muller-Decker, K.; Neufang, G.; Berger, I.; Neumann, M.; Marks, F.; Furstenberger, G. Transgenic cyclooxygenase-2 overexpression sensitizes mouse skin for carcinogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 12483–12488. [Google Scholar] [CrossRef] [PubMed]
- Oshima, M.; Dinchuk, J.E.; Kargman, S.L.; Oshima, H.; Hancock, B.; Kwong, E.; Trzaskos, J.M.; Evans, J.F.; Taketo, M.M. Suppression of intestinal polyposis in Apc delta716 knockout mice by inhibition of cyclooxygenase 2 (COX-2). Cell 1996, 87, 803–809. [Google Scholar] [CrossRef]
- Tiano, H.F.; Loftin, C.D.; Akunda, J.; Lee, C.A.; Spalding, J.; Sessoms, A.; Dunson, D.B.; Rogan, E.G.; Morham, S.G.; Smart, R.C.; et al. Deficiency of either cyclooxygenase (COX)-1 or COX-2 alters epidermal differentiation and reduces mouse skin tumorigenesis. Cancer Res. 2002, 62, 3395–3401. [Google Scholar]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Hata, K.; Sugie, S.; Niho, N.; Sakano, K.; Takahashi, M.; Wakabayashi, K. Dextran sodium sulfate strongly promotes colorectal carcinogenesis in Apc(Min/+) mice: Inflammatory stimuli by dextran sodium sulfate results in development of multiple colonic neoplasms. Int. J. Cancer 2006, 118, 25–34. [Google Scholar] [CrossRef]
- Chun, K.S.; Cha, H.H.; Shin, J.W.; Na, H.K.; Park, K.K.; Chung, W.Y.; Surh, Y.J. Nitric oxide induces expression of cyclooxygenase-2 in mouse skin through activation of NF-kappaB. Carcinogenesis 2004, 25, 445–454. [Google Scholar] [CrossRef]
- Surh, Y.J.; Kundu, J.K.; Na, H.K.; Lee, J.S. Redox-sensitive transcription factors as prime targets for chemoprevention with anti-inflammatory and antioxidative phytochemicals. J. Nutr. 2005, 135, 2993S–3001S. [Google Scholar] [CrossRef]
- Jung, M.; Triebel, S.; Anke, T.; Richling, E.; Erkel, G. Influence of apple polyphenols on inflammatory gene expression. Mol. Nutr. Food Res. 2009, 53, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Alsanea, S.; Gao, M.; Liu, D. Phloretin Prevents High-Fat Diet-Induced Obesity and Improves Metabolic Homeostasis. AAPS J. 2017, 19, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Park, S.H.; Han, S.Y.; Kim, Y.H.; Lee, E.J.; Yoon Park, J.H.; Kang, Y.H. Phloretin suppresses thrombin-mediated leukocyte-platelet-endothelial interactions. Mol. Nutr. Food Res. 2014, 58, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.S.; Chen, X.Y.; Zhu, R.Z.; Choi, B.M.; Kim, S.J.; Kim, B.R. Dual effects of phloretin on aflatoxin B1 metabolism: Activation and detoxification of aflatoxin B1. Biofactors 2012, 38, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kwon, J.Y.; Kang, N.J.; Lee, K.W.; Lee, H.J. Phloretin induces apoptosis in H-Ras MCF10A human breast tumor cells through the activation of p53 via JNK and p38 mitogen-activated protein kinase signaling. Ann. N. Y. Acad. Sci. 2009, 1171, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Kobori, M.; Shinmoto, H.; Tsushida, T.; Shinohara, K. Phloretin-induced apoptosis in B16 melanoma 4A5 cells by inhibition of glucose transmembrane transport. Cancer Lett. 1997, 119, 207–212. [Google Scholar] [CrossRef]
- Kobori, M.; Iwashita, K.; Shinmoto, H.; Tsushida, T. Phloretin-induced apoptosis in B16 melanoma 4A5 cells and HL60 human leukemia cells. Biosci. Biotechnol. Biochem. 1999, 63, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Czuba, Z.P.; Mazur, B.; Paradysz, A.; Krol, W. Chalcones and dihydrochalcones augment TRAIL-mediated apoptosis in prostate cancer cells. Molecules 2010, 15, 5336–5353. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fan, C.; Pu, L.; Wei, C.; Jin, H.; Teng, Y.; Zhao, M.; Yu, A.C.; Jiang, F.; Shu, J.; et al. Phloretin induces cell cycle arrest and apoptosis of human glioblastoma cells through the generation of reactive oxygen species. J. Neurooncol. 2016, 128, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Ma, W.; An, L. Cortactin in cancer cell migration and invasion. Oncotarget 2017, 8, 88232–88243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zou, Y.; Liang, M.; Chen, Y.; Luo, Y.; Yang, B.; Liu, F.; Qin, Y.; He, D.; Wang, F.; et al. Suppressor of fused (Sufu) promotes epithelial-mesenchymal transition (EMT) in cervical squamous cell carcinoma. Oncotarget 2017, 8, 114226–114238. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Park, Y.S.; Kim, H.J.; Kim, C.H.; Lim, S.W.; Huh, J.W.; Lee, J.H.; Kim, H.R. CD44 enhances the epithelial-mesenchymal transition in association with colon cancer invasion. Int. J. Oncol. 2012, 41, 211–218. [Google Scholar] [PubMed]
- Cao, X.; Fang, L.; Gibbs, S.; Huang, Y.; Dai, Z.; Wen, P.; Zheng, X.; Sadee, W.; Sun, D. Glucose uptake inhibitor sensitizes cancer cells to daunorubicin and overcomes drug resistance in hypoxia. Cancer Chemother. Pharmacol. 2007, 59, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.A.; Falk, R.E. The efficacy of phloridzin and phloretin on tumor cell growth. Anti-Cancer Res. 1993, 13, 2287–2292. [Google Scholar]
- Eno, J. Immunotherapy Through the Years. J. Adv. Pract. Oncol. 2017, 8, 747–753. [Google Scholar] [PubMed]
- Zhu, S.P.; Liu, G.; Wu, X.T.; Chen, F.X.; Liu, J.Q.; Zhou, Z.H.; Zhang, J.F.; Fei, S.J. The effect of phloretin on human gammadelta T cells killing colon cancer SW-1116 cells. Int. Immunopharmacol. 2013, 15, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Ma, J.; Zhou, W.; Zhou, X.; Cao, B.; Zhang, H.; Zhao, Q.; Fan, D.; Hong, L. Molecular mechanisms and clinical implications of miRNAs in drug resistance of esophageal cancer. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 1151–1163. [Google Scholar] [CrossRef]
- Goldstein, L.J. Clinical reversal of drug resistance. Curr. Probl. Cancer 1995, 19, 65–124. [Google Scholar] [CrossRef]
- Bai, X.; Chen, Y.; Hou, X.; Huang, M.; Jin, J. Emerging role of NRF2 in chemoresistance by regulating drug-metabolizing enzymes and efflux transporters. Drug Metab. Rev. 2016, 48, 541–567. [Google Scholar] [CrossRef]
- Xue, X.; Liang, X.J. Overcoming drug efflux-based multidrug resistance in cancer with nanotechnology. Chin. J. Cancer 2012, 31, 100–109. [Google Scholar] [CrossRef]
- Nguyen, H.; Zhang, S.; Morris, M.E. Effect of flavonoids on MRP1-mediated transport in Panc-1 cells. J. Pharm. Sci. 2003, 92, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Molnar, J.; Engi, H.; Hohmann, J.; Molnar, P.; Deli, J.; Wesolowska, O.; Michalak, K.; Wang, Q. Reversal of multidrug resitance by natural substances from plants. Curr. Top. Med. Chem. 2010, 10, 1757–1768. [Google Scholar] [PubMed]
- Choi, B.M.; Chen, X.Y.; Gao, S.S.; Zhu, R.; Kim, B.R. Anti-apoptotic effect of phloretin on cisplatin-induced apoptosis in HEI-OC1 auditory cells. Pharmacol. Rep. 2011, 63, 708–716. [Google Scholar] [CrossRef]
- Zhou, M.; Zheng, J.; Bi, J.; Wu, X.; Lyu, J.; Gao, K. Synergistic inhibition of colon cancer cell growth by a combination of atorvastatin and phloretin. Oncol. Lett. 2018, 15, 1985–1992. [Google Scholar] [CrossRef]
- Adam, A.; Crespy, V.; Levrat-Verny, M.A.; Leenhardt, F.; Leuillet, M.; Demigne, C.; Remesy, C. The bioavailability of ferulic acid is governed primarily by the food matrix rather than its metabolism in intestine and liver in rats. J. Nutr. 2002, 132, 1962–1968. [Google Scholar] [CrossRef] [PubMed]
- Kahle, K.; Kraus, M.; Scheppach, W.; Richling, E. Colonic availability of apple polyphenols--a study in ileostomy subjects. Mol. Nutr. Food Res. 2005, 49, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Deusser, H.; Rogoll, D.; Scheppach, W.; Volk, A.; Melcher, R.; Richling, E. Gastrointestinal absorption and metabolism of apple polyphenols ex vivo by the pig intestinal mucosa in the Ussing chamber. Biotechnol. J. 2013, 8, 363–370. [Google Scholar] [CrossRef]
- Geohagen, B.C.; Korsharskyy, B.; Vydyanatha, A.; Nordstroem, L.; LoPachin, R.M. Phloretin cytoprotection and toxicity. Chem. Biol. Interact. 2018, 296, 117–123. [Google Scholar] [CrossRef]


{kind=link}
| Type | Experimental Model | Dose/Concentration | Mechanism of Action | Ref. | |
|---|---|---|---|---|---|
| Anti-inflammatory | HEK293 cells engineered to either overexpress or deficient in hTLR2 | 10 or 20 µM | Inhibits the heterodimerization of TLR2/1; reduced the secretion of TNF-α and IL-8 | [33] | |
| Specific pathogen-free male BALB/c mice (6–8 weeks, 22–24 g) | 20 mg/kg | Suppressed the mucins secretion, inflammatory cell infiltration and cytokine release in mouse lungs induced by cigarette smoke (CS) | [27] | ||
| OVA-challenged asthmatic mice | 5, 10, or 20 mg/kg | Decreased hyperresponsiveness, inflammation, and oxidative responses; reduced ROS, and cytokines production | [26] | ||
| LPS-induced acute lung injury in mice | 5 or 20 mg/kg | Suppressed LPS-induced neutrophil infiltration, and reduce the levels of IL-6 and TNF-α in serum and bronchoalveolar lavage fluid; blockade of the NF-κB and MAPK pathways | [34] | ||
| A549 cells | 3–100 μM | Inhibited proinflammatory cytokine, COX-2, and ICAM-1 expression; blocked NF-κB and MAPK signaling pathways. | [35] | ||
| TNF-α-stimulated HaCaT human keratinocytes | 10, 30, or 100 μM | Decreased the production of IL-6, IL-8, and CCL5; inhibited NF-κB nuclear translocation; suppressed phosphorylation of Akt and MAPK signal. | [36] | ||
| Human THP-1 monocytes | 1, 10, or 30 μg/mL | Reduced TNF-α, IL-6 and COX-2 expression | [37] | ||
| Rat basophilic leukemia RBL-2H3 cells | 12.5, 25, or 50 μM | Attenuated ROS production, phosphorylation of Akt, ERK1/2, p38 MAP kinase, and JNK | [38] | ||
| LPS-stimulated murine RAW264.7 macrophages | 3, 10, 30, or 100 μM | Reduced the levels of NO, PGE2, IL-6, TNF-α, iNOS and COX-2; suppressed nuclear translocation of NF-κB subunit p65 proteins, and decreased phosphorylation of MAPK pathways | [16] | ||
| Anti-cancer | Gastric cancer (AGS) cells | IC50 8 μM | Arrested the cell cycle in G2/M phase and decreased the expression of p-JNK and p-p38 MAP kinase | [39] | |
| Esophageal cancer EC-109 cell lines | 60 µg/mL | Apoptosis increased to 225.6 ± 16.0%; increased p53 activity; increased the level of Bax and declined Bcl-2 levels | [40] | ||
| Non-small cell lung cancer (NSCLC): A549, Calu-1, H838 and H520 cells | 25, 50 or 75 μg/mL | Suppressed the expression of Bcl-2; increased the protein expression of cleaved-caspase-3 and -9, and deregulated the expression of MMP-2 and -9 on gene and protein levels | [41] | ||
| Human erythroid leukemia K-562 cells | 20 μM | Increased the efficacy of HSP70 penetration; increases anti-tumor activity of HSP70 with phloretin combination | [42] | ||
| A549 human lung cancer cell line, Bel 7402 liver cancer cell line, HepG2 human ileocecal cancer cell line, and HT-29 human colon cancer cell line | 0–150 mg/mL | Significant positive anti-cancer activities against several human cancer cell lines, IC50: A549 (27 μg/mL), BEL7402 (37 μg/mL, HepG2 (37 μg/mL), HT29 (33 μg/mL) | [43] | ||
| HepG2-xenografted tumor | 10 mg/kg phloretin or +1 mg/kg paclitaxel | Reduced tumor growth more than fivefold in the phloretin and paclitaxel-treated mice compared to the paclitaxel only treated mice | [15] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.Y. Biochemical Basis of Anti-Cancer-Effects of Phloretin—A Natural Dihydrochalcone. Molecules 2019, 24, 278. https://doi.org/10.3390/molecules24020278
Choi BY. Biochemical Basis of Anti-Cancer-Effects of Phloretin—A Natural Dihydrochalcone. Molecules. 2019; 24(2):278. https://doi.org/10.3390/molecules24020278
Chicago/Turabian StyleChoi, Bu Young. 2019. "Biochemical Basis of Anti-Cancer-Effects of Phloretin—A Natural Dihydrochalcone" Molecules 24, no. 2: 278. https://doi.org/10.3390/molecules24020278
APA StyleChoi, B. Y. (2019). Biochemical Basis of Anti-Cancer-Effects of Phloretin—A Natural Dihydrochalcone. Molecules, 24(2), 278. https://doi.org/10.3390/molecules24020278