Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress




Abstract
1. Introduction
2. Results
2.1. Thiobarbituric-Acid-Reactive Substances (TBARS) Levels
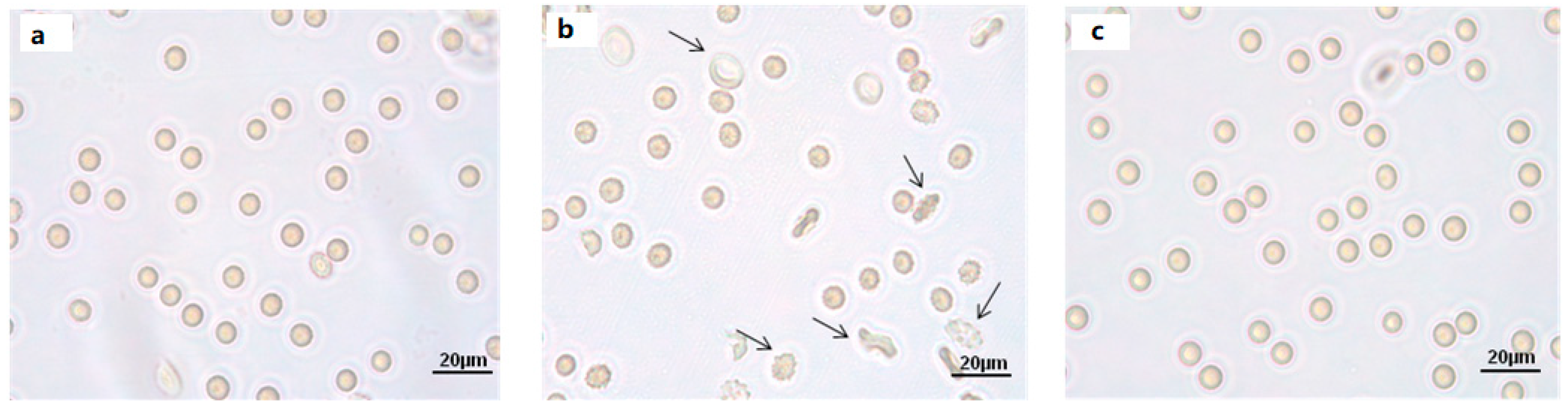
2.2. Cell Shape
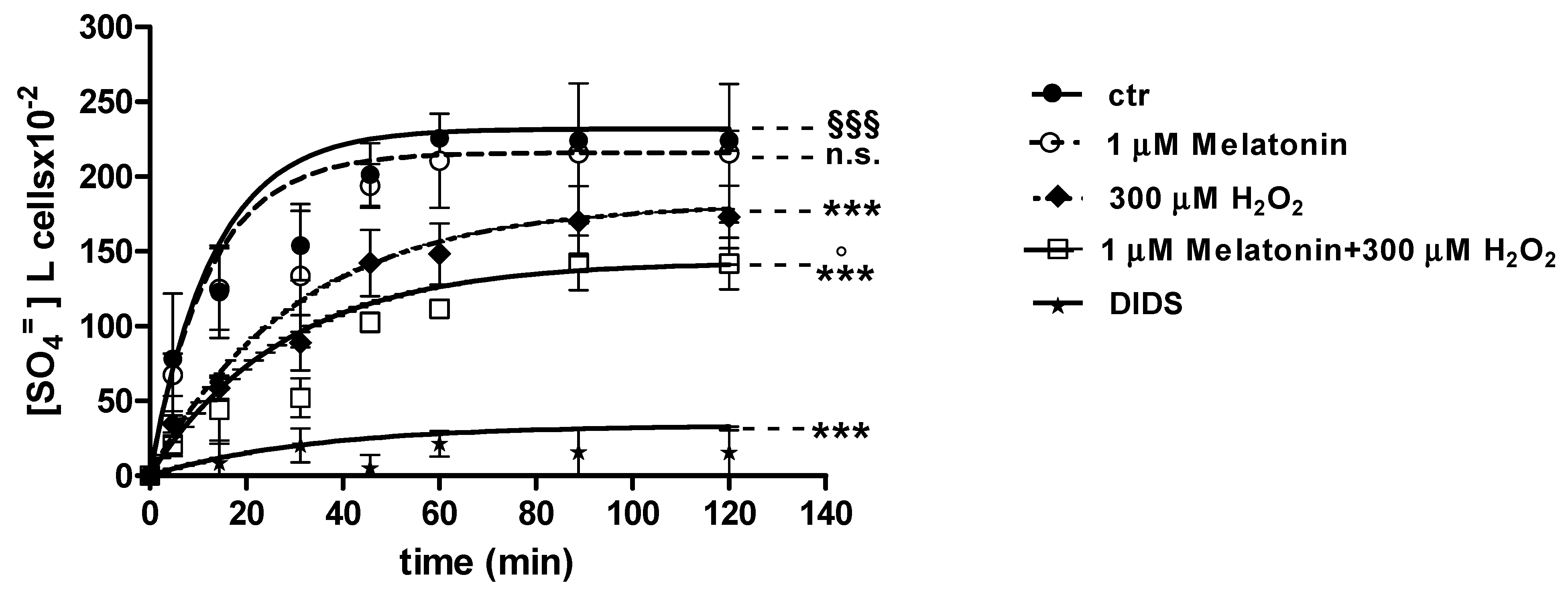
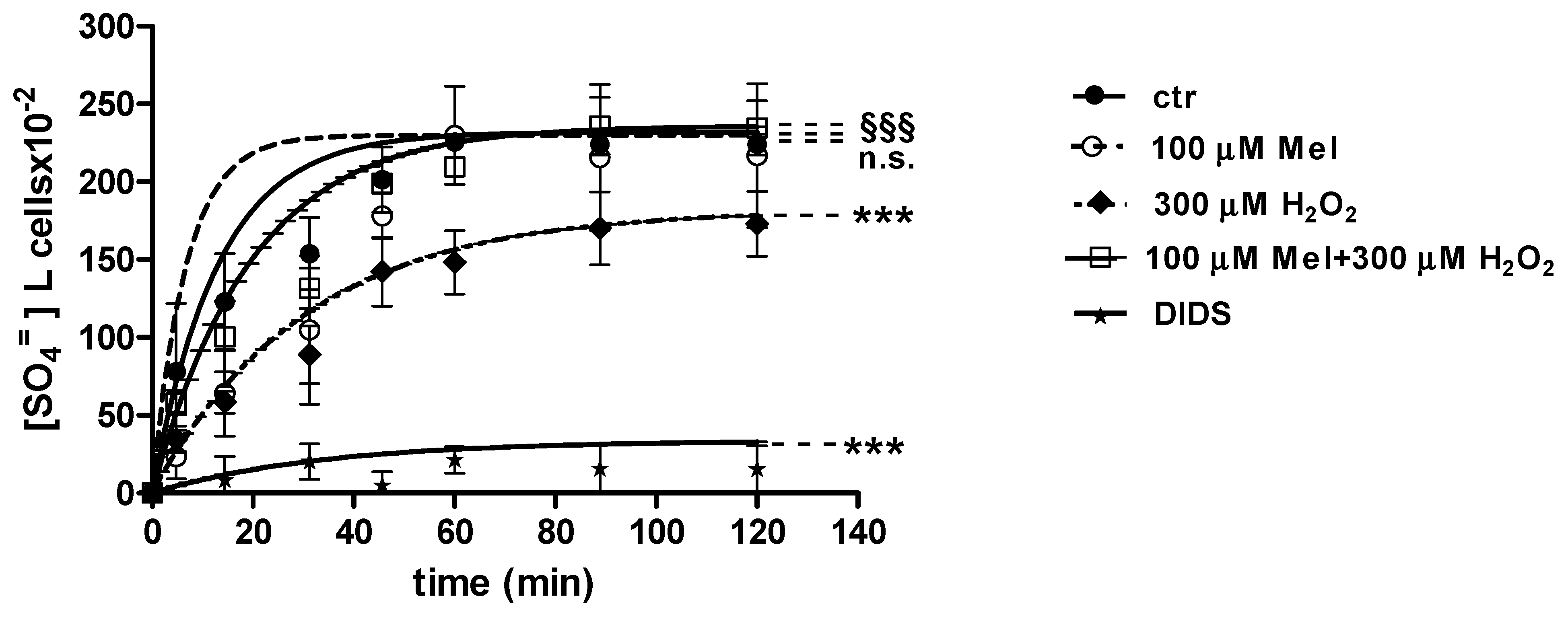
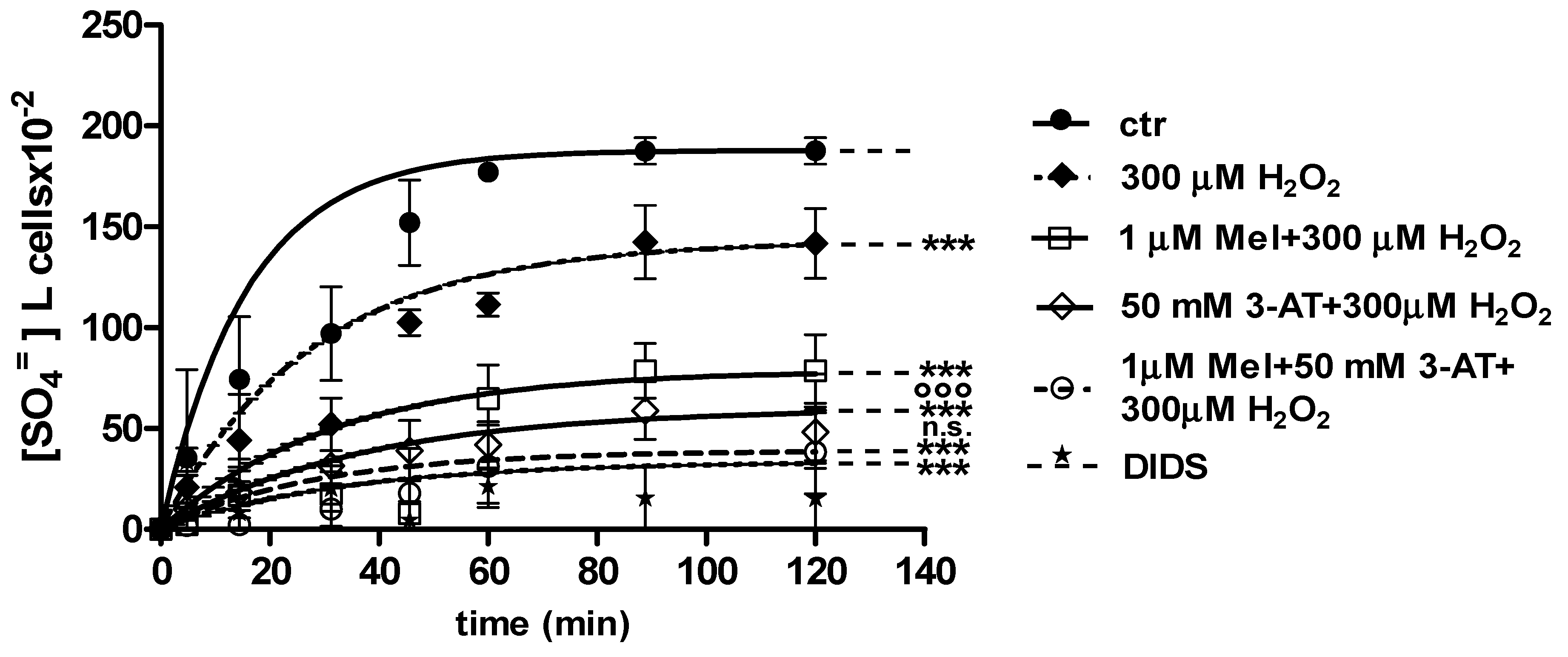
2.3. SO4=Uptake Measurement
2.3.1. H2O2 Treatment
2.3.2. H2O2 and 3-AT Treatment
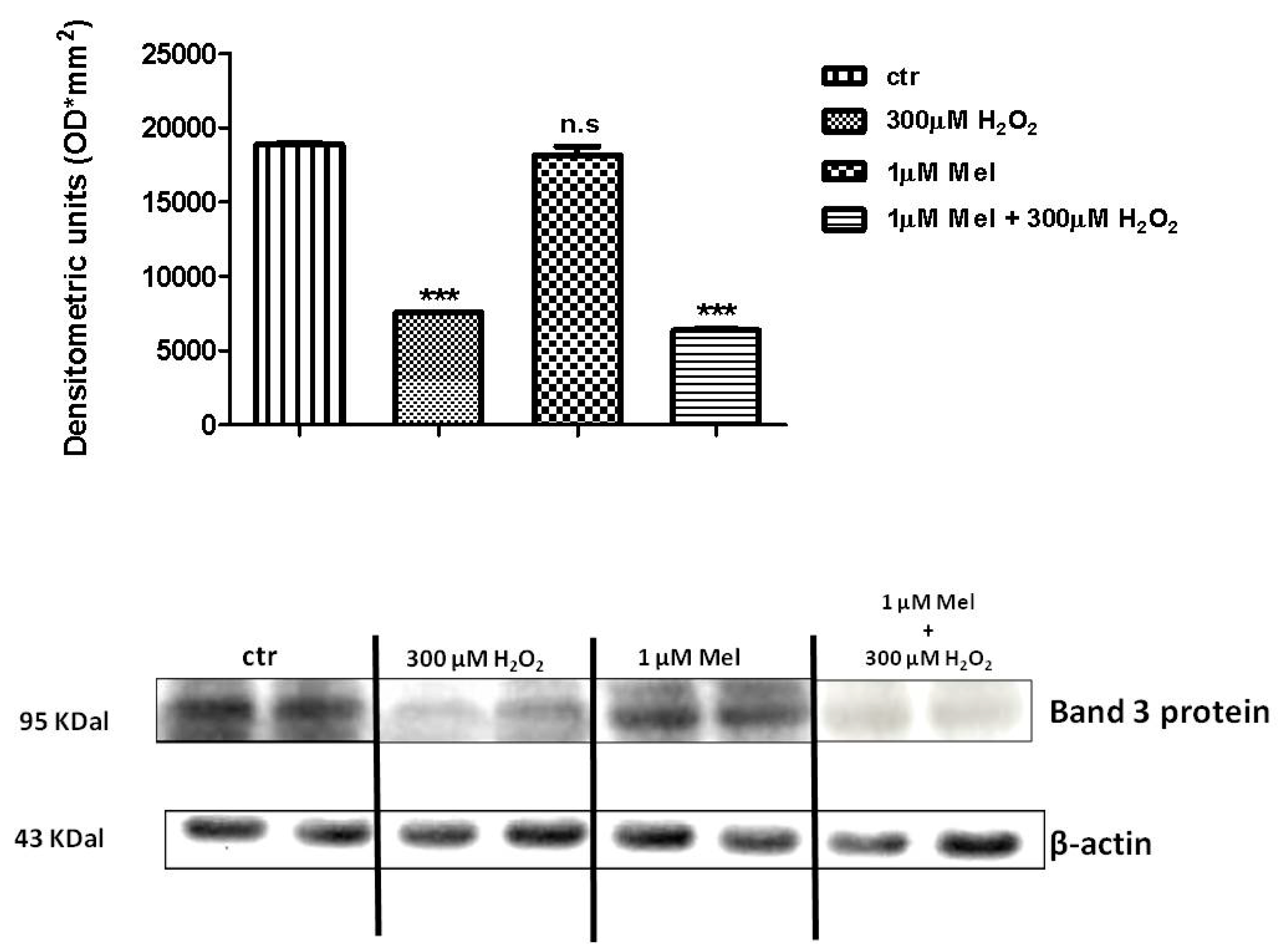
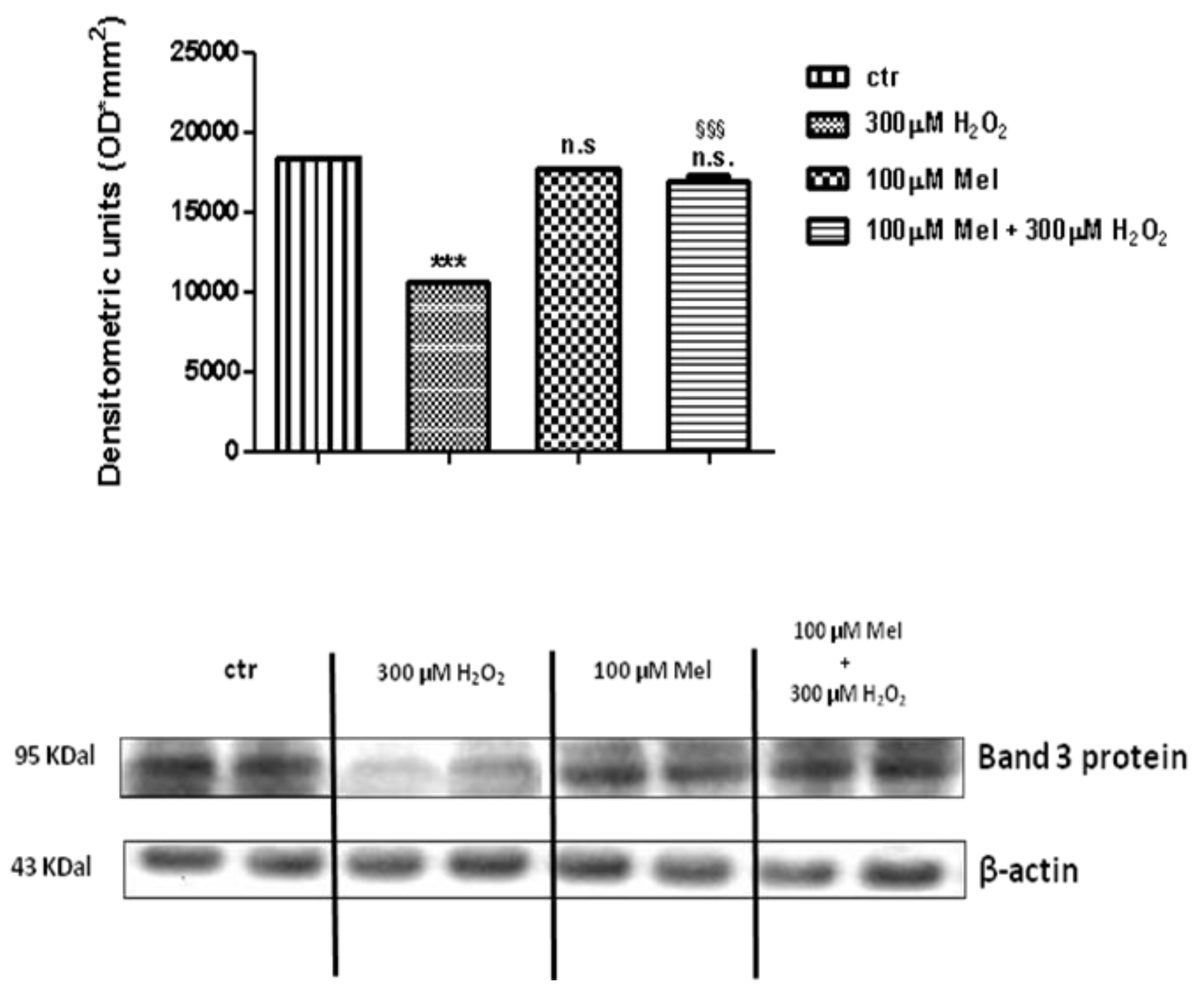
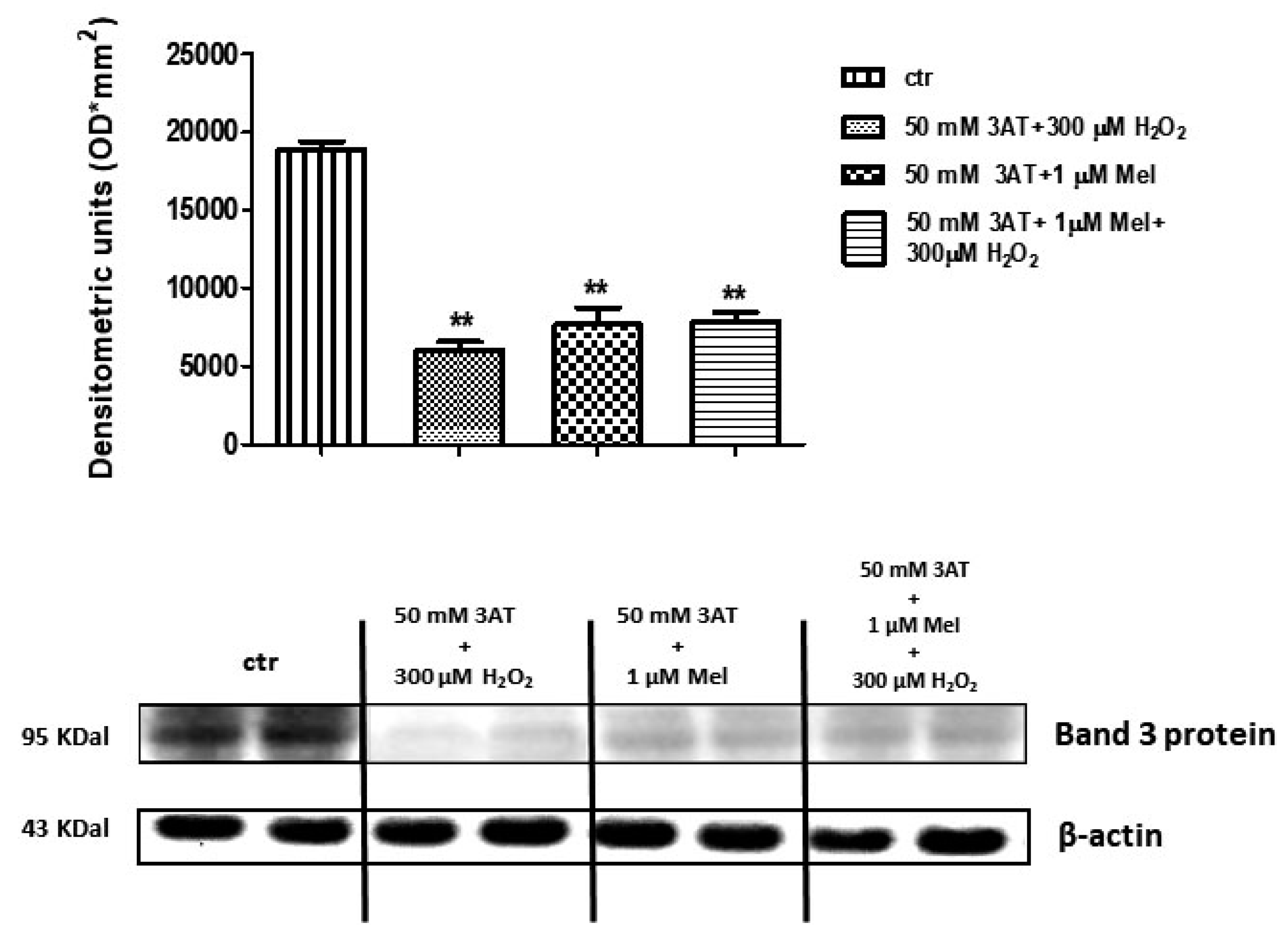
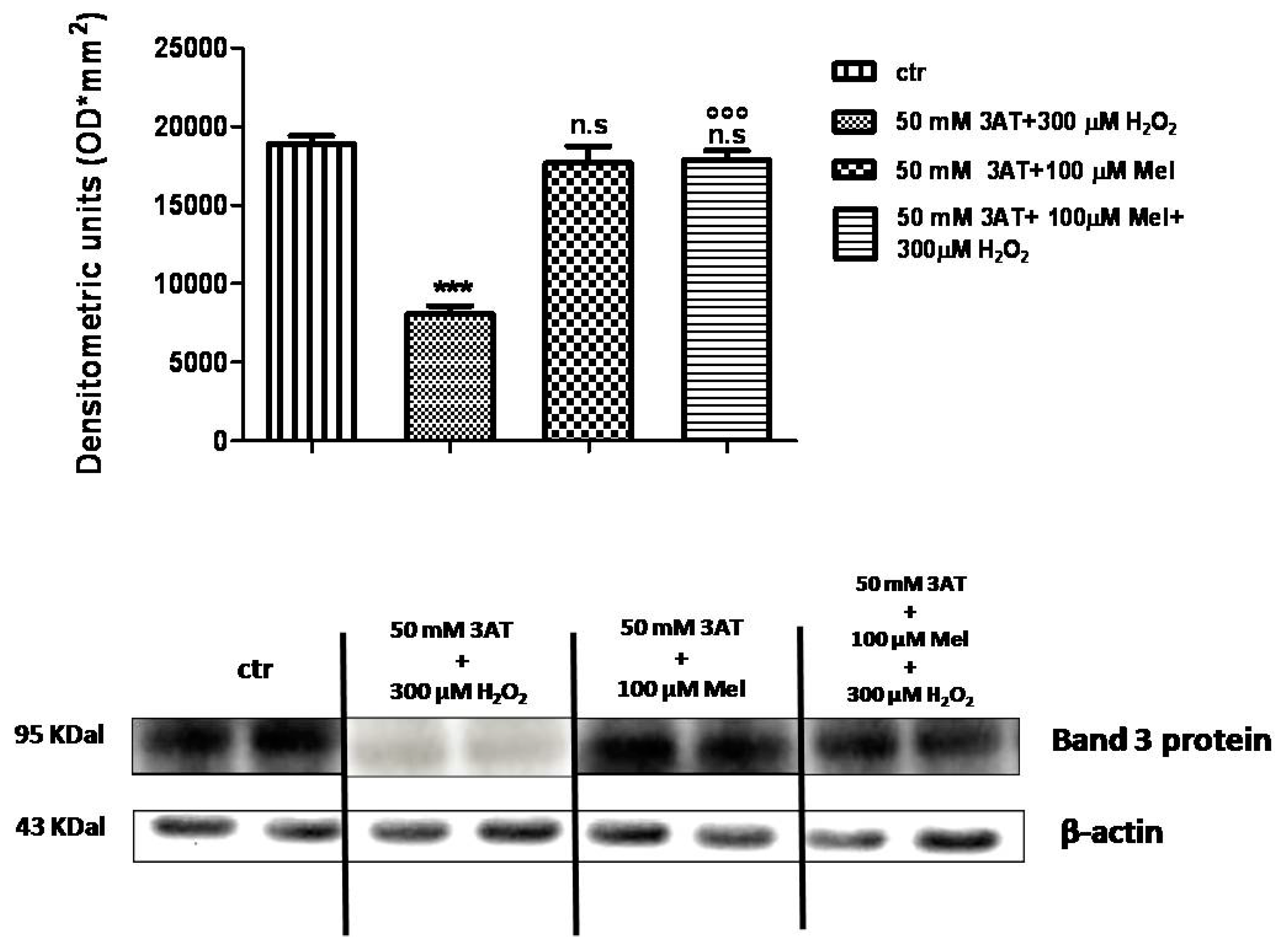
2.4. Band 3 Protein Expression Levels Determination
3. Discussion
4. Materials and Methods
4.1. Solutions and Chemicals
4.2. Erythrocytes Preparation
4.3. Lipid Peroxidation
4.4. SO4= Uptake Measurement
4.4.1. Control Conditions
4.4.2. Exposure to H2O2
4.5. Light microscope Observations
4.6. Erythrocytes Membranes Preparation and SDS-PAGE
4.7. Western Blot Analysis
4.8. Catalase Inhibition
4.9. Experimental Data and Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Reithmeier, R.A.F.; Casey, J.R.; Kall, A.C.; Sansomc, M.S.P.; Alguel, Y.; Iwata, S. Band 3, the human red cell chloride/bicarbonate anion exchanger (AE1, SLC4A1), in a structural context. Biochim. Biophys. Acta 2016, 1858, 1507–1532. [Google Scholar] [CrossRef]
- Jennings, M.L. Proton fluxes associated with erythrocyte membrane anion exchange. J. Membr. Biol. 1976, 28, 187–205. [Google Scholar] [CrossRef]
- Romano, L.; Passow, H. Characterization of anion transport system in trout red blood cell. Am. J. Physiol. 1984, 246, 330–338. [Google Scholar] [CrossRef]
- Morabito, R.; Romano, O.; La Spada, G.; Marino, A. H2O2-induced oxidative stress affects SO4= transport in human erythrocytes. PLoS ONE 2016, 11, e0146485. [Google Scholar] [CrossRef]
- Mueller, S.; Riedel, H.D.; Stremmel, W. Determination of catalase activity at physiological hydrogen peroxide concentrations. Anal. Biochem. 1997, 245, 55–60. [Google Scholar] [CrossRef]
- Anong, W.A.; Franco, T.; Chu, H.; Weis, T.L.; Devlin, E.E.; Bodine, D.M.; An, X.; Mohandas, N.; Low, P.S. Adducin forms a bridge between the erythrocyte membrane and its cytoskeleton and regulates membrane cohesion. Blood 2009, 114, 1904–1912. [Google Scholar] [CrossRef]
- De Luca, G.; Gugliotta, T.; Scuteri, A.; Romano, P.; Rinaldi, C.; Sidoti, A.; Amato, A.; Romano, L. The interaction of haemoglobin, magnesium, organic phosphates and band 3 protein in nucleated and anucleated erythrocytes. Cell. Biochem. Funct. 2004, 22, 179–186. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Bagnato, G.; Roberts, W.N.; Sciortino, D.; D’Angelo, T.; Iannelli, F.; Iannelli, D.; Cordova, F.; Cirillo, M.; et al. Band 3 protein function and oxidative stress in erythrocytes from Systemic Sclerosis patients with interstitial lung disease. Eur. J. Clin. Biomed. Sci. 2017, 3, 80–84. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Cavallaro, M.; Taormina, A.; La Spada, G.; Marino, A. Anion exchange through band 3 protein in canine leishmaniasis at different stages of disease. Pflugers Arch. 2017, 469, 713–724. [Google Scholar] [CrossRef]
- Teti, D.; Crupi, M.; Busá, M.; Valenti, A.; Loddo, S.; Mondello, M.; Romano, L. Chemical and pathological oxidative influences on band 3 protein anion-exchanger. Cell. Physiol. Biochem. 2005, 16, 77–86. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Marino, A. Protective role of magnesium against oxidative stress on SO4= uptake through Band 3 protein in human erythrocytes. Cell. Physiol. Biochem. 2019, 52, 1292–1308. [Google Scholar]
- Konturek, S.J.; Konturek, P.C.; Brozozowska, I.; Pawlik, M.; Sliwowski, Z.; Czenikiewicz-Guzik, M.; Kwiecień, S.; Brzozowski, T.; Bubenik, G.A.; Pawlik, W.W. Localization and biological activities of melatonin in intact and diseased gastrointestinal tract (GIT). J. Physiol. Pharmacol. 2007, 58, 381–405. [Google Scholar]
- Hevia, D.; Gonzàlez-Menèndez, P.; Quiros-Gonzàlez, I.; Miar, A.; Rodríguez-García, A.; Tan, D.X.; Reiter, R.J.; Mayo, J.C.; Sainz, R.M. Melatonin uptake through glucose transporters: A new target for melatonin inhibition of cancer. J. Pineal Res. 2015, 58, 234–250. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. Camb. Philos. Soc. 2010, 85, 607–623. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhang, Y. Melatonin A well-documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef]
- Abuja, P.M.; Liebmann, P.; Hayn, M.; Schauenstein, K.; Esterbauer, H. Antioxidant role of melatonin in lipid peroxidation of human LDL. FEBS Lett. 1997, 413, 289–293. [Google Scholar] [CrossRef]
- Chakravarty, S.; Rizvi, S.I. Day and Night GSH and MDA levels in healthy adults and effects of different doses of melatonin on these parameters. Int. J. Cell Biol. 2011, 2011, 404591. [Google Scholar] [CrossRef]
- Duell, P.B.; Wheaton, D.L.; Shultz, A.; Nguyen, H. Inhibition of LDL oxidation by melatonin requires supraphysiologic concentrations. Clin. Chem. 1998, 44, 1931–1936. [Google Scholar]
- Da Silva, D.G.H.; Chaves, N.A.; Miyamoto, S.; de Almeida, E.A. Prolonged erythrocyte auto-incubation as an alternative model for oxidant generation system. Toxicol. In Vitro 2019, 56, 62–74. [Google Scholar] [CrossRef]
- Buehler, P.W.; Alayash, A.I. Redox biology of blood revisited: The role of red blood cells in maintaining circulatory reductive capacity. Antioxid. Redox Signal. 2005, 7, 1755–1760. [Google Scholar] [CrossRef]
- Dikmenoglu, N.; Ileri, E.; Seringec, N.; Ercil, D. Melatonin prevents lipid peroxidation in human erythrocytes but augments deterioration of deformability after in vitro oxidative stress. Clin. Hemorheol. Microcirc. 2008, 40, 235–242. [Google Scholar]
- Morabito, R.; Remigante, A.; Di Pietro, M.L.; Giannetto, A.; La Spada, G.; Marino, A. SO4= uptake and catalase role in preconditioning after H2O2-induced oxidative stress in human erythrocytes. Pflugers Arch. 2017, 469, 235–250. [Google Scholar] [CrossRef]
- Stefanovic, M.; Markham, N.O.; Parry, E.M.; Garrett-Beal, L.J.; Cline, A.P.; Gallagher, P.G.; Low, P.S.; Bodine, D.M. An 11-amino acid beta-hairpinloop in the cytoplasmic domain of band 3 is responsible for ankyrin binding in mouse erythrocytes. Proc. Natl. Acad. Sci. USA 2007, 104, 13972–13977. [Google Scholar] [CrossRef]
- Dumaswala, U.J.; Zhuo, L.; Jacobsen, D.W.; Jain, S.K.; Sukalski, K.A. Protein and lipid oxidation of banked human erythrocytes: Role of glutathione. Free Radic. Biol. Med. 1999, 27, 1041–1049. [Google Scholar] [CrossRef]
- Winterbourn, C.C. The biological chemistry of hydrogen peroxide. Methods Enzymol. 2013, 528, 3–25. [Google Scholar]
- Shiga, T.; Maeda, N.; Kon, K. Erythrocyte rheology. Crit. Rev. Oncol. Hematol. 1990, 10, 9–48. [Google Scholar] [CrossRef]
- Morabito, R.; Falliti, G.; Geraci, A.; La Spada, G.; Marino, A. Curcumin protects–SH groups and sulphate transport after oxidative damage in human erythrocytes. Cell. Physiol. Biochem. 2015, 36, 345–357. [Google Scholar] [CrossRef]
- Romano, L.; Scuteri, A.; Gugliotta, T.; Romano, P.; De Luca, G.; Sidoti, A.; Amato, A. Sulphate influx in the erythrocytes of normotensive, diabetic and hypertensive patients. Cell. Biol. Int. 2002, 26, 421–426. [Google Scholar] [CrossRef]
- Galano, A.; Reiter, R.J. Melatonin and its metabolites vs. oxidative stress: From individual actions to collective protection. J. Pineal Res. 2018, 65, e12514. [Google Scholar] [CrossRef]
- Venegas, C.; Garcıa, J.A.; Escames, G.; Ortiz, F.; Lopez, A.; Doerrier, C.; Garcıa-Corzo, L.; Lopez, L.C.; Reiter, R.J.; Acuna-Castroviejo, D. Extrapineal melatonin: Analysis of its subcellular distribution and daily fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef]
- Da Silva, D.G.H.; Ricci, O., Jr.; Alves de Almeida, E.; Bonini-Domingos, C.R. Potential utility of melatonin as an antioxidant therapy in the management of sickle cell anemia. J. Pineal Res. 2015, 58, 178–188. [Google Scholar] [CrossRef]
- Ciftci, M.; Dilek, B.; Kufrevioglu, O.I. Effects of melatonin on enzyme activities of glucose 6-phosphate dehydrogenase from human erythrocytes in vitro and from rat erythrocytes in vivo. Pharmacol. Res. 2001, 44, 7–11. [Google Scholar] [CrossRef]
- Tailleux, A.; Torpier, G.; Bonnefont-Rousselot, D.; Lestavel, S.; Lemdani, M.; Caudeville, B.; Furman, C.; Foricher, R.; Gardes-Albert, M.; Lesieur, D. Daily melatonin supplementation in mice increases atherosclerosis in proximal aorta. Biochem. Biophys. Res. Commun. 2002, 293, 1114–1123. [Google Scholar] [CrossRef]
- Fulia, F.; Gitto, E.; Cuzzocrea, S.; Reiter, R.J.; Dugo, L.; Gitto, P.; Barberi, S.; Cordaro, S.; Barberi, I. Increased levels of malondialdehyde and nitrite/nitrate in the blood of asphyxiated newborns: Reduction by melatonin. J. Pineal Res. 2001, 31, 343–349. [Google Scholar] [CrossRef]
- García, J.J.; Piñol-Ripoll, G.; Martínez-Ballarín, E.; Fuentes-Broto, L.; Miana-Mena, F.J.; Venegas, C.; Caballero, B.; Escames, G.; Coto-Montes, A.; Acuña-Castroviejo, D. Melatonin reduces membrane rigidity and oxidative damage in the brain of SAMP8 mice. Neurobiol. Aging 2011, 32, 2045–2054. [Google Scholar] [CrossRef]
- Cimen, M.Y. Free radical metabolism in human erythrocytes. Clin. Chim. Acta 2008, 390, 1–11. [Google Scholar] [CrossRef]
- García, J.J.; López-Pingarrón, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.; Kim, S.J.; Manchester, L.C.; Qi, W.; Garcia, J.J.; Cabrera, J.C.; El-Sokkary, G.; Rouvier-Garay, V. Augmentation of indices of oxidative damage in life-long melatonin-deficient rats. Mech. Ageing Dev. 1999, 110, 157–173. [Google Scholar] [CrossRef]
- Tesoriere, L.; D’Arpa, D.; Conti, S.; Giaccone, V.; Pintaudi, A.M.; Livrea, M.A. Melatonin protects human red blood cells from oxidative hemolysis: New insights into the radical-scavenging activity. J. Pineal Res. 1999, 27, 95–105. [Google Scholar] [CrossRef]
- Srour, M.A.; Bilto, Y.Y.; Jouma, M.; Irhimeh, M.R. Exposure of human erythrocytes to oxygen radicals causes loss of deformability, increased osmotic fragility, lipid peroxidation and protein degradation. Clin. Hemorheol. Microcirc. 2000, 23, 13–21. [Google Scholar]
- Welbourn, E.M.; Wilson, M.T.; Yusof, A.; Metodiev, M.V.; Cooper, C.E. The mechanism of formation, structure and physiological relevance of covalent hemoglobin attachment to the erythrocyte membrane. Free Radic. Biol. Med. 2017, 95–106. [Google Scholar] [CrossRef]
- Miller, E.; Walczak, A.; Majsterek, I.; Kedziora, J. Melatonin reduces oxidative stress in the erythrocytes of multiple patients with secondary progressive clinical course. J. Neuroimmunol. 2013, 257, 97–101. [Google Scholar] [CrossRef]
- Erat, M.; Ciftci, M. Effect of melatonin on enzyme activities of glutathione reductase from human erythrocytes in vitro and from rat erythrocytes in vivo. Eur. J. Pharmacol. 2006, 537, 59–63. [Google Scholar] [CrossRef]
- Emamgholipour, S.; Hossein-Nezhad, A.; Ansari, M. Can Melatonin Act as an Antioxidant in Hydrogen Peroxide-Induced Oxidative Stress Model in Human Peripheral Blood Mononuclear Cells? Biochem. Res. Int. 2016, 2016, 5857940. [Google Scholar] [CrossRef]
- Margoliash, E.; Novogrodsky, A.; Schejter, A. Irreversible reaction of 3-amino-1,2,4-triazole and related inhibitors with the protein of catalase. Biochem. J. 1960, 74, 339–348. [Google Scholar] [CrossRef]
- Almroth, B.C.; Sturve, J.; Berglund, A.; Forlin, L. Oxidative damage in eelpout (Zoarces viviparus), measured as protein carbonyls and TBARS, as biomarkers. Aquatic Toxicol. 2005, 73, 171–180. [Google Scholar] [CrossRef]
- Jessen, F.; Sjoholm, C.; Hoffmann, E.K. Identification of the anion exchange protein of Ehrlich cells: A kinetic analysis of the inhibitory effects of 4,4′-diisothiocyano-2,2′-stilbene-disulfonic acid(DIDS) and labeling of membrane proteins with 3H-DIDS. J. Membr. Biol. 1986, 92, 195–205. [Google Scholar] [CrossRef]
- Pantaleo, A.; Ferru, E.; Pau, M.C.; Khadjavi, A.; Mandili, G.; Mattè, A.; Spano, A.; De Franceschi, L.; Pippia, P.; Turrini, F. Band 3 erythrocyte membrane protein acts as redox stress sensor leading to its phosphorylation by p (72) Syk. Oxid. Med. Cell. Longev. 2016, 1–11. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
Sample Availability: Not available. |












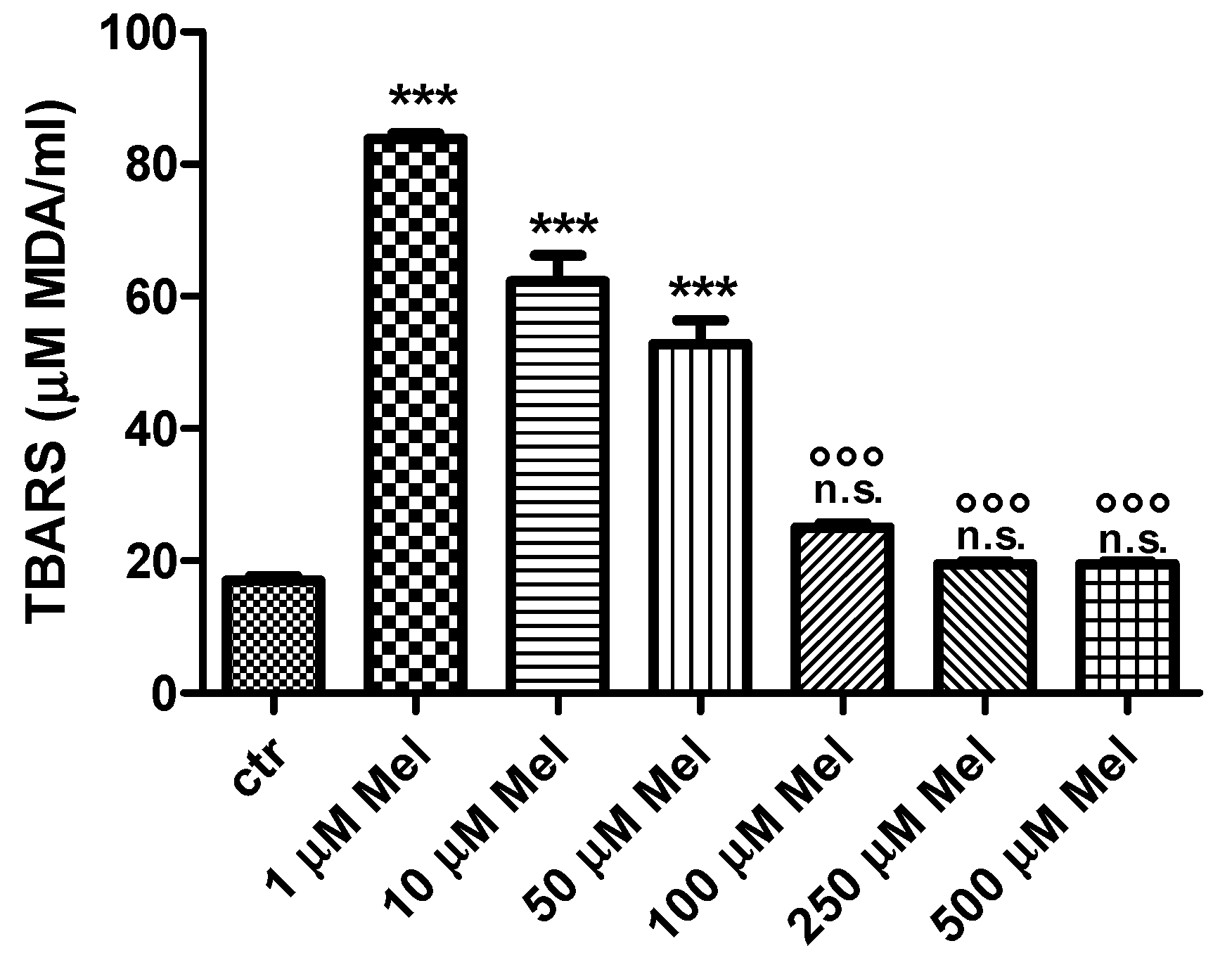{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rate Constant (min−1) | Time (min) | % Decrease | N | SO4=Amount Internalized at 45 min of SO4=Medium Incubation [SO4=] L cells × 10−2 | |
|---|---|---|---|---|---|
| Control | 0.076 ± 0.001 | 12 | 0 | 10 | 201.3 ± 21 |
| 1 µM Mel | 0.080± 0.001 n.s. | 12.5 | 0 | 8 | 192.8 ± 23 n.s |
| 100 µM Mel | 0.072 ± 0.001 n.s. | 13 | 0 | 8 | 229.8 ± 14.5 n.s |
| 300 µM H2O2 | 0.046 ± 0.001 *** | 21 | 40 | 8 | 142.2 ± 22.2 *** |
| 1 µM Mel + 300 µM H2O2 | 0.048± 0.001 *** | 20 | 37 | 8 | 142.8 ± 21.2 *** |
| 100 µM Mel + 300 µM H2O2 | 0.074 ± 0.001§§§ | 13.5 | 0 | 10 | 176.2 ± 6.3 n.s. |
| 50 mM 3-AT + 300 µM H2O2 | 0.018 ± 0.001 *** | 55.5 | 77 | 9 | 58 ± 12 ***,### |
| 50 mM 3-AT +1 µM Mel + 300 µM H2O2 | 0.019± 0.001 *** | 52.6 | 75 | 8 | 38.4 ± 11 *** |
| 50 mM 3-AT +100 µM Mel + 300 µM H2O2 | 0.077 ± 0.001 n.s. | 12 | 0 | 8 | 219 ± 17 *** |
| 10 µM DIDS | 0.017 ± 0.001 *** | 55 | 78 | 5 | 4.75 ± 9 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morabito, R.; Remigante, A.; Marino, A. Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress. Molecules 2019, 24, 2741. https://doi.org/10.3390/molecules24152741
Morabito R, Remigante A, Marino A. Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress. Molecules. 2019; 24(15):2741. https://doi.org/10.3390/molecules24152741
Chicago/Turabian StyleMorabito, Rossana, Alessia Remigante, and Angela Marino. 2019. "Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress" Molecules 24, no. 15: 2741. https://doi.org/10.3390/molecules24152741
APA StyleMorabito, R., Remigante, A., & Marino, A. (2019). Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress. Molecules, 24(15), 2741. https://doi.org/10.3390/molecules24152741