
Selective Cannabinoid 2 Receptor Agonists as Potential Therapeutic Drugs for the Treatment of Endotoxin-Induced Uveitis

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Chemical Structure
2.2. In Vitro Pharmacology of CB2R Ligands
2.3. Early Absorption, Distribution, Metabolism, Excretion, and Toxicology Profile
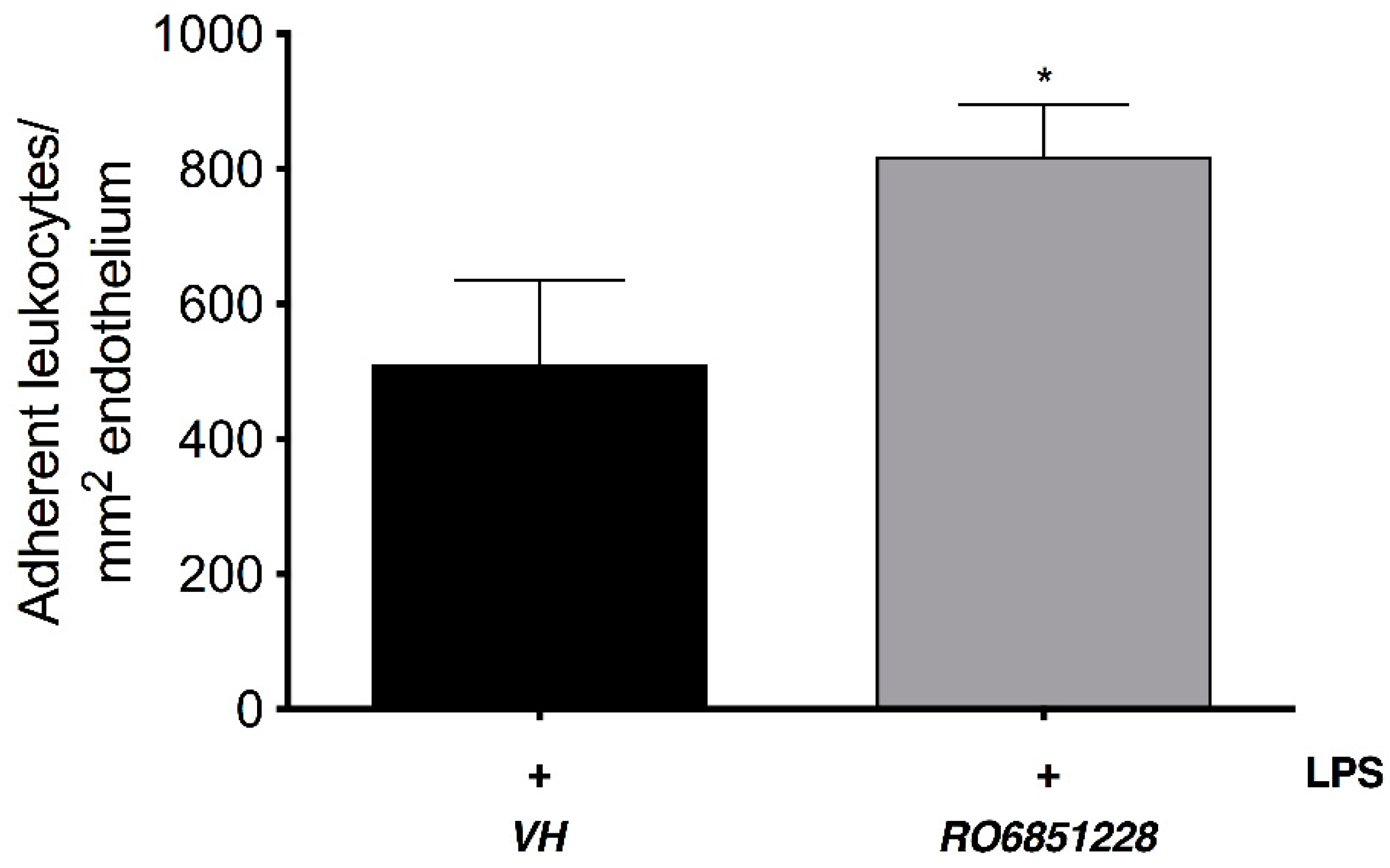
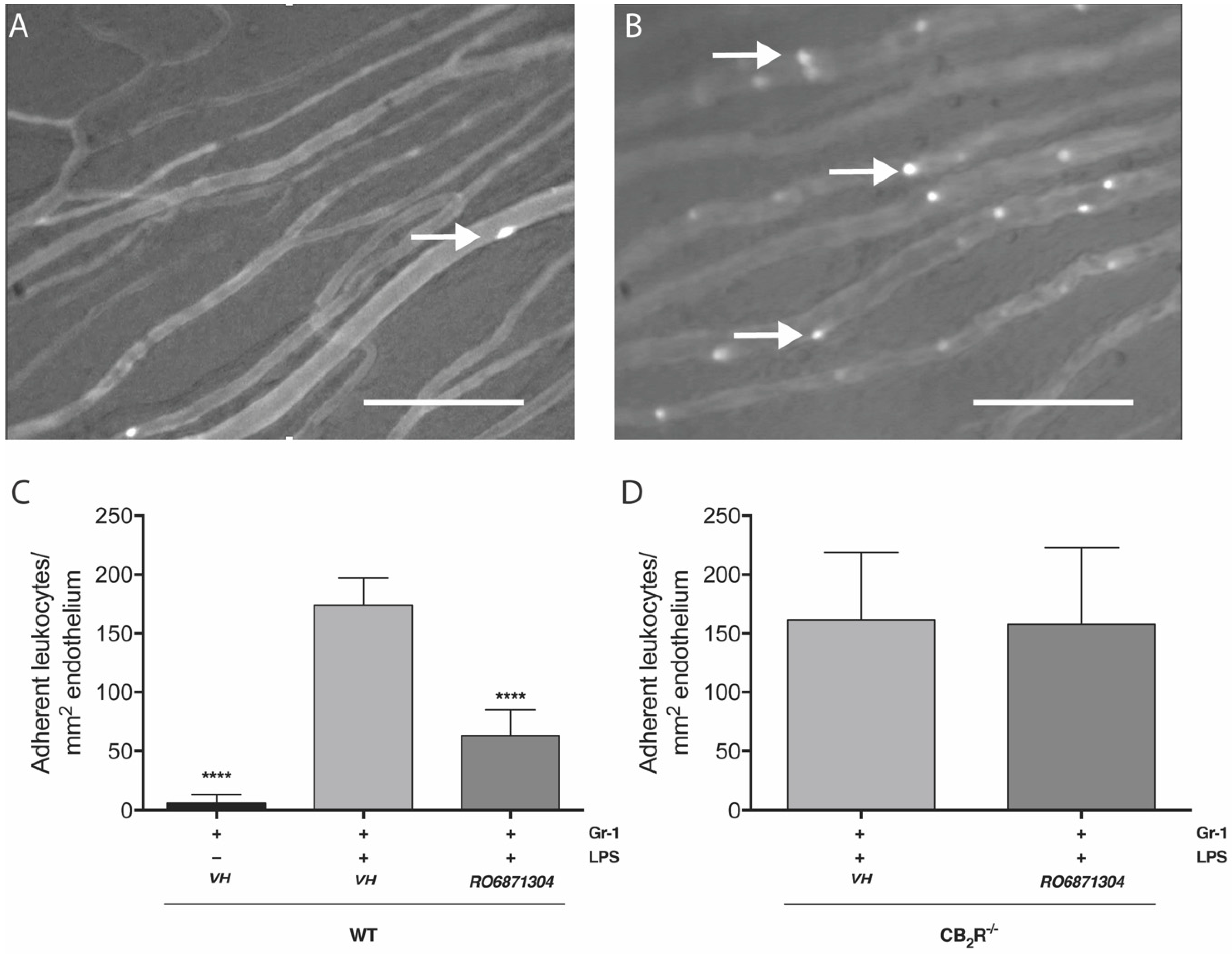
2.4. Immunomodulatory Properties of CB2R-Selective Ligands in EIU
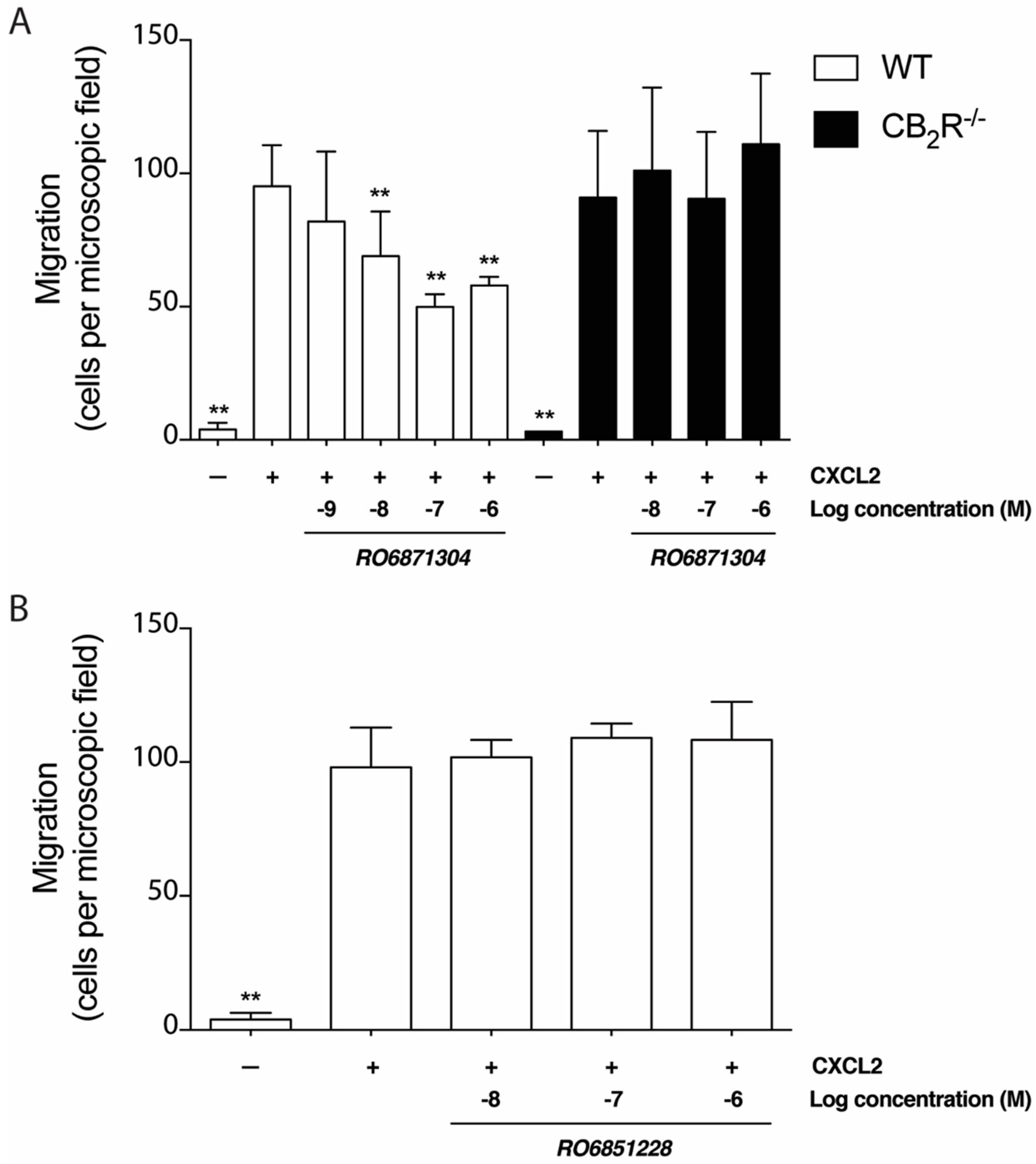
2.5. Effects of CB2R-Selective Ligands on Neutrophil Migration In Vitro
2.6. Adoptive Transfer
CB2R Activation on Resident Immune Cells Mediates CB2R Leukocyte Adherence Effects
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Membrane Preparations
4.2. Mouse CB2R and Human CB2R and CB1R Binding Assays
4.3. PathHunter β-Arrestin Recruitment Assays
4.4. cAMP Assays
4.5. Animals
4.6. Endotoxin-Induced Uveitis (EIU) Model
4.7. Intravital Microscopy (IVM)
4.8. Isolation of Murine Bone Marrow Cell Neutrophils
4.9. Neutrophil Migration Assay
4.10. Neutrophil-Depletion and Adoptive Transfer
4.11. Peripheral Blood Leukocyte Isolation
4.12. Antibodies and Flow Cytometry
4.13. Drug Treatments
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guly, C.M.; Forrester, J.V. Investigation and management of uveitis. BMJ 2010, 341, c4976. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.C.S.; Subramanyam, S.; Ramana, K.V. Prevention of Endotoxin-Induced Uveitis in Rats by Benfotiamine, a Lipophilic Analogue of Vitamin B1. Invest. Ophthalmol. Vis. Sci. 2009, 50, 2276–2282. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lu, H.; Hu, X.; Chen, W.; Xu, Y.; Wang, J. Expression of TLR4-MyD88 and NF-κB in the iris during endotoxin-induced uveitis. Mediators Inflamm. 2010, 2010, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Barry, R.J.; Nguyen, Q.D.; Wlee, R.; Imurray, P.; Denniston, A.K. Pharmacotherapy for uveitis: Current management and emerging therapy. Clin. Ophthalmol. 2014, 8, 1891–1911. [Google Scholar] [PubMed]
- Kersey, J.P.; Broadway, D.C. Corticosteroid-induced glaucoma: A review of the literature. Eye 2006, 20, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Batkai, S.; Kunos, G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef]
- Munoz-Luque, J.; Ros, J.; Fernandez-Varo, G.; Tugues, S.; Morales-Ruiz, M.; Alvarez, C.E.; Friedman, S.L.; Arroyo, V.; Jimenez, W. Regression of fibrosis after chronic stimulation of cannabinoid CB2 receptor in cirrhotic rats. J. Pharmacol. Exp. Ther. 2007, 324, 475–483. [Google Scholar] [CrossRef]
- Rajesh, M.; Mukhopadhyay, P.; Bátkai, S.; Patel, V.; Saito, K.; Matsumoto, S.; Kashiwaya, Y.; Horváth, B.; Mukhopadhyay, B.; Becker, L.; et al. Cannabidiol attenuates cardiac dysfunction, oxidative stress, fibrosis, and inflammatory and cell death signaling pathways in diabetic cardiomyopathy. J. Am. Coll. Cardiol. 2010, 56, 2115–2125. [Google Scholar] [CrossRef]
- Kreitzer, F.R.; Stella, N. The therapeutic potential of novel cannabinoid receptors. Pharmacol. Ther. 2009, 122, 83–96. [Google Scholar] [CrossRef]
- Mechoulam, R.; Parker, L.A. The endocannabinoid system and the brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M.; Duncan, M.; Di Marzo, V.; Pertwee, R. Are cannabidiol and Δ(9)-tetrahydrocannabivarin negative modulators of the endocannabinoid system? A systematic review. Br. J. Pharmacol. 2015, 172, 737–753. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of pheripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Cairns, E.A.; Toguri, T.J.; Porter, R.F.; Szczesniak, A.M.; Kelly, M.E.M. Seeing over the horizon: Targeting the endocannabiond system for the treatment of ocular disease. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Rom, S.; Zuluaga-Ramirez, V.; Dykstra, H.; Reichenbach, N.L.; Pacher, P.; Persidsky, Y. Selective activation of cannabinoid receptor 2 in leukocytes suppresses their engagement of the brain endothelium and protects the blood-brain barrier. Am. J. Pathol. 2013, 183, 1548–1558. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tanasescu, R.; Constantinescu, C.S. Cannabinoids and the immune system: An overview. Immunobiology 2010, 215, 588–597. [Google Scholar] [CrossRef]
- Maresz, K.; Carrier, E.J.; Ponomarev, E.D.; Hillard, C.J.; Dittel, B.N. Modulation of the cannabinoid CB2 receptor in microglial cells in response to inflammatory stimuli. J. Neurochem. 2005, 95, 437–445. [Google Scholar] [CrossRef]
- Storr, M.A.; Keenan, C.M.; Zhang, H.; Patel, K.D.; Makriyannis, A.; Sharkey, K.A. Activation of the cannabinoid 2 receptor (CB2) protects against experimental colitis. Inflamm. Bowel Dis. 2009, 15, 1678–1685. [Google Scholar] [CrossRef]
- Horváth, B.; Magid, L.; Mukhopadhyay, P.; Bátkai, S.; Rajesh, M.; Park, O.; Tanchian, G.; Gao, R.Y.; Goodfellow, C.E.; Glass, M.; et al. A new cannabinoid CB2 receptor agonist HU-910 attenuates oxidative stress, inflammation and cell death associated with hepatic ischaemia/reperfusion injury. Br. J. Pharmacol. 2012, 165, 2462–2478. [Google Scholar] [CrossRef]
- Di Marzo, V. Targeting the endocannabinoid system: To enhance or reduce? Nat. Rev. Drug Discov. 2008, 7, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Szczesniak, A.; Porter, R.; Toguri, J.; Borowska-Fielding, J.; Siwakoti, A.; Lehmann, C.; Kelly, M. Cannabinoid 2 receptor is a novel anti-inflammatory target in experimental proliferative vitreoretinopathy. Neuropharmcology 2017, 113, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Toguri, J.T.; Lehmann, C.; Laprairie, R.B.; Szczesniak, A.M.; Zhou, J.; Denovan-Wright, E.M.; Kelly, M.E.M. Anti-inflammatory effects of cannabinoid CB2 receptor activation in endotoxin-induced uveitis. Br. J. Pharmacol. 2014, 171, 1448–1461. [Google Scholar] [CrossRef] [PubMed]
- Toguri, T.; Leishman, E.; Szczesniak, A.; Laprairie, R.; Oehler, O.; Straiker, A.; Kelly, M.E.M.; Bradshaw, H.B. Inflammation and CB2 signaling drive novel changes in the ocular lipidome and regulate immune cell activity in the eye. Prostaglandins Other Lipid Mediat. 2018, 139, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-C.; Mackie, K. An introduction to the endogenous cannabinoid system. Biol. Psychiatry 2016, 59, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, P.; Di Giacomo, D.; Geremia, R. The endocannabinoid system and spermatogenesis. Front. Endocrinol. (Lausanne). 2013, 4. [Google Scholar] [CrossRef]
- Pertwee, R.G. Targeting the endocannabinoid system with cannabinoid receptor agonists: Pharmacological strategies and therapeutic possibilities. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3353–3363. [Google Scholar] [CrossRef]
- Soethoudt, M.; Grether, U.; Fingerle, J.; Grim, T.W.; Fezza, F.; de Petrocellis, L.; Ullmer, C.; Rothenhäusler, B.; Perret, C.; van Gils, N.; et al. Cannabinoid CB2 receptor ligand profiling reveals biased signalling and off-target activity. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Bissantz, C.; Grether, U.; Kimbara, A.; Nettekoven, M.; Roever, S.; Rogers-Evans, M. Preparation of [1,2,3]triazolo [4,5-d]pyrimidine derivatives useful as cannabinoid receptor 2 agonists. WO 2013076182 A1, 30 May 2013. [Google Scholar]
- Grether, U.; Kimbara, A.; Nettekoven, M.; Ricklin, F.; Roever, S.; Rogers-Evans, M.; Rombach, D.; Schulz-Gasch, T.; Westphal, M. Pyridine-2-amides useful as CB2 agonists. WO 2014086806 A1, 12 June 2014. [Google Scholar]
- Frei, B.; Gobbi, L.; Grether, U.; Ricklin, F.; Roever, S.; Rogers-Evans, M.; Rombach, D. Pyridine derivatives. WO 2018234284 A1, 27 December 2018. [Google Scholar]
- Ouali Alami, N.; Schurr, C.; Olde Heuvel, F.; Tang, L.; Li, Q.; Tasdogan, A.; Kimbara, A.; Nettekoven, M.; Ottaviani, G.; Raposo, C.; et al. NF-κB activation in astrocytes drives a stage-specific beneficial neuroimmunological response in ALS. EMBO J. 2018, e98697. [Google Scholar] [CrossRef] [PubMed]
- Nettekoven, M.; Adam, J.M.; Bendels, S.; Bissantz, C.; Fingerle, J.; Grether, U.; Grüner, S.; Guba, W.; Kimbara, A.; Ottaviani, G.; et al. Novel triazolopyrimidine-derived cannabinoid receptor 2 agonists as potential treatment for inflammatory kidney diseases. Chem. Med. Chem. 2016, 11, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Hanus, L.; Breuer, A.; Tchilibon, S.; Shiloah, S.; Goldenberg, D.; Horowitz, M.; Pertwee, R.G.; Ross, R.A.; Mechoulam, R.; Fride, E. HU-308: A specific agonist for CB(2), a peripheral cannabinoid receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 14228–14233. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hua, T.; Vemuri, K.; Ho, J.H.; Wu, Y.; Wu, L.; Popov, P.; Benchama, O.; Zvonok, N.; Locke, K.; et al. Crystal Structure of the Human Cannabinoid Receptor CB2. Cell 2019, 176, 459–467.e13. [Google Scholar] [CrossRef]
- Lorenzen, E.; Sakmar, T.P. Receptor Structures for a Caldron of Cannabinoids. Cell 2019, 176, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Ullmer, C.; Zoffmann, S.; Bohrmann, B.; Matile, H.; Lindemann, L.; Flor, P.J.; Malherbe, P. Functional monoclonal antibody acts as a biased agonist by inducing internalization of metabotropic glutamate receptor 7. Br. J. Pharmacol. 2012, 167, 1448–1466. [Google Scholar] [CrossRef]
- Di, L.; Rong, H.; Feng, B. Demystifying brain penetration in central nervous system drug discovery. J. Med. Chem. 2013, 56, 2–12. [Google Scholar] [CrossRef]
- Honer, M.; Gobbi, L.; Martarello, L.; Comley, R.A. Radioligand development for molecular imaging of the central nervous system with positron emission tomography. Drug Discov. Today 2014, 19, 1936–1944. [Google Scholar] [CrossRef]
- Kansy, M.; Senner, F.; Gubernator, K. Physicochemical high throughput screening: Parallel artificial membrane permeation assay in the description of passive absorption processes. J. Med. Chem. 1998, 41, 1007–1010. [Google Scholar] [CrossRef]
- De Filippo, K.; Dudeck, A.; Hasenberg, M.; Nye, E.; van Rooijen, N.; Hartmann, K.; Gunzer, M.; Roers, A.; Hogg, N. Mast cell and macrophage chemokines CXCL1/CXCL2 control the early stage of neutrophil recruitment during tissue inflammation. Blood 2013, 121, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.D.; Nobiling, R.; Planck, S.R.; Rosenbaum, J.T. Digital video-imaging of leukocyte migration in the iris: Intravital microscopy in a physiological model during the onset of endotoxin-induced uveitis. J. Immunol. Methods 2000, 240, 23–37. [Google Scholar] [CrossRef]
- McHugh, D.; Tanner, C.; Mechoulam, R.; Pertwee, R.G.; Ross, R.A. Inhibition of human neutrophil chemotaxis by endogenous cannabinoids and phytocannabinoids: Evidence for a site distinct from CB1 and CB2. Mol. Pharmacol. 2008, 73, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Dubald, M.; Bourgeois, S.; Andrieu, V.; Fessi, H. Ophthalmic drug delivery systems for antibiotherapy- A review. Pharmaceutics 2018, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Benito, C.; Núñez, E.; Tolón, R.M.; Carrier, E.J.; Rábano, A.; Hillard, C.J.; Romero, J. Cannabinoid CB2 receptors and fatty acid amide hydrolase are selectively overexpressed in neuritic plaque-associated glia in Alzheimer’s disease brains. J. Neurosci. 2003, 23, 11136–11141. [Google Scholar] [CrossRef] [PubMed]
- Carrier, E.J.; Kearn, C.S.; Barkmeier, A.J.; Breese, N.M.; Yang, W.; Nithipatikom, K.; Pfister, S.L.; Campbell, W.B.; Hillard, C.J. Cultured rat microglial cells synthesize the endocannabinoid 2-arachidonylglycerol, which increases proliferation via a CB2 receptor-dependent mechanism. Mol. Pharmacol. 2004, 65, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Eljaschewitsch, E.; Witting, A.; Mawrin, C.; Lee, T.; Schmidt, P.M.; Wolf, S.; Hoertnagl, H.; Raine, C.S.; Schneider-Stock, R.; Nitsch, R.; et al. The endocannabinoid anandamide protects neurons during CNS inflammation by induction of MKP-1 in microglial cells. Neuron 2006, 49, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, B.G. Prevention of Alzheimer’s disease pathology by cannabinoids: Neuroprotection mediated by blockade of microglial activation. J. Neurosci. 2005, 25, 1904–1913. [Google Scholar] [CrossRef]
- Sardinha, J.; Kelly, M.E.M.; Zhou, J.; Lehmann, C. Experimental cannabinoid 2 receptor-mediated immune modulation in sepsis. Mediators Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Lehmann, C.; Kianian, M.; Zhou, J.; Küster, I.; Kuschnereit, R.; Whynot, S.; Hung, O.; Shukla, R.; Johnston, B.; Cerny, V.; et al. Cannabinoid receptor 2 activation reduces intestinal leukocyte recruitment and systemic inflammatory mediator release in acute experimental sepsis. Crit. Care 2012, 16, 1–22. [Google Scholar] [CrossRef]
- Xu, H.; Cheng, C.L.; Chen, M.; Manivannan, A.; Cabay, L.; Pertwee, R.G.; Coutts, A.; Forrester, J. V Anti-inflammatory property of the cannabinoid receptor-2-selective agonist JWH-133 in a rodent model of autoimmune uveoretinitis. J. Leukoc. Biol. 2007, 82, 532–541. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.C.; Drummond, G.B.; McLachlan, E.M.; Kilkenny, C.; Wainwright, C.L. Editorial: Guidelines for reporting experiments involving animals: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1573–1576. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Kezic, J.; Taylor, S.; Gupta, S.; Planck, S.R.; Rosenzweig, H.L.; Rosenbaum, J.T. Endotoxin-induced uveitis is primarily dependent on radiation-resistant cells and on MyD88 but not TRIF. J. Leukoc. Biol. 2011, 90, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Forster, C.; Nakamura, F.; Glogauer, M. Filamin-A regulates neutrophil uropod retraction through RhoA during chemotaxis. PLoS ONE 2013, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tole, S.; Mukovozov, I.M.; Huang, Y.-W.; Magalhaes, M.A.O.; Yan, M.; Crow, M.R.; Liu, G.Y.; Sun, C.X.; Durocher, Y.; Glogauer, M.; et al. The axonal repellent, Slit2, inhibits directional migration of circulating neutrophils. J. Leukoc. Biol. 2009, 86, 1403–1415. [Google Scholar] [CrossRef]
- Sieve, A.N.; Meeks, K.D.; Bodhankar, S.; Lee, S.; Kolls, J.K.; Simecka, J.W.; Berg, R.E. A novel IL-17-dependent mechanism of cross protection: Respiratory infection with mycoplasma protects against a secondary listeria infection. Eur. J. Immunol. 2009, 39, 426–438. [Google Scholar] [CrossRef][Green Version]
- Daley, J.M.; Thomay, A.A.; Connolly, M.D.; Reichner, J.S.; Albina, J.E. Use of Ly6G-specific monoclonal antibody to deplete neutrophils in mice. J. Leukoc. Biol. 2007, 83, 64–70. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | HU-308 | HU-910 | RO6871304 | RO6871085 | RO6851228 |
|---|---|---|---|---|---|
| Chemical structure |  |  |  |  |  |
| IUPAC name | {(1S,4S,5S)-4-[2,6-Dimethoxy-4-(2-methyloctan-2-yl)phenyl]-6,6-dimethylbicyclo[3.1.1]hept-2-en-2-yl}methanol | {(1S,4R)-2-[2,6-dimethoxy-4-(2-methyloctan-2-yl)phenyl]-7,7-dimethylbicyclo[2.2.1]hept-2-en-1-yl}methanol | (3S)-1-{5-tert-butyl-3-[(1-cyclopropyl-1H-tetrazol-5-yl)methyl]-3H-[1,2,3]triazolo[4,5-d]pyrimidin-7-yl}pyrrolidin-3-ol | 5-Cyclopropyl-N-[(2S)-1-cyclopropyl-2-(5-methyl-1,2,4-oxadiazol-3-yl)propan-2-yl]-4-(2,2,2-trifluoroethoxy)pyridine-2-carboxamide | 5-Cyclopropyl-N-[(2S)-3,3-dimethyl-1-(methylamino)-1-oxobutan-2-yl]-4-[(3-methyloxetan-3-yl)methoxy]pyridine-2-carboxamide |
| Compound | HU308 | HU910 | RO6871304 | RO6871085 | RO6851228 |
|---|---|---|---|---|---|
| hCB2R: Ki (nM) | 10 * | 43 * | 17 | 76 * | 45 * |
| hCB1R: Ki (nM) | 4600 * | 3734 * | >10,000 * | 3378 * | 6487 * |
| hCB1R Ki/hCB2R Ki | 460 | 89 | >588 * | 44 * | 147 * |
| mouse CB2R: Ki (nM) | 223 | 276 | 34 | 6 * | 1 * |
| hCB2R: cAMP EC50 (nM), efficacy (%) | 3, 98 * | 4, 98 * | 1, 102 * | 8, 88 * | 26, −159 * |
| hCB1R: cAMP EC50 (nM) | >10,000 * | >10,000 * | >10,000 * | 1449, 38 * | >10,000 |
| hCB1R cAMP EC50/hCB2R cAMP EC50 | >3333 | >2500 | >10,000 | 174 * | >383 * |
| mouse CB2R: cAMP EC50 (nM), efficacy (%) | 6, 99 | 3, 99 | 0.5, 101 | 3, 81 * | 4, −174 * |
| hCB2R: beta arrestin EC50 (nM), efficacy (%) | 101, 107 * | 109, 81 * | 21, 119 * | 55, 51 | 3, −18 |
| hCB1R: beta arrestin EC50 (nM) | >10,000 * | >30,000 * | >30,000 | >30,000 * | n.d. |
| hCB1R beta arrestin EC50/hCB2R beta arrestin EC50 | >99 | >275 | >1429 | >545 | n.d. |
| mouse CB2R: beta arrestin EC50 (nM), efficacy (%) | 592, 76 | 224, 54 | 26, 94 | >30,000 | >30,000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porter, R.F.; Szczesniak, A.-M.; Toguri, J.T.; Gebremeskel, S.; Johnston, B.; Lehmann, C.; Fingerle, J.; Rothenhäusler, B.; Perret, C.; Rogers-Evans, M.; et al. Selective Cannabinoid 2 Receptor Agonists as Potential Therapeutic Drugs for the Treatment of Endotoxin-Induced Uveitis. Molecules 2019, 24, 3338. https://doi.org/10.3390/molecules24183338
Porter RF, Szczesniak A-M, Toguri JT, Gebremeskel S, Johnston B, Lehmann C, Fingerle J, Rothenhäusler B, Perret C, Rogers-Evans M, et al. Selective Cannabinoid 2 Receptor Agonists as Potential Therapeutic Drugs for the Treatment of Endotoxin-Induced Uveitis. Molecules. 2019; 24(18):3338. https://doi.org/10.3390/molecules24183338
Chicago/Turabian StylePorter, Richard Frederick, Anna-Maria Szczesniak, James Thomas Toguri, Simon Gebremeskel, Brent Johnston, Christian Lehmann, Jürgen Fingerle, Benno Rothenhäusler, Camille Perret, Mark Rogers-Evans, and et al. 2019. "Selective Cannabinoid 2 Receptor Agonists as Potential Therapeutic Drugs for the Treatment of Endotoxin-Induced Uveitis" Molecules 24, no. 18: 3338. https://doi.org/10.3390/molecules24183338
APA StylePorter, R. F., Szczesniak, A.-M., Toguri, J. T., Gebremeskel, S., Johnston, B., Lehmann, C., Fingerle, J., Rothenhäusler, B., Perret, C., Rogers-Evans, M., Kimbara, A., Nettekoven, M., Guba, W., Grether, U., Ullmer, C., & Kelly, M. E. M. (2019). Selective Cannabinoid 2 Receptor Agonists as Potential Therapeutic Drugs for the Treatment of Endotoxin-Induced Uveitis. Molecules, 24(18), 3338. https://doi.org/10.3390/molecules24183338