
Array of Metabolites in Italian Hericium erinaceus Mycelium, Primordium, and Sporophore

 , , ,
, , , 


Abstract
:1. Introduction
2. Results
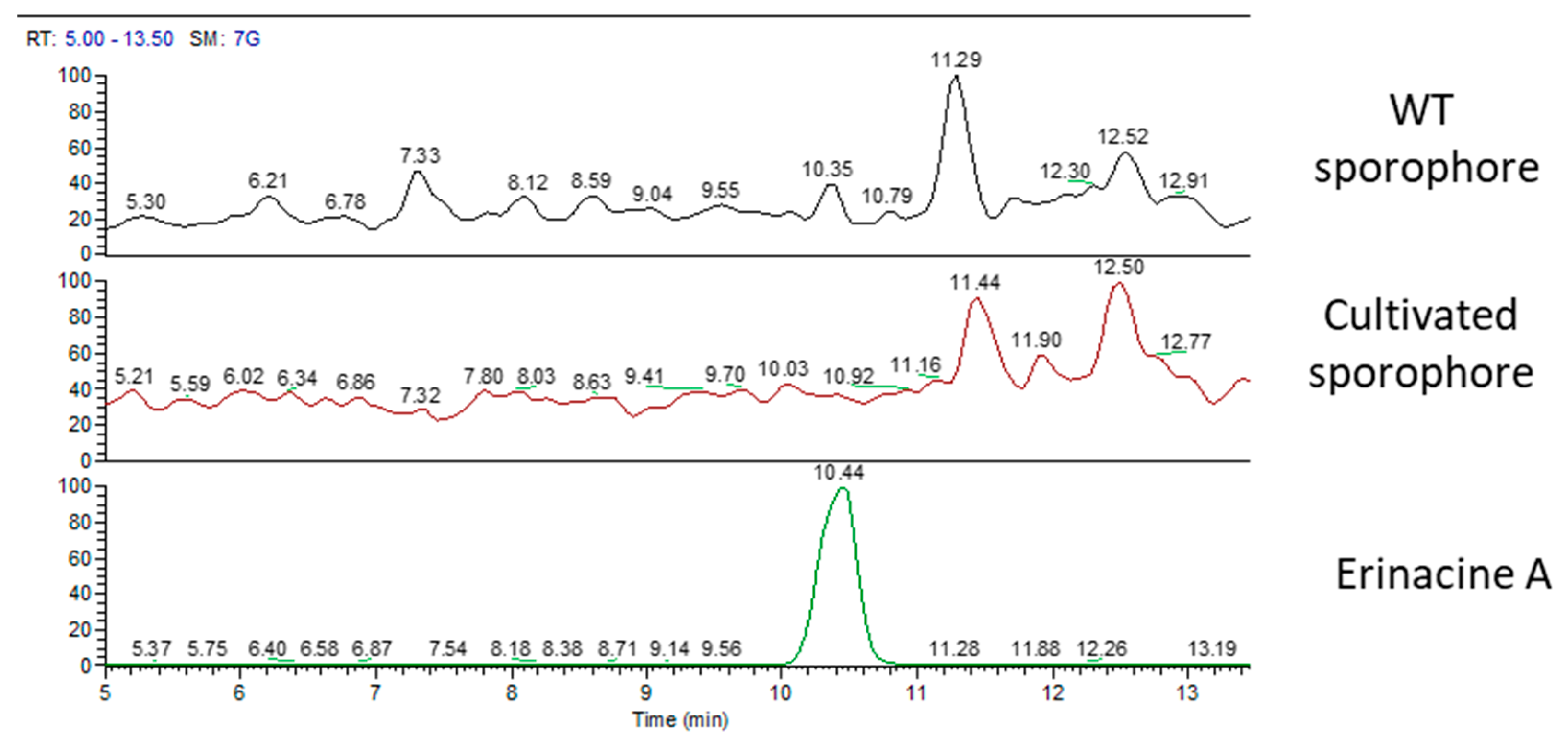
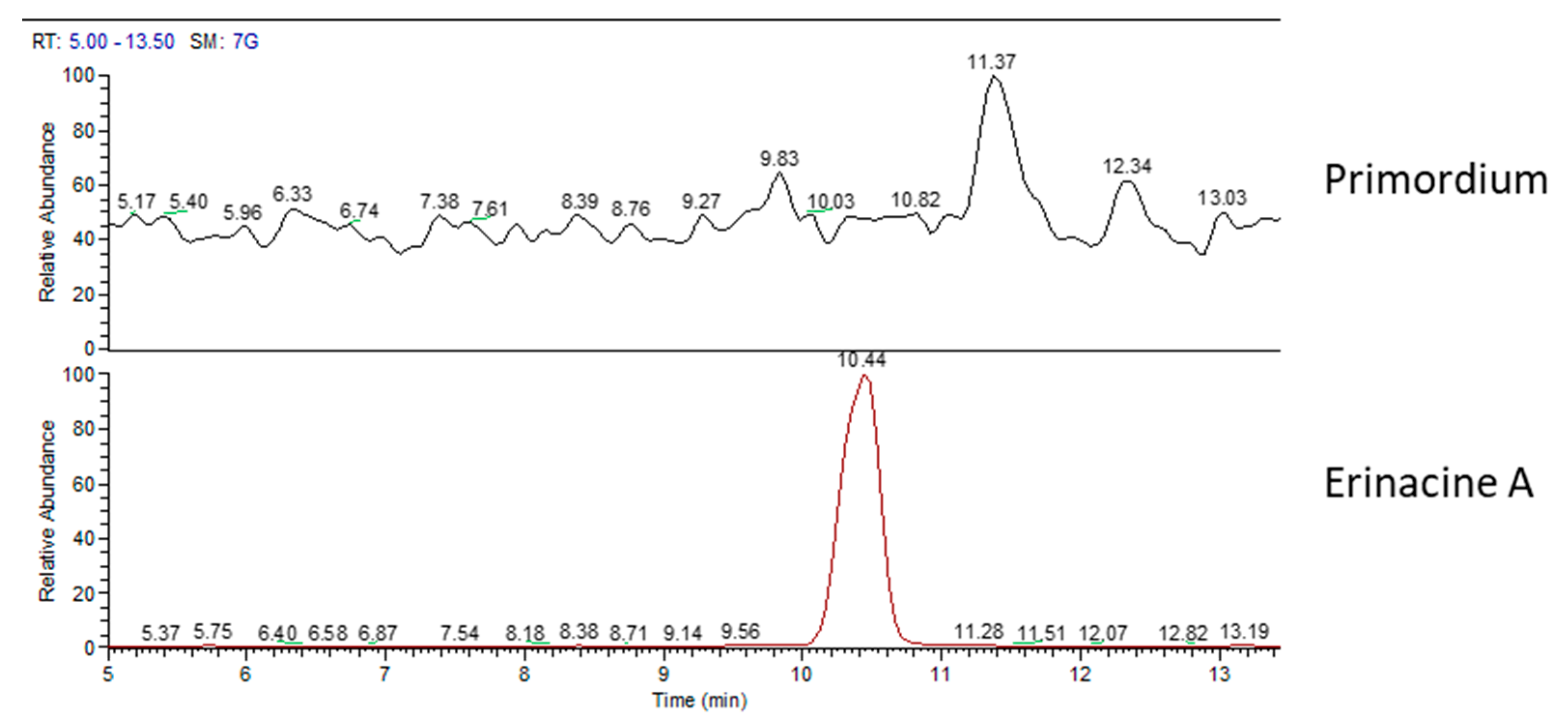
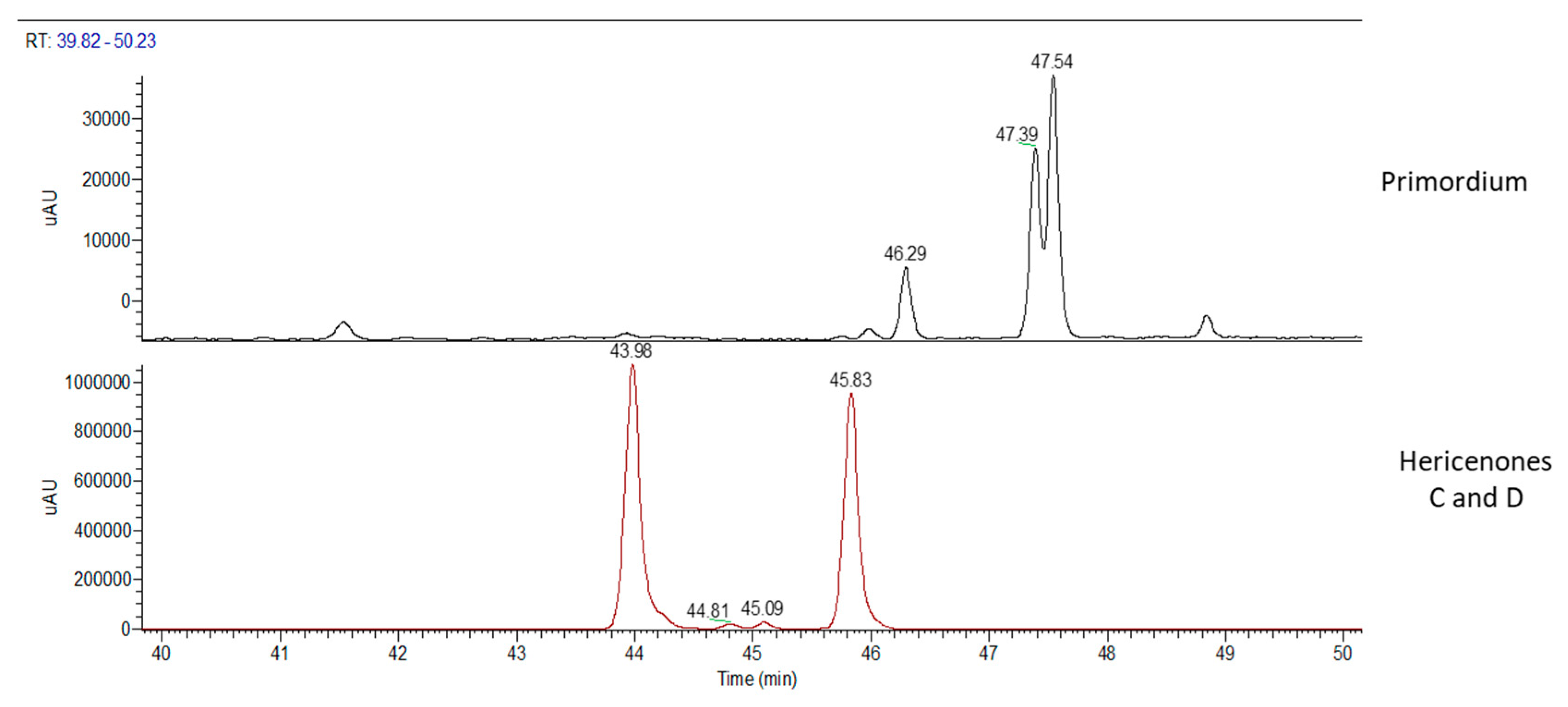
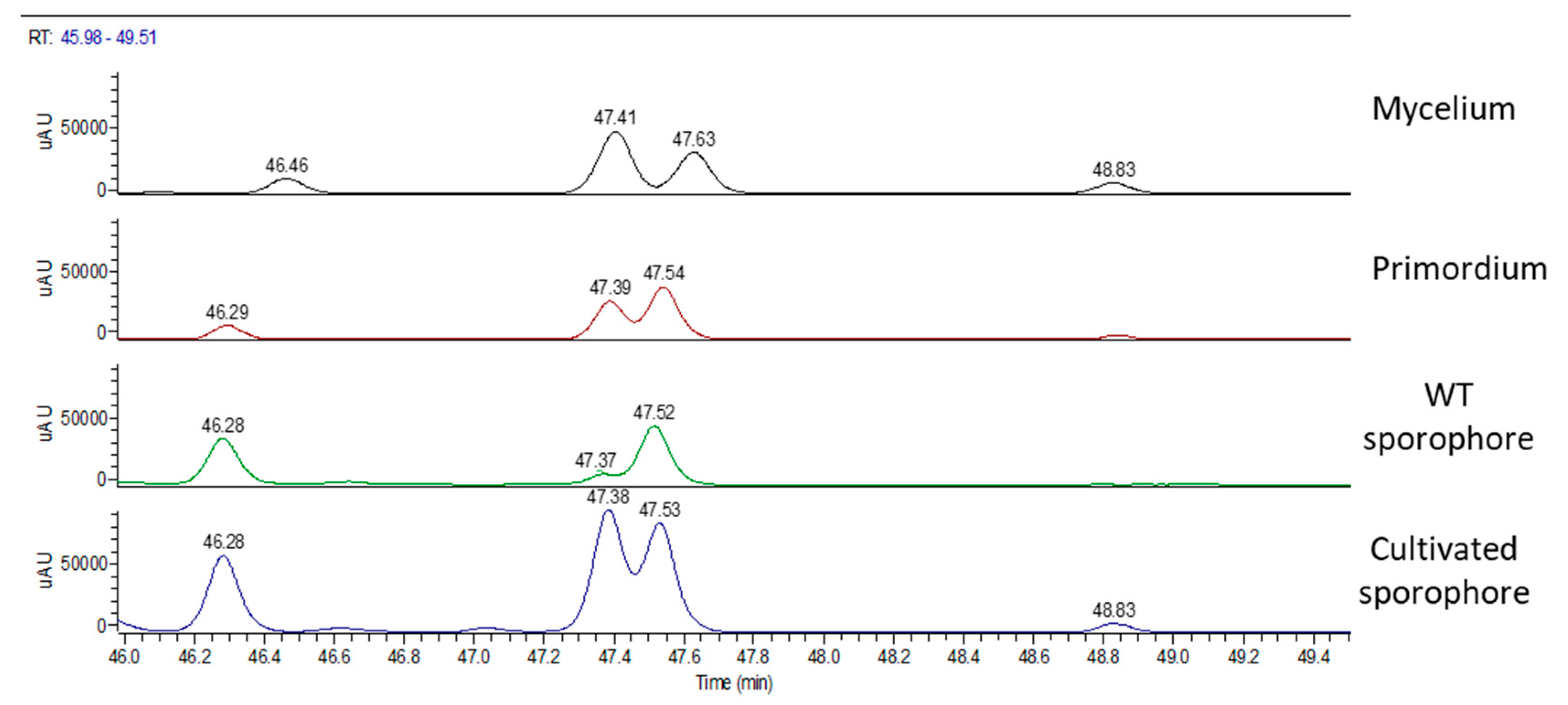
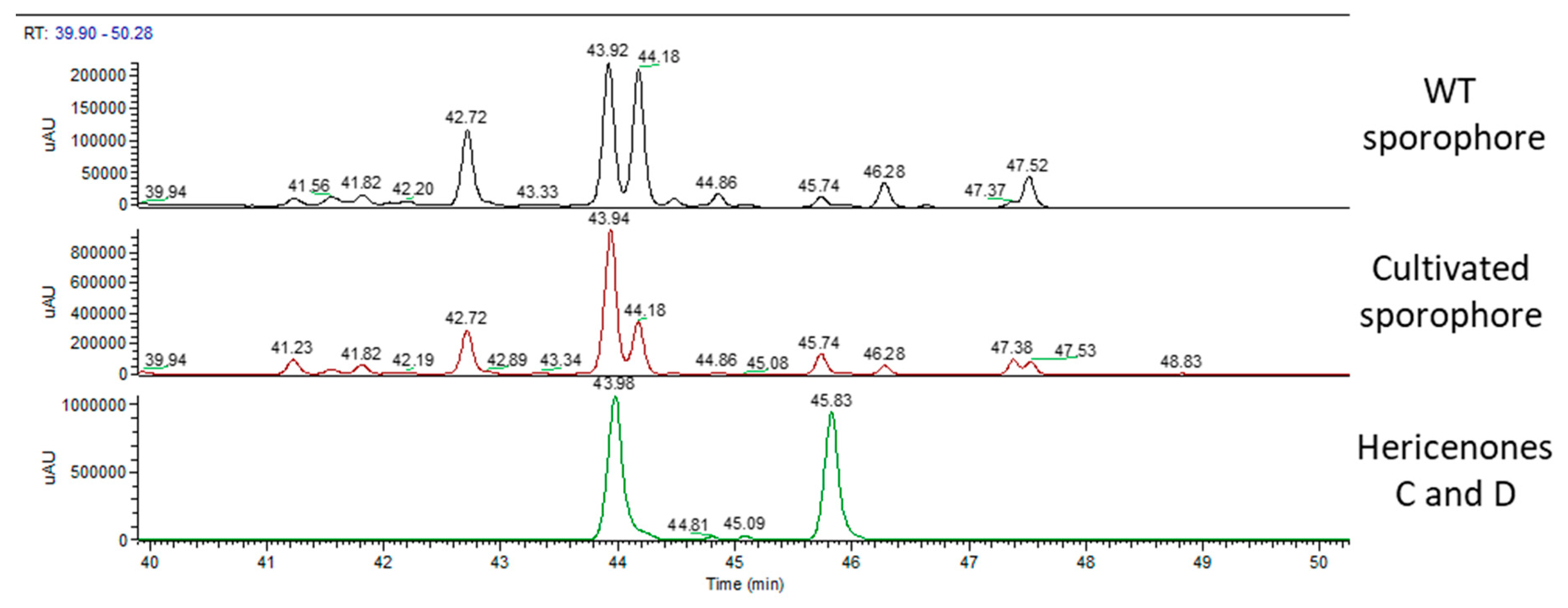
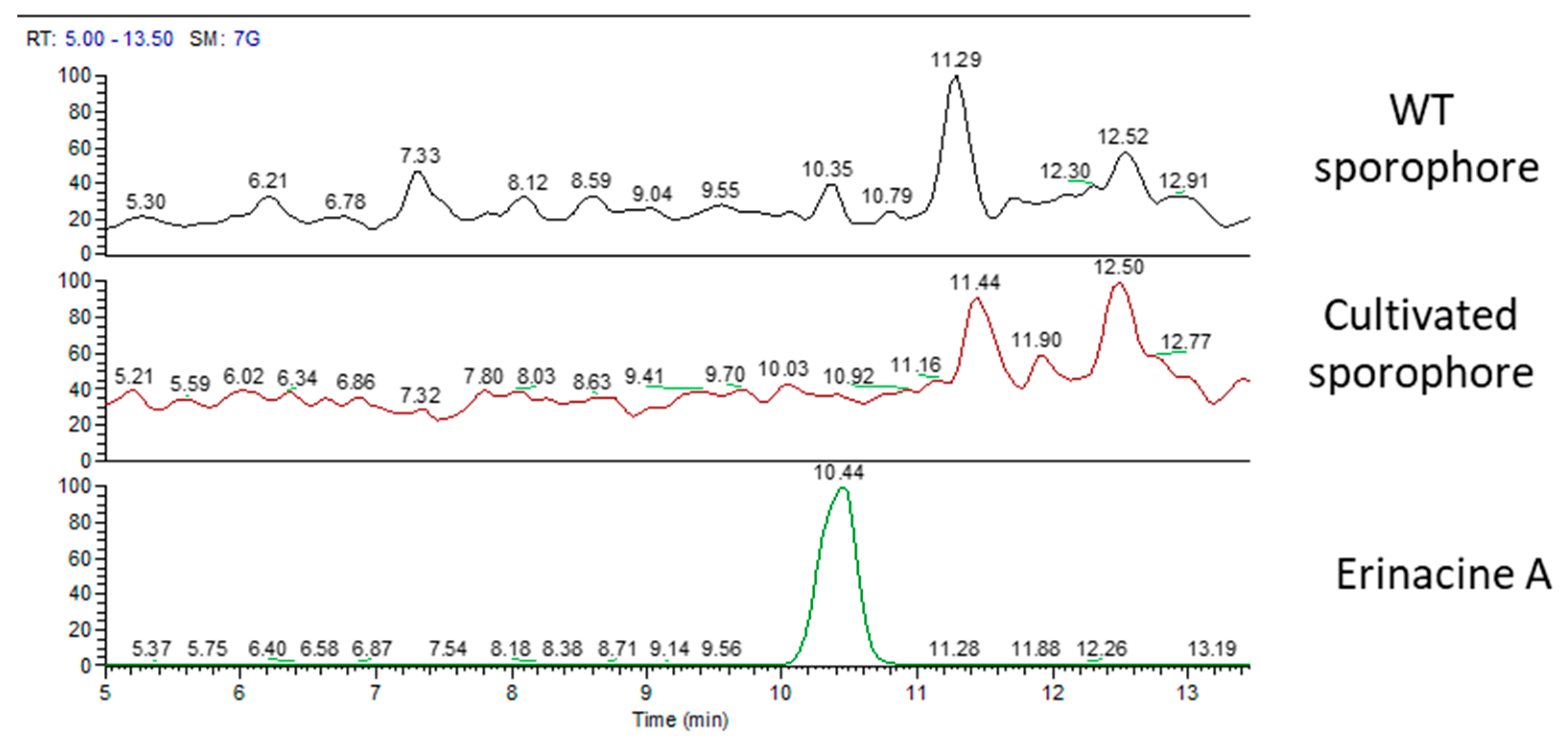
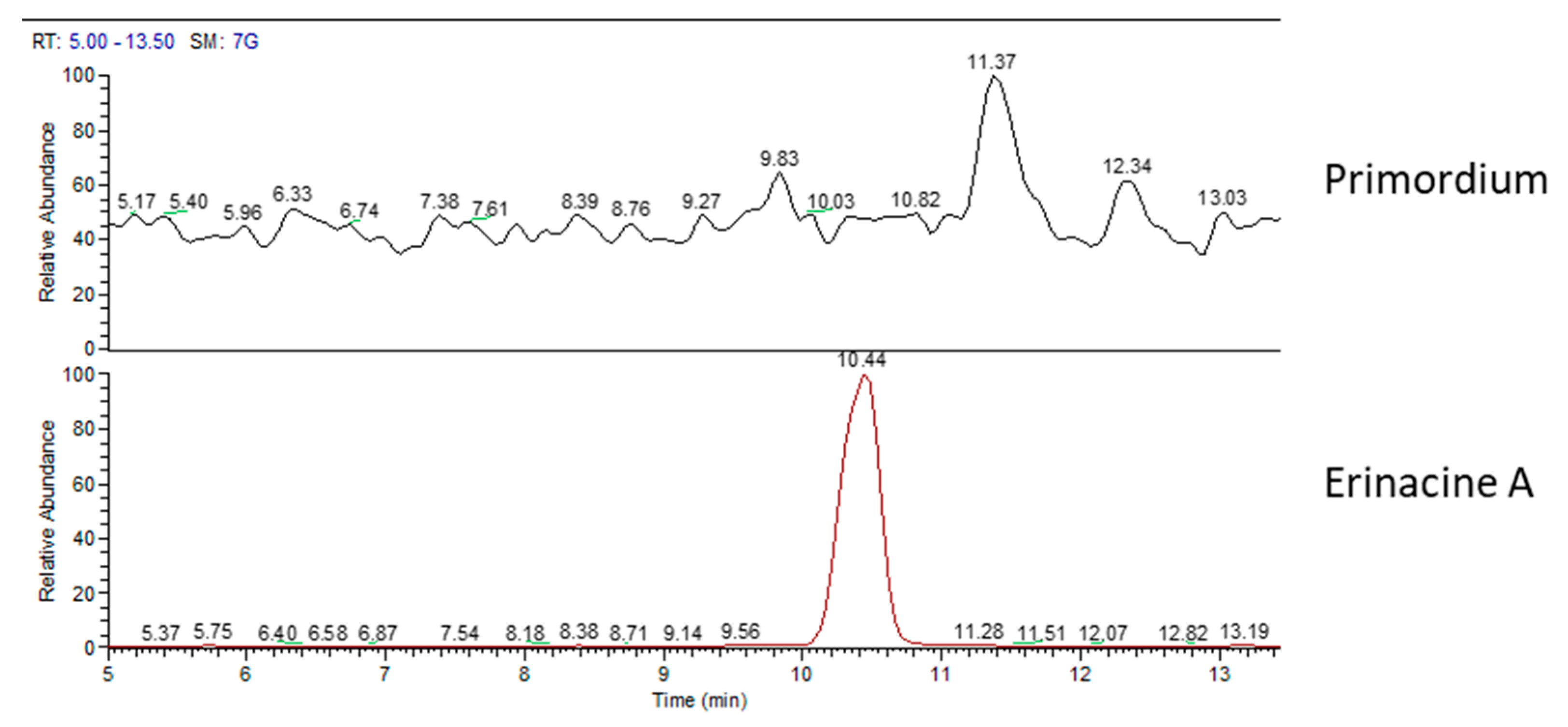
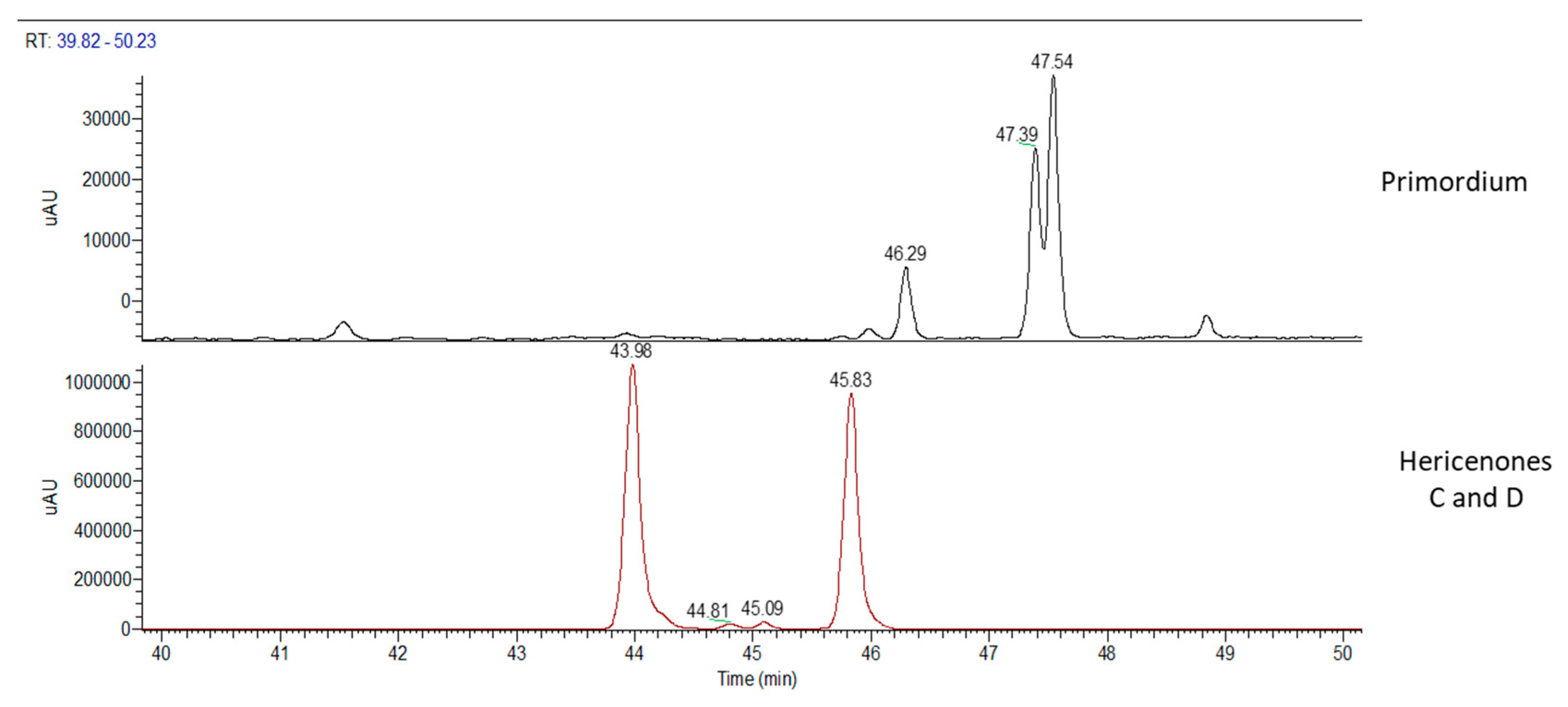
2.1. H. erinaceus Samples for Chemical Analyses
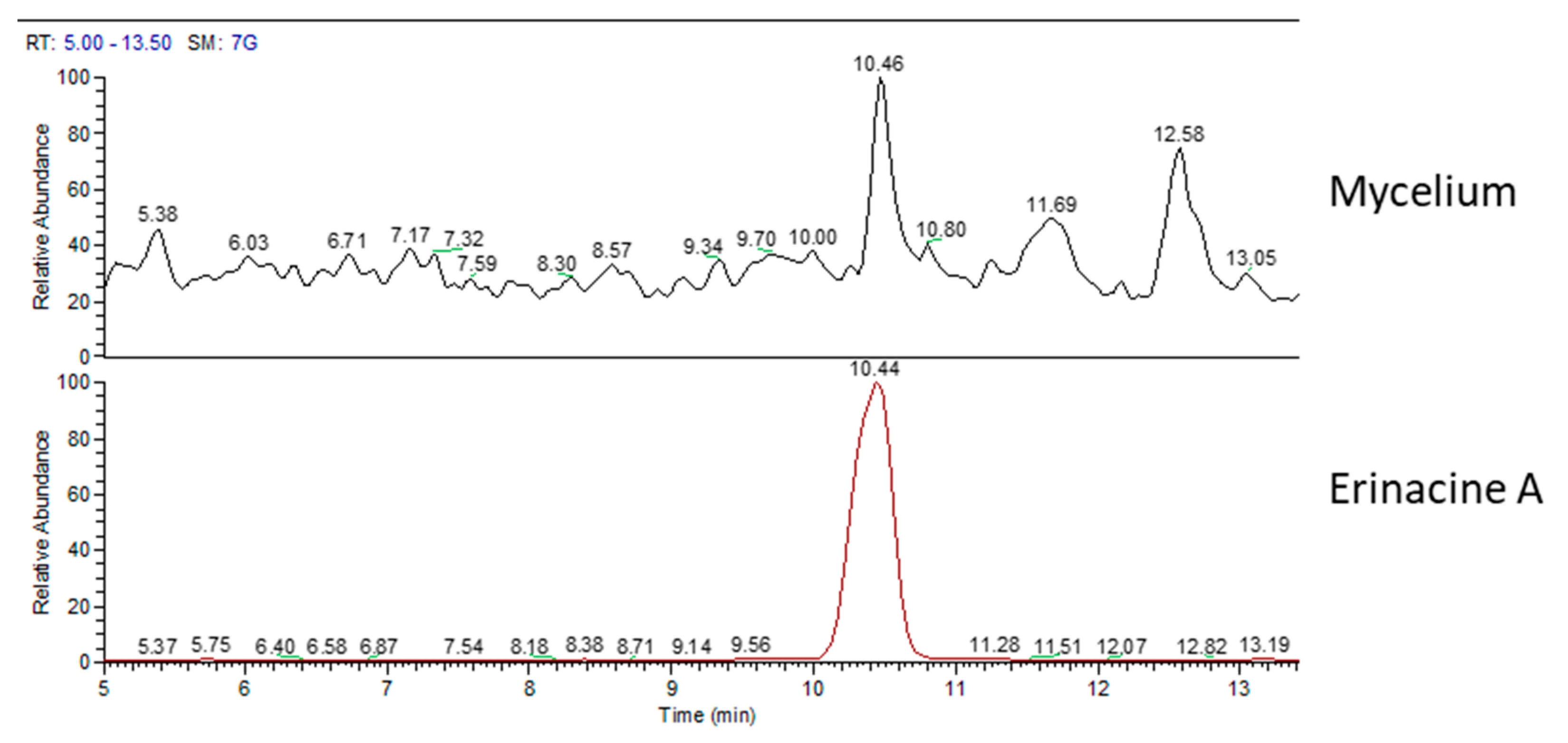
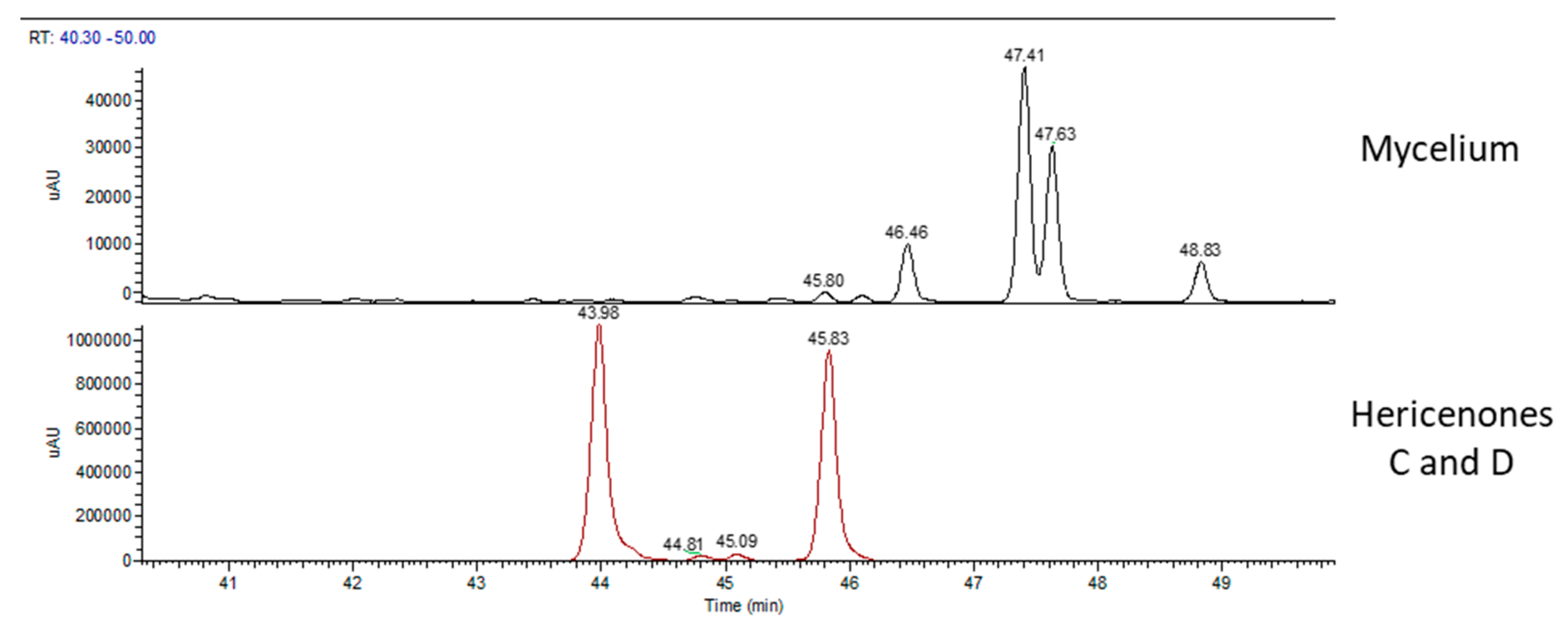
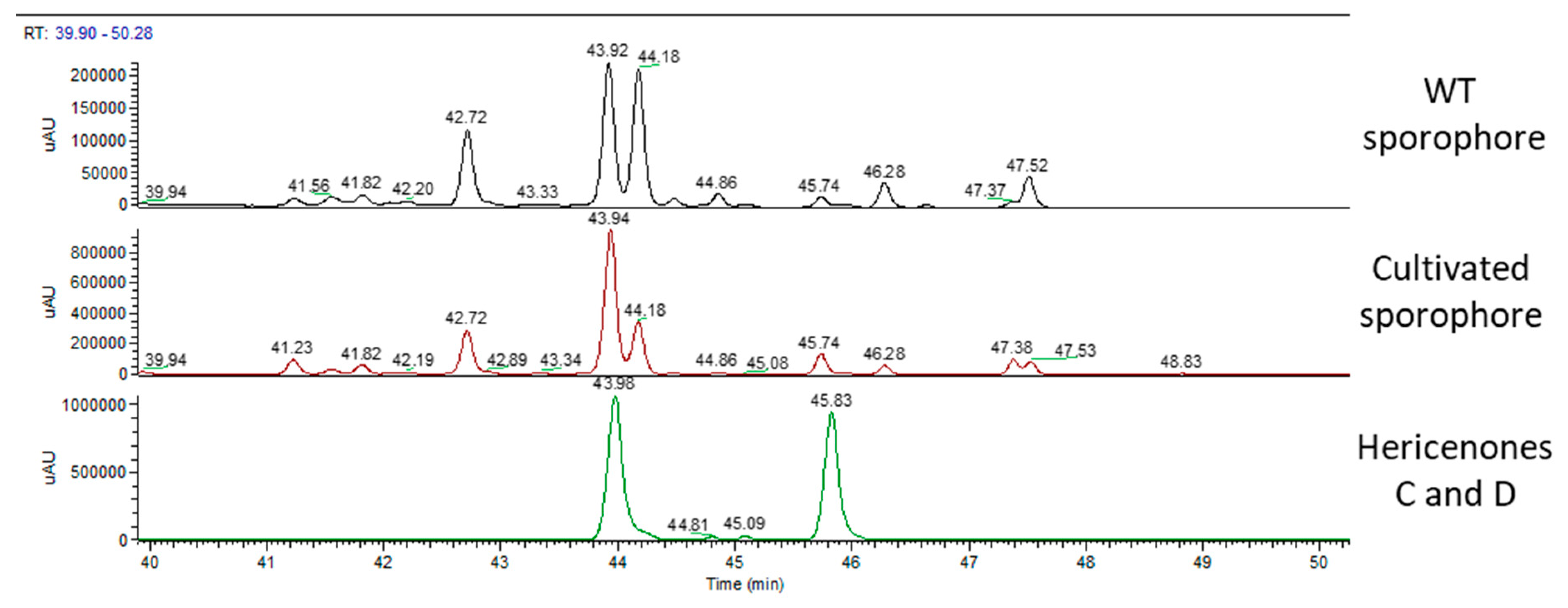
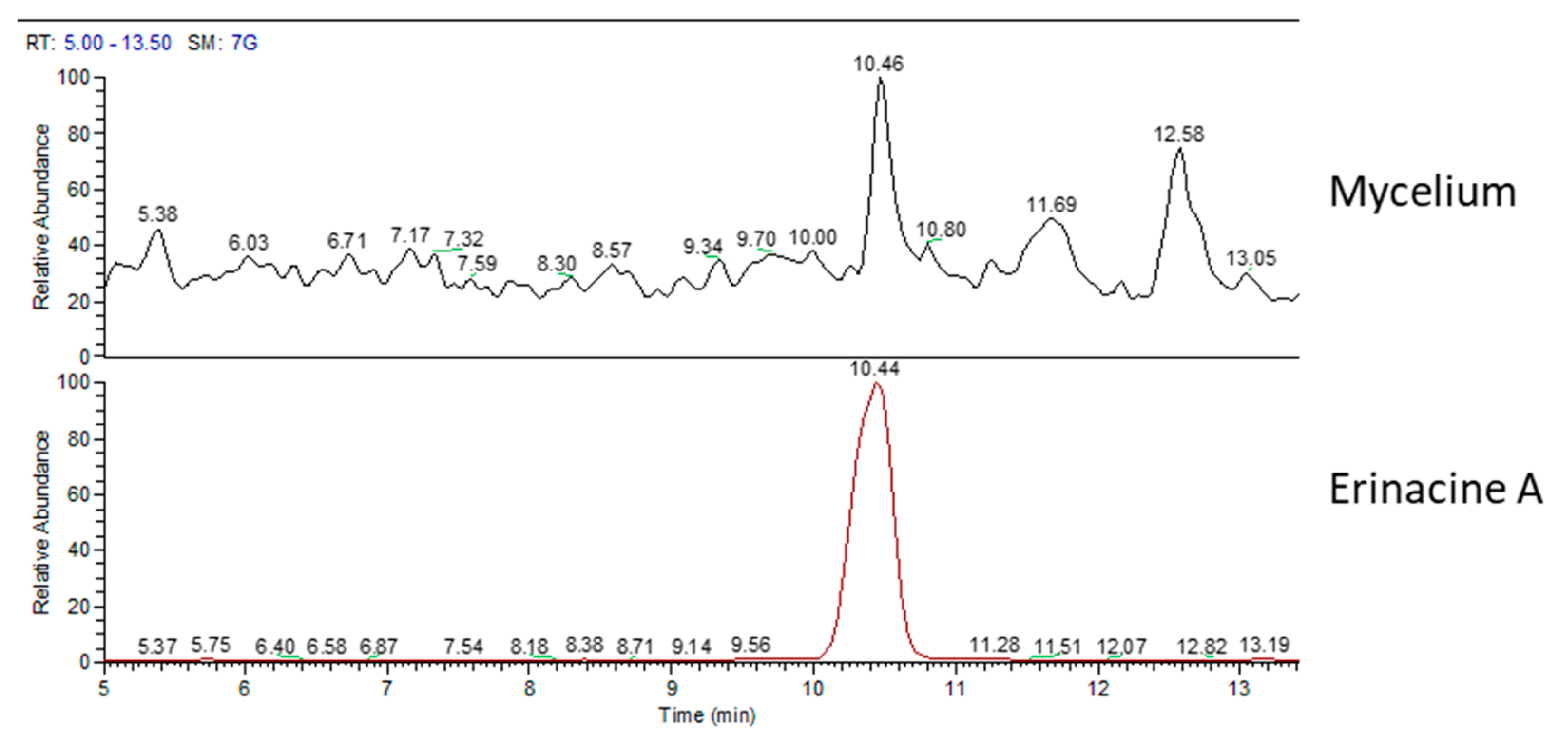
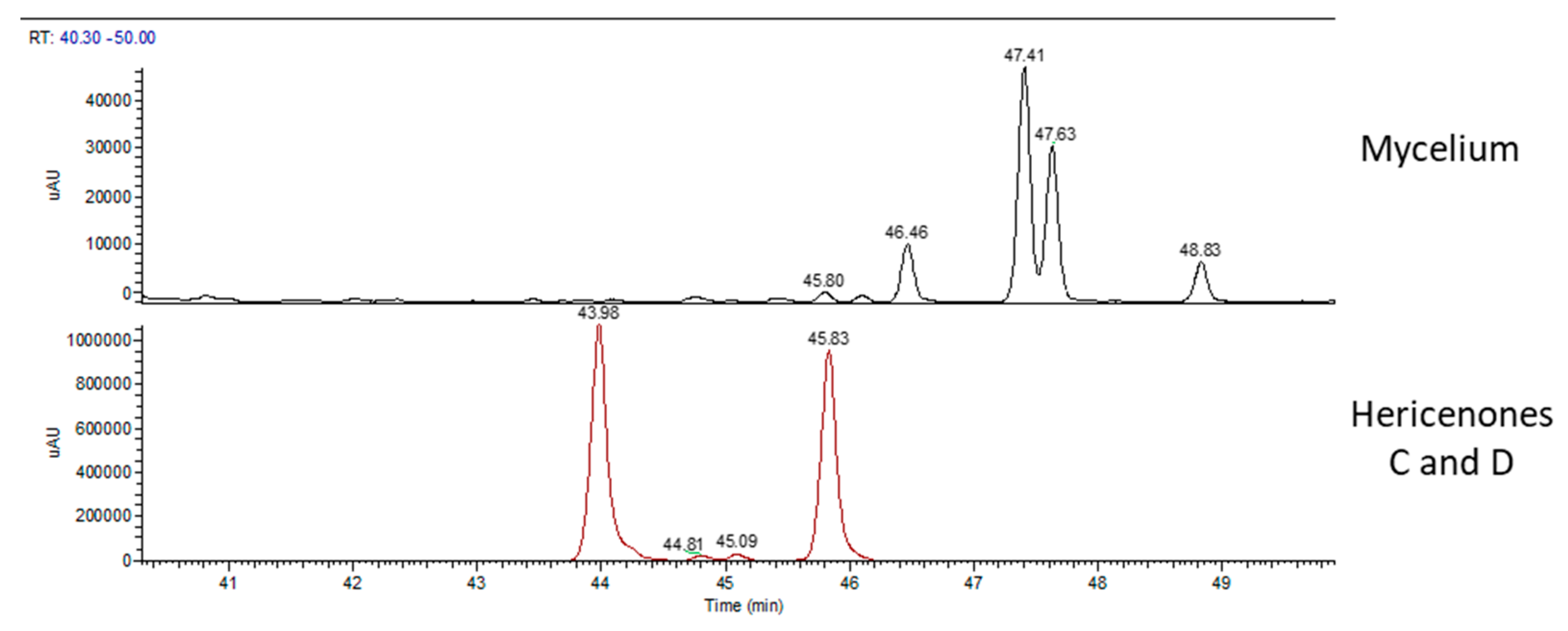
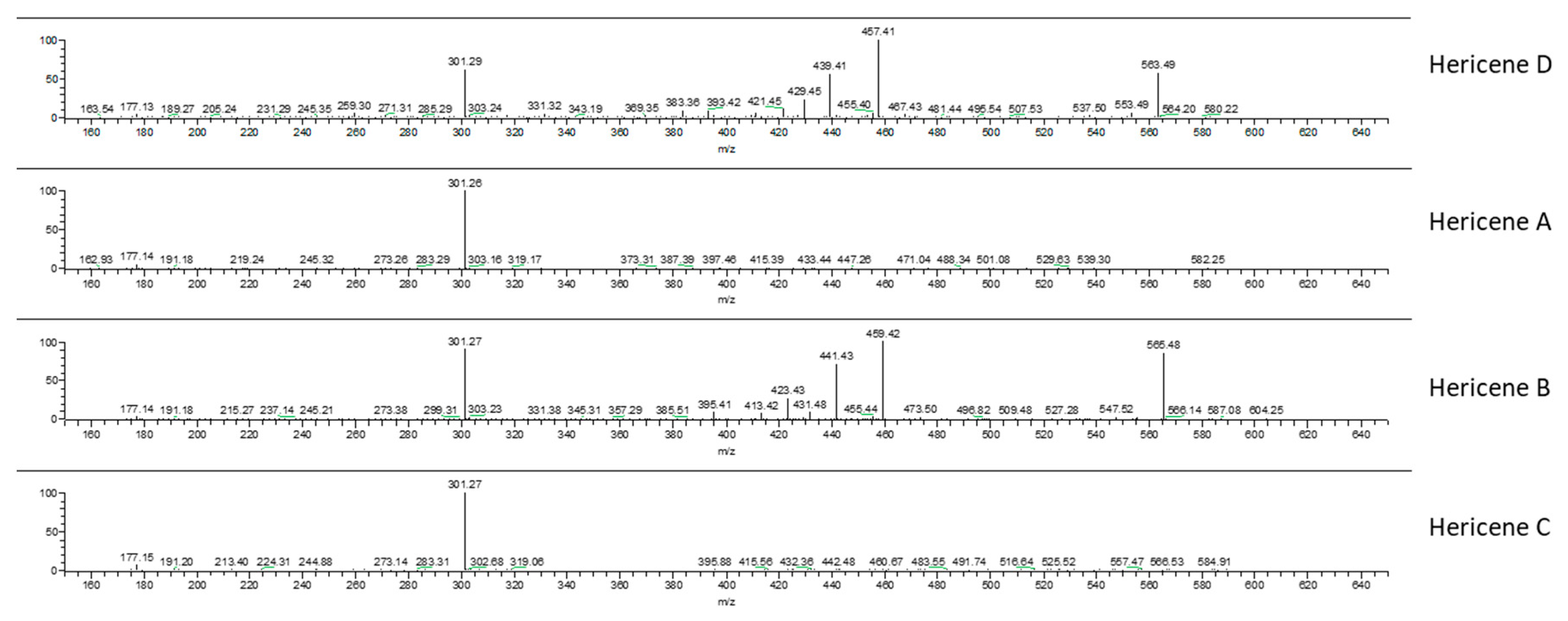
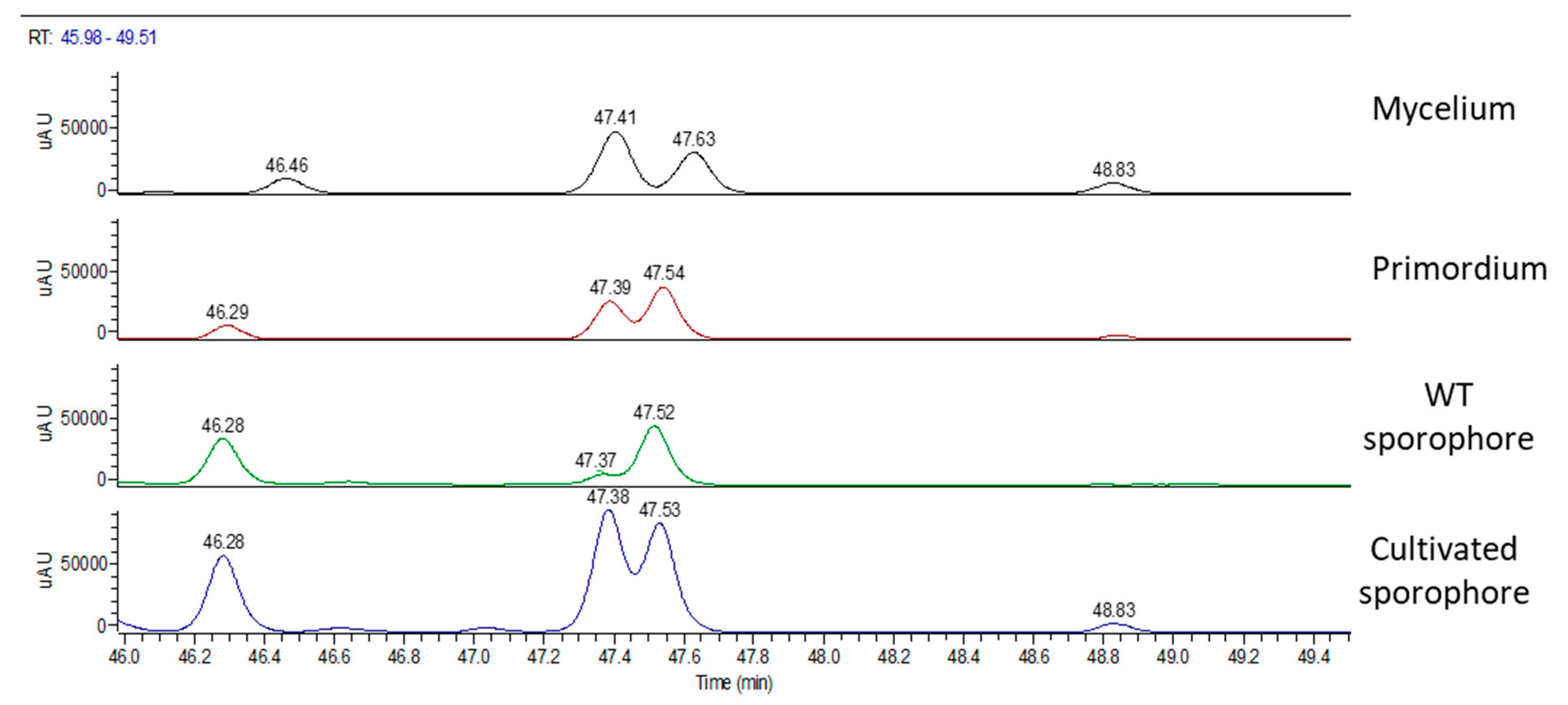
2.2. Chemical Analyses
3. Discussion
4. Materials and Methods
4.1. Study Area and Sampling
4.2. H. erinaceus Samples for Chemical Analyses
4.3. H. erinaceus Strain Isolation
4.4. H. erinaceus Sporophores Cultivation
4.5. Extraction Procedures
4.6. HPLC-UV-ESI/MS Method
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mycobank. Available online: www.mycobank.org (accessed on 24 September 2019).
- Rossi, P.; Cesaroni, V.; Brandalise, F.; Occhinegro, A.; Ratto, D.; Perrucci, F.; Lanaia, V.; Girometta, C.; Orrù, G.; Savino, E. Dietary Supplementation of Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Agaricomycetes), and Spatial Memory in Wild-Type Mice. Int. J. Med. Mushrooms 2018, 20, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Brandalise, F.; Cesaroni, V.; Gregori, A.; Repetti, M.; Romano, C.; Orrù, G.; Botta, L.; Girometta, C.; Guglielminetti, M.L.; Savino, E. Dietary Supplementation of Hericium erinaceus Increases Mossy Fiber-CA3 Hippocampal Neurotransmission and Recognition Memory in Wild-Type Mice. Evid. Based Complement. Alternat. Med. 2017, 3864340. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, G.; Ng, T.B.; Wang, H. A novel lectin with antiproliferative and HIV-1 reverse transcriptase inhibitory activities from dried fruiting bodies of the monkey head mushroom Hericium erinaceum. BioMed Res. Int. 2010, 2010, 716515. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhou, W.; Kim, E.J.; Shim, S.H.; Kang, H.K.; Kim, Y.H. Isolation and identification of aromatic compounds in Lion’s Mane Mushroom and their anticancer activities. Food Chem. 2015, 170, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Zan, X.; Cui, F.; Li, Y.; Yang, Y.; Wu, D.; Sun, W.; Ping, L. Hericium erinaceus polysaccharide-protein HEG-5 inhibits SGC-7901 cell growth via cell cycle arrest and apoptosis. Int. J. Biol. Macromol. 2015, 76, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Vigna, L.; Morelli, F.; Agnelli, G.M.; Napolitano, F.; Ratto, D.; Occhinegro, A.; Di Iorio, C.; Savino, E.; Girometta, C.; Brandalise, F.; et al. Hericium erinaceus Improves Mood and Sleep Disorders in Patients Affected by Overweight or Obesity: Could Circulating Pro-BDNF and BDNF Be Potential Biomarkers? Evid Based Complement Alternat Med. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [PubMed]
- De Silva, D.D.; Rapior, S.; Hyde, K.D.; Bahkali, A.H. Medicinal mushrooms in prevention and control of diabetes mellitus—A review. Fungal Divers. 2012, 56, 1–29. [Google Scholar] [CrossRef]
- Yang, B.K.; Park, J.B.; Song, C.H. Hypolipidemic Effect of an Exo-biopolymer Produced from a Submerged Mycelial Culture of Hericium erinaceus. Biosci. Biotech. Bioch. 2003, 67, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Ratto, D.; Corana, F.; Mannucci, B.; Priori, E.C.; Cobelli, F.; Roda, E.; Ferrari, B.; Occhinegro, A.; Di Iorio, C.; De Luca, F.; et al. Hericium erinaceus improves recognition memory and induces hippocampal and cerebellar neurogenesis in frail mice during aging. Nutrients 2019, 11, 715. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, nutrition, and health-promoting properties of Hericium erinaceus (Lion’s Mane) mushroom fruiting bodies and mycelia and their bioactive compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef]
- Kawagishi, H.; Shimada, A.; Shirai, R.; Okamoto, K.; Ojima, F.; Sakamoto, H.; Ishiguro, Y.; Furukawa, S. Erinacines A, B and C strong stimulators of nerve growth factor (NGF)-synthesis from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1994, 35, 1569–1572. [Google Scholar] [CrossRef]
- Ma, B.J.; Yu, H.Y.; Shen, J.W.; Ruan, Y.; Zhao, X.; Zhou, H.; Wu, T.T. Cytotoxic aromatic compunds from Hericium erinaceum. J. Antibiotics 2010, 63, 713–715. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, H.; Ando, M.; Mizuno, T. Hericenone A and B as cytotoxic principles from the mushroom Hericium erinaceum. Tetrahedron Lett. 1990, 31, 373–376. [Google Scholar] [CrossRef]
- Keong, C.Y.; Rashid, B.A.A.; Ing, Y.S.; Ismail, Z. Quantification and identification of polysaccharide contents in Hericium erinaceus. Nutr. Food Sci. 2007, 37, 260–271. [Google Scholar] [CrossRef]
- Ma, B.J.; Shen, J.W.; Yu, H.Y.; Ruan, Y.; Wu, T.T.; Zhao, X. Hericenones and erinacines: Stimulators of nerve growth factor (NGF) biosynthesis in Hericium erinaceus. Mycology 2010, 1, 2–92. [Google Scholar] [CrossRef]
- Kawagishi, H.; Ando, M.; Sakamoto, H.; Yoshida, S.; Ojima, F.; Ishiguro, Y.; Ukai, N.; Furukawa, S.C. Hericenones C, D and E, stimulators of nerve growth factor (NGF)-synthesis, from the mushroom Hericium erinaceum. Tetrahedron Lett. 1991, 32, 4561–4564. [Google Scholar] [CrossRef]
- Lee, D.G.; Kang, H.W.; Park, C.G.; A, Y.S.; Shin, Y. Isolation and identification of pyotochemicals and biological activities of Hericium erinaceus and their contents in Hericium strains using HPLC/UV analysis. J. Ethnopharmacol. 2016, 184, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.Y.; Yin, X.; Zhang, C.C.; Jia, Q.; Gao, J.M. Structure diversity, synthesis, and biological activity of cyathane diterpenoids in higher fungi. Curr. Med. Chem. 2015, 22, 2375–2391. [Google Scholar] [CrossRef]
- Kawagishi, H.; Shimada, A.; Hosokawa, S.; Mori, H.; Sakamoto, H.; Ishiguro, Y.; Sakemi, S.; Bordner, J.; Kojima, N.; Furukawa, S. Erinacines E, F, and G, stimulators of nerve growth factor (NGF) synthesis from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1996, 37, 7399–7402. [Google Scholar] [CrossRef]
- Kawagishi, H.; Simada, A.; Shizuki, K.; Mori, H.; Sakamoto, H.; Furukawa, S. Erinacine D, a stimulator of NGF-synthesis from the mycelia of Hericium erinaceum. Heterocycl Commun. 1996, 2, 51–54. [Google Scholar] [CrossRef]
- Lee, E.W.; Shizuki, K.; Hosokawa, S.; Suzuki, M.; Suganuma, H.; Inakuma, T.; Kawagishi, H. Two novel diterpenoids, erinacines H and I from the mycelia of Hericium erinaceum. Biosci. Biotech. Biochem. 2000, 64, 2402–2405. [Google Scholar] [CrossRef] [PubMed]
- Kenmoku, H.; Sassa, T.; Kato, N. Isolation of erinacine P, a new parental metabolite of cyathane-xylosides from Hericium erinaceus and its biomimetic conversion into erinacines A and B. Tetrahedron Lett. 2000, 41, 4389–4393. [Google Scholar] [CrossRef]
- Kenmoku, H.; Shimai, T.; Toyomasu, T.; Kato, N.; Sassa, T. Erinacine Q, a new erinacine from Hericium erinaceus, and its biosynthetic route to erinacine C in the basidiomycete. Biosci. Biotech. Biochem. 2002, 66, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, H.; Masui, A.; Tokuyamab, S.; Nakamurac, T. Erinacines J and K from the mycelia of Hericium erinaceum. Tetrahedron Lett. 2006, 62, 8463–8466. [Google Scholar] [CrossRef]
- Ma, B.J.; Zhou, Y.; Li, L.Z.; Li, H.M.; Gao, Z.M.; Ruan, Y. A new cyathanexyloside from the mycelia of Hericium erinaceum. Z Naturforsch 2008, 63b, 1241–1242. [Google Scholar] [CrossRef]
- Bernicchia, A.; Gorjon, S.P. Hericium erinaceus. In Corticiaceae s.l. Fungi europaei; Candusso: Alassio, Italy, 2010; Volume 12, pp. 318–319. ISBN 978-88-901057-9-8. [Google Scholar]
- Cesaroni, V.; Brusoni, M.; Cusaro, C.M.; Girometta, C.; Perini, C.; Picco, A.M.; Rossi, P.; Salerni, E.; Savino, E. Phylogenetic Comparison between Italian and Worldwide Hericium Species (Agaricomycetes). Int. J. Med. Mushrooms 2019, accepted. [Google Scholar]
- PubChem. Available online: http://pubchem.ncbi.nlm.nih.gov/ (accessed on 26 September 2019).
- Arnone, A.; Cardillo, R.; Nasini, G.; De Pava, O.V. Secondary mold metabolites: Part 46. Hericenes A-C and erinapyrone C, new metabolites produced by the fungus Hericium erinaceus. J. Nat. Prod. 1994, 57, 602–606. [Google Scholar] [CrossRef]
- Kobayashi, S.; Hamada, Y.; Yasumoto, T.; Hashino, Y.; Masuyama, A.; Nagai, K. Total syntheses and endoplasmic reticulum stress suppressive activities of hericenes A-C and their derivatives. Tetrahedron Lett. 2018, 59, 1733–1736. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal Mushroom Science: Current Perspectives, Advances, Evidences, and Challenges. Biomed. J. 2014, 37, 345–356. [Google Scholar] [CrossRef]
- Krzyczkowski, W.; Malinowska, E.; Herold, F. Erinacine A biosynthesis in submerged cultivation of Hericium erinaceum: Quantification and improved cultivation. Eng. Life Sci. 2010, 10, 446–457. [Google Scholar] [CrossRef]
- Zhang, G.; Sun, Z.; Ren, A.; Shi, L.; Shi, D.; Li, X.; Zhao, M. The mitogen-activated protein kinase GlSlt2 regulates fungal growth, fruiting body development, cell wall integrity, oxidative stress and ganoderic acid biosynthesis in Ganoderma lucidum. Fungal Genet. Biol. 2017, 104, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Gong, W.; Zhu, Z.; Yan, L.; Hu, Z.; Peng, Y. Comparative transcriptomics of Pleurotus eryngii reveals blue-light regulation of carbohydrate-active enzymes (CAZymes) expression at primordium differentiated into fruiting body stage. Genomics 2018, 110, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Stamets, P. The lion’s mane mushroom. In Growing Gourmet and Medicinal Mushrooms, 3rd ed.; Ten Speed Press: Berkeley, Toronto, 2000; pp. 387–394. ISBN 13 978-1-58008-175-7. [Google Scholar]
- Stalpers, J.A. Identification of wood-inhabiting Aphyllophorales in pure culture. Stud. Mycol. 1978, 16, 114. [Google Scholar]
- Sturini, M.; Girometta, C.; Maraschi, F.; Savino, E.; Profumo, A. A preliminary investigation on Metal Bioaccumulation by Perenniporia fraxinea. Bull. Environ. Contam. Toxicol. 2017, 98, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.G.; Park, H.G.; Park, S.H.; Choi, C.W.; Kim, S.H.; Park, W.M. Comparative study of mycelial growth and basidiomata formation in seven different species of the edible mushroom genus Hericium. Bioresource Technol. 2005, 96, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.R.H. Cultivation of the Monkey Head Mushroom (Hericium erinaceus) in Egypt. J. App. Sci. Res. 2007, 3, 1229–1233. [Google Scholar]
- Savino, E.; Girometta, C.; Baiguera, R.M.; Cesaroni, V.; Guglielminetti, M.L.; Rodolfi, M.; Rossi, P.; Picco, A.M. Different approaches for Hericium erinaceus spawn in the perspective of gluten free products. ISMS 2016, 19, 106. [Google Scholar]
- Gerbec, B.; Tavčar, E.; Gregori, A.; Kreft, S.; Berovic, M. Solid State Cultivation of Hericium erinaceus Biomass and Erinacine: A Production. J. Bioproces. Biotech. 2015, 5, 1–5. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from one of the authors (H. K.) under an MTA. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. erinaceus | |
|---|---|
| fresh weight (g) | 620 |
| dried weight (g) | 153 |
| diameter (cm) | about 20 |
| remarks on the sporophore | the collected specimen was mature, without any alteration by atmospheric or animal agents |
| Erinacine A | Hericenone C | Hericenone D | |
|---|---|---|---|
| molecular formula | C25H36O6 | C35H54O6 | C37H58O6 |
| molecular weight (MW) (g/mol) | 432 | 570 | 598 |
| chemical structure | 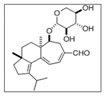 | 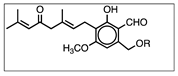 R = palmitoyl | 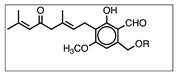 R = stearoyl |
| Hericene A | Hericene B | Hericene C | Hericene D | |
|---|---|---|---|---|
| molecular formula | C35H56O5 | C37H58O5 | C37H60O5 | C37H56O5 |
| molecular weight (MW) (g/mol) | 556 | 582 | 584 | 580 |
| chemical structure |  | |||
| R = palmitoyl | R = oleoyl | R = stearoyl | R = lineoyl | |
| erinacine A | hericenone C | hericenone D | hericenes | |
|---|---|---|---|---|
| mycelium | ✓ | - | - | ✓ |
| primordium | - | - | - | ✓ |
| WT sporophore | - | ✓ | ✓ | ✓ |
| cultivated sporophore | - | ✓ | ✓ | ✓ |
| Erinacine A (µg/g) | Hericenone C (µg/g) | Hericenone D (µg/g) | |
|---|---|---|---|
| mycelium | 105 | - | - |
| primordium | - | - | - |
| WT sporophore | - | 760 | 100 |
| cultivated sporophore | - | 1560 | 188 |
| Total Area 103 | Hericene A Area 103 | Hericene B Area 103 | Hericene C Area 103 | Hericene D Area 103 | |
|---|---|---|---|---|---|
| mycelium | 684 | 327 | 232 | 51 | 74 |
| primordium | 557 | 201 | 262 | 21 | 73 |
| WT sporophore | 627 | 70 | 305 | / | 252 |
| cultivated sporophore | 1685 | 645 | 588 | 42 | 410 |
| Time | Solvent A | Solvent B |
|---|---|---|
| 0 | 70 | 30 |
| 9 | 50 | 50 |
| 27 | 40 | 60 |
| 54 | 00 | 100 |
| 69 | 70 | 30 |
| 75 | 70 | 30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corana, F.; Cesaroni, V.; Mannucci, B.; Baiguera, R.M.; Picco, A.M.; Savino, E.; Ratto, D.; Perini, C.; Kawagishi, H.; Girometta, C.E.; et al. Array of Metabolites in Italian Hericium erinaceus Mycelium, Primordium, and Sporophore. Molecules 2019, 24, 3511. https://doi.org/10.3390/molecules24193511
Corana F, Cesaroni V, Mannucci B, Baiguera RM, Picco AM, Savino E, Ratto D, Perini C, Kawagishi H, Girometta CE, et al. Array of Metabolites in Italian Hericium erinaceus Mycelium, Primordium, and Sporophore. Molecules. 2019; 24(19):3511. https://doi.org/10.3390/molecules24193511
Chicago/Turabian StyleCorana, Federica, Valentina Cesaroni, Barbara Mannucci, Rebecca Michela Baiguera, Anna Maria Picco, Elena Savino, Daniela Ratto, Claudia Perini, Hirokazu Kawagishi, Carolina Elena Girometta, and et al. 2019. "Array of Metabolites in Italian Hericium erinaceus Mycelium, Primordium, and Sporophore" Molecules 24, no. 19: 3511. https://doi.org/10.3390/molecules24193511
APA StyleCorana, F., Cesaroni, V., Mannucci, B., Baiguera, R. M., Picco, A. M., Savino, E., Ratto, D., Perini, C., Kawagishi, H., Girometta, C. E., & Rossi, P. (2019). Array of Metabolites in Italian Hericium erinaceus Mycelium, Primordium, and Sporophore. Molecules, 24(19), 3511. https://doi.org/10.3390/molecules24193511