Genome-Wide Identification of the LAC Gene Family and Its Expression Analysis Under Stress in Brassica napus


 ,
, 
Abstract
1. Introduction
2. Results
2.1. Characterization of the 45 BnLACs
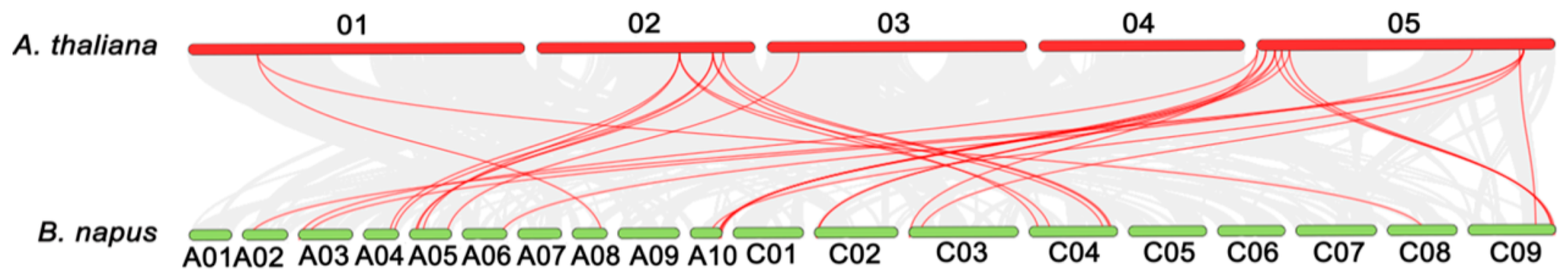
2.2. Phylogenetic Analysis of LACs in A. thaliana, B. napus, B. rapa and B. oleracea
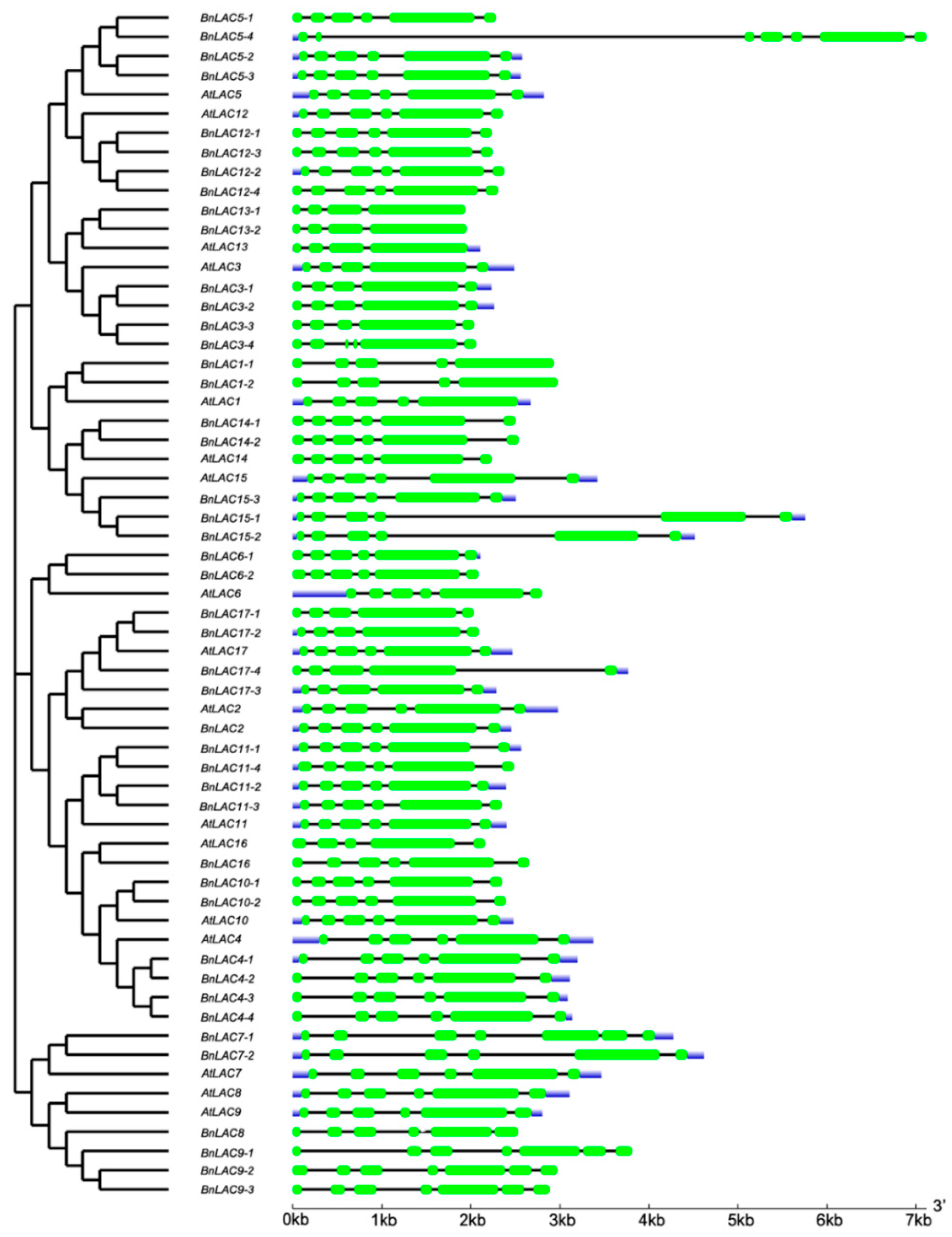
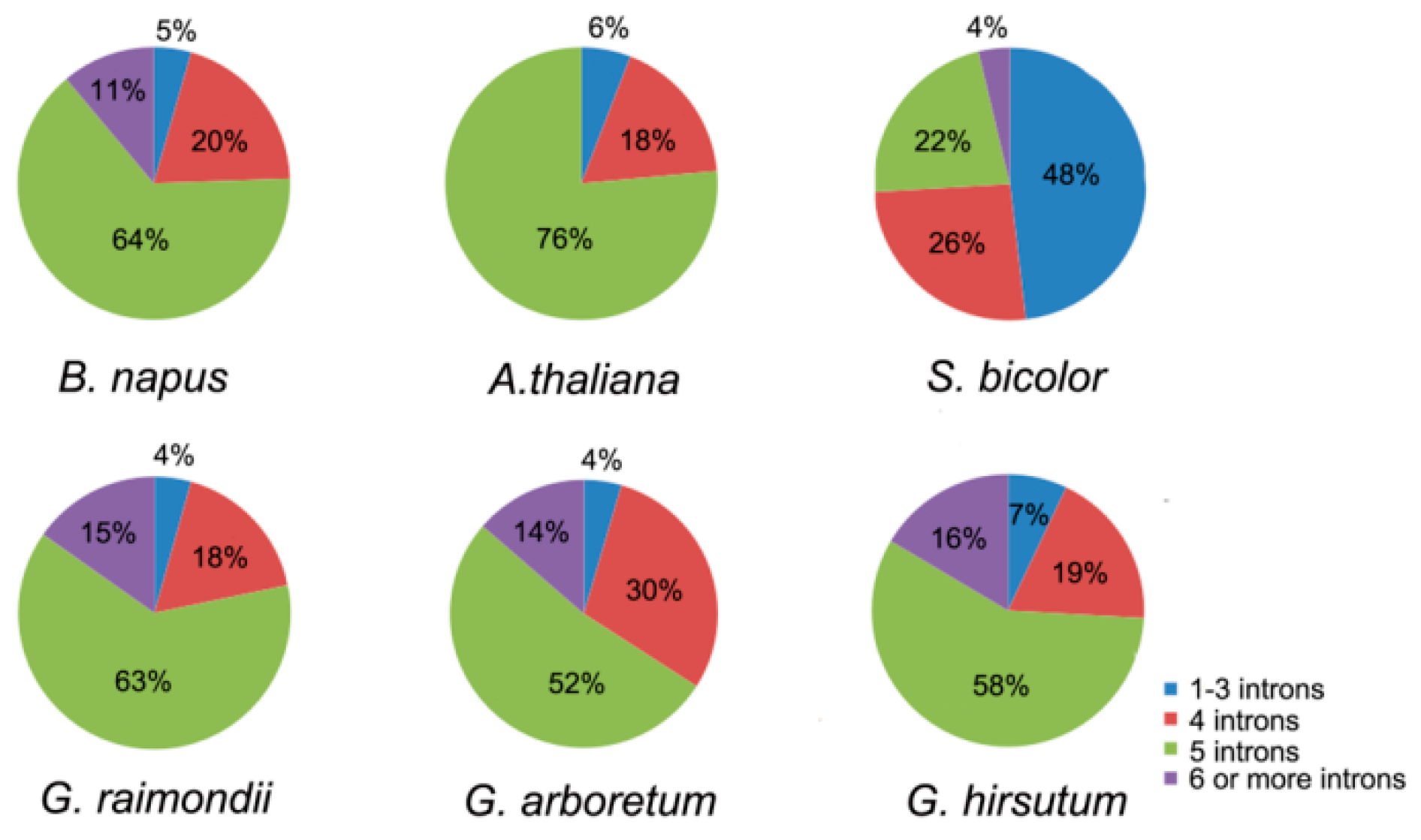
2.3. Gene Structure and Conservative Domain Analysis of BnLACs and AtLACs
2.4. Diverse cis Regulatory Elements and miRNAs are Predicted
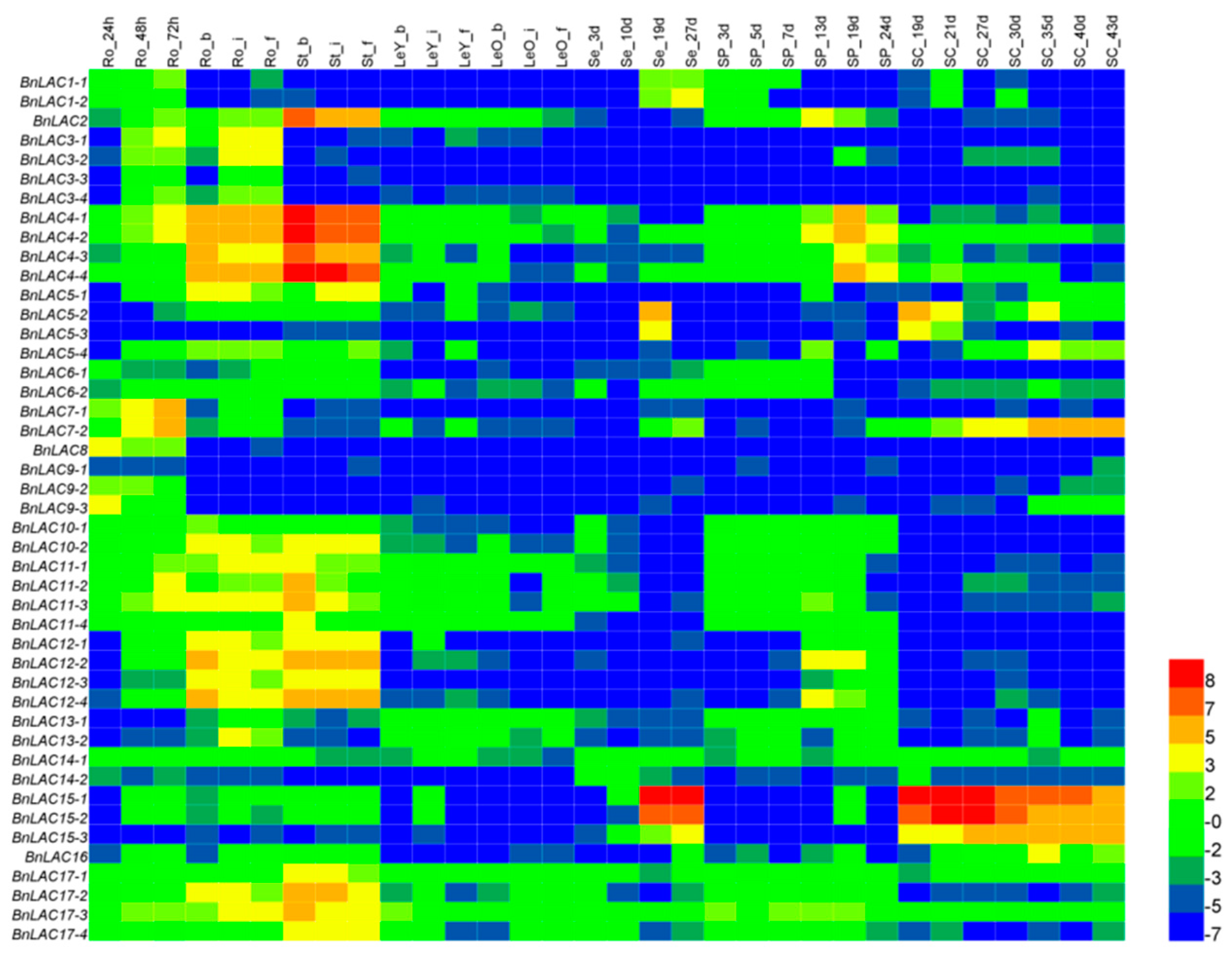
2.5. Expression Pattern Analysis of BnLACs
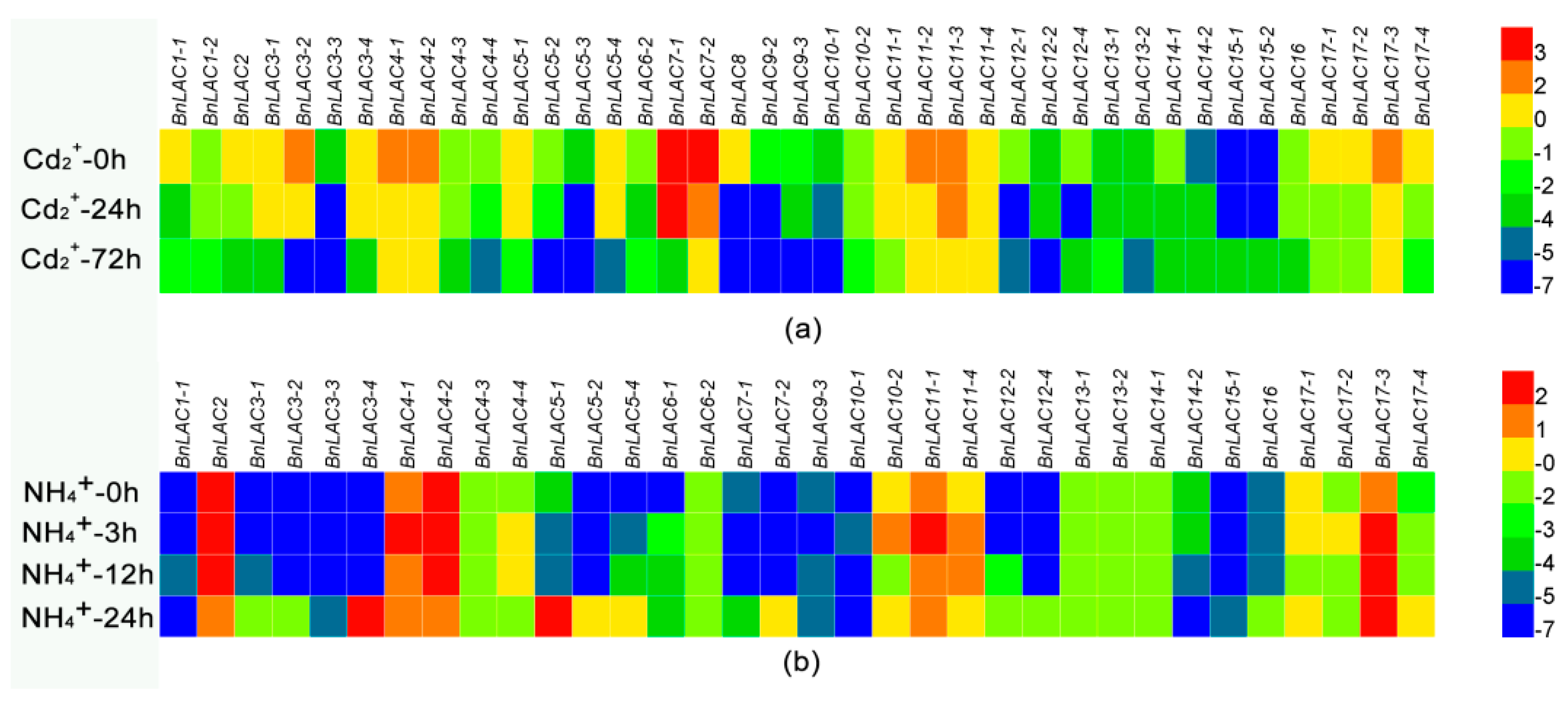
2.6. Responses of BnLACs upon Abiotic Stress
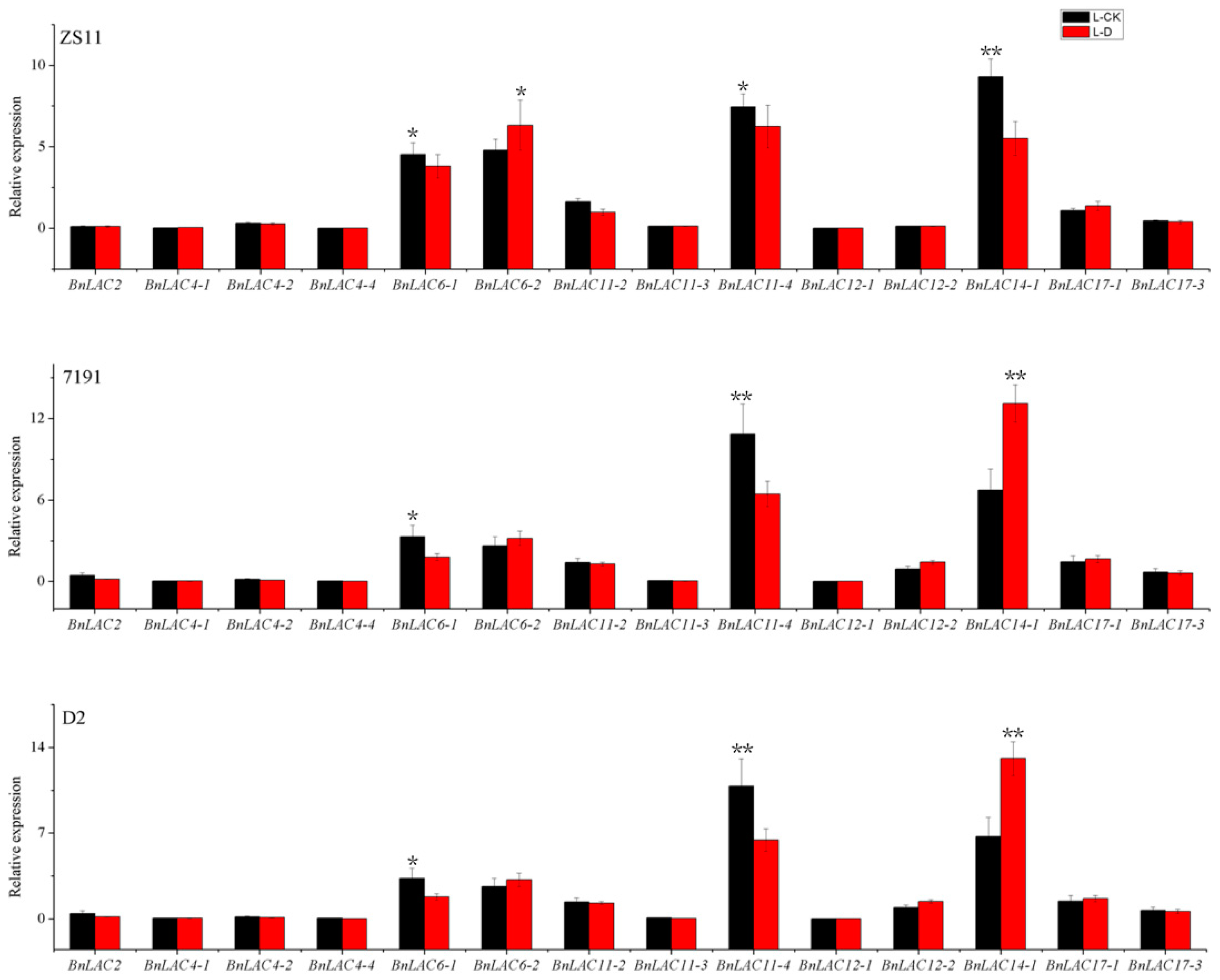
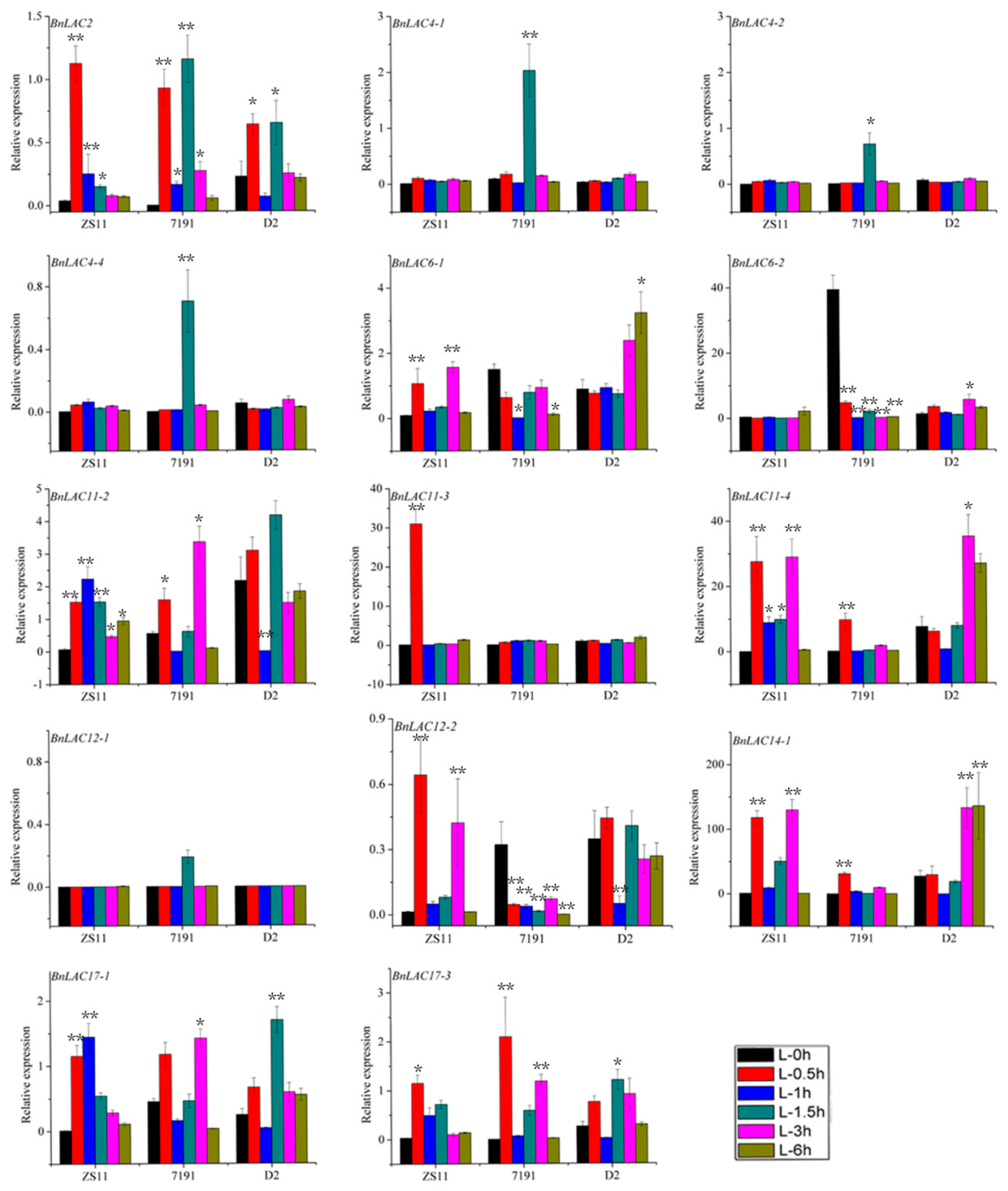
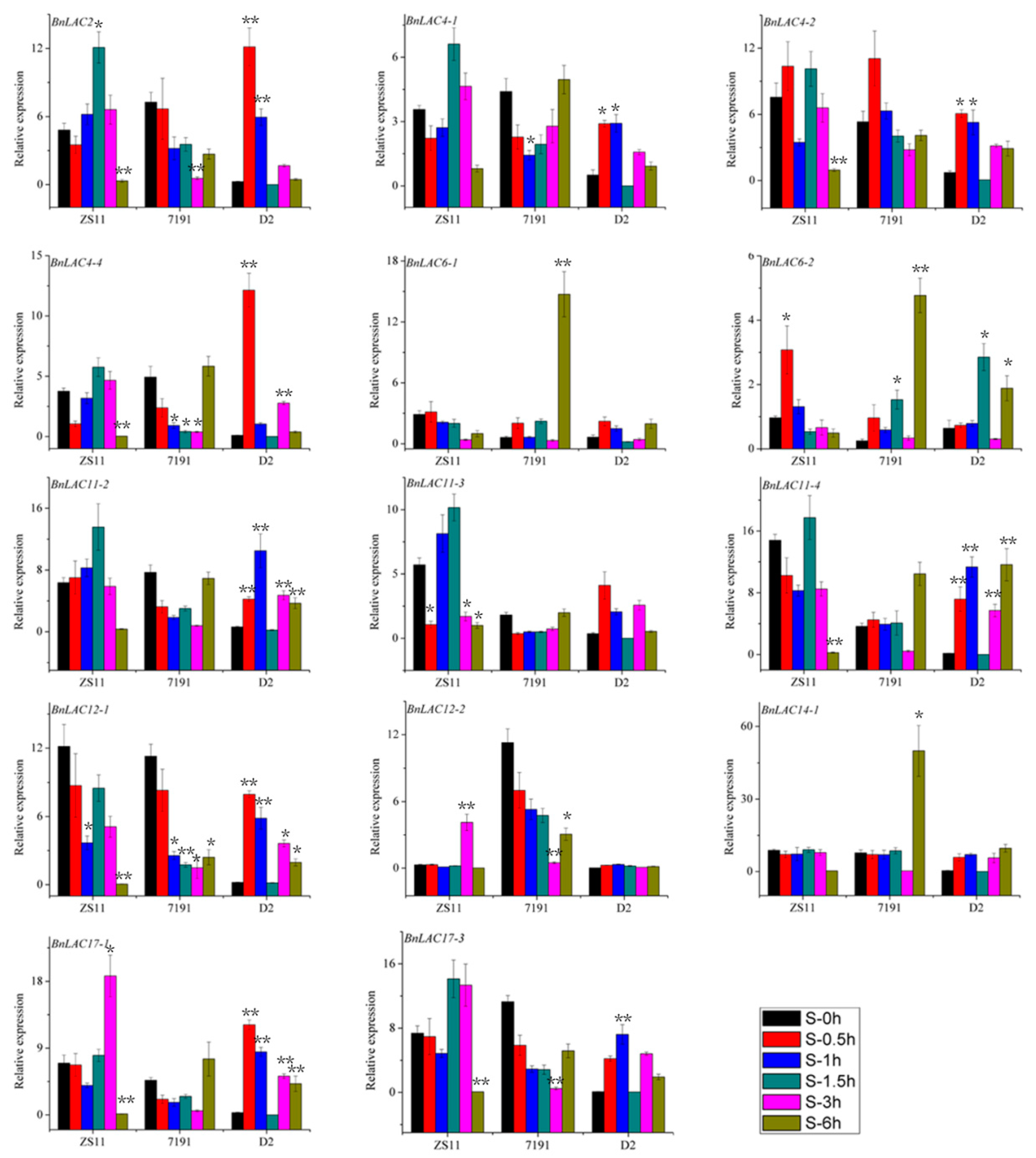
2.7. BnLAC4 and its Predicted Downstream Genes are Differentially Expressed in the Silique Pericarp between Long and Short Silique Lines
3. Discussion
3.1. Loss Events Occurred in the LAC gene family Along with the Evolution
3.2. Regulation of BnLAC Genes
3.3. Expression Patterns and Response to Stress
3.4. BnLAC4 and its Downstream Genes May Participate in Silique Elongation in B. napus
4. Materials and Methods
4.1. Plant Materials and Stress Treatment
4.2. Characterisation of the LAC Gene Family
4.3. Evolutionary relationship of the LAC Genes Family in A. thaliana, B. napus, B. rapa, and B. oleracea
4.4. Gene Structure and Conserved Motif Analysis
4.5. Cis-Elements Analysis and Prediction of miRNA Target BnLACs
4.6. Expression Patterns Analysis of B. napus LAC Genes
4.7. RNA Extraction, Reverse Transcription and qRT-PCR
4.8. Proteins Interaction with AtLAC4 and Identified their Homologous Genes in the B. napus genome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B. Early Allopolyploid Evolution in the Post-Neolithic Brassica Napus Oilseed Genome. Science 2014, 6199, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Mourasobczak, J.; Souza, U.; Mazzafera, P. Drought Stress and Changes in the Lignin Content and Composition in Eucalyptus. BMC Proc. 2011, 7, 103. [Google Scholar]
- Peng, D.; Chen, X.; Yin, Y.; Lu, K.; Yang, W.; Tang, Y.; Wang, Z. Lodging Resistance of Winter Wheat (Triticum aestivum L.): Lignin Accumulation and Its Related Enzymes Activities Due to the Application of Paclobutrazol or Gibberellin Acid. Field Crops Res. 2014, 2, 1–7. [Google Scholar] [CrossRef]
- Cruickshank, A.W.; Cooper, M.; Ryley, M.J.; Cruickshank, A.W.; Cooper, M.; Ryley, M.J. Peanut Resistance to Sclerotinia Minor and S. Sclerotiorum. Aust. J. Agric. Res. 2002, 10, 1105–1110. [Google Scholar] [CrossRef]
- Mayer, A.M.; Harel, E. Polyphenol Oxidase in Plants. Phytochemistry 1979, 18, 193–195. [Google Scholar] [CrossRef]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper Oxidases and Oxygenases. Chem. Rev. 1996, 7, 2563. [Google Scholar] [CrossRef]
- Cohen, R.; Persky, L.; Hadar, Y. Biotechnological Applications and Potential of Wood-Degrading Mushrooms of the Genus Pleurotus. Appl. Microbiol. Biotechnol. 2002, 5, 582–594. [Google Scholar] [CrossRef]
- Bao, W.; O’malley, D.M.; Whetten, R.; Sederoff, R.R. A Laccase Associated with Lignification in Loblolly Pine Xylem. Science 1993, 5108, 672–674. [Google Scholar] [CrossRef] [PubMed]
- Sterjiades, R.; Dean, J.F.; Eriksson, K.E. Laccase from Sycamore Maple (Acer pseudoplatanus) Polymerizes Monolignols. Plant Physiol. 1992, 3, 1162–1168. [Google Scholar]
- Liang, M.; Haroldsen, V.; Cai, X.; Wu, Y. Expression of a Putative Laccase Gene, Zmlac1, in Maize Primary Roots under Stress. Plant Cell Environ. 2006, 5, 746. [Google Scholar] [CrossRef]
- Hüttermann, A.; Mai, C.; Kharazipour, A. Modification of Lignin for the Production of New Compounded Materials. Appl. Microbiol. Biotechnol. 2001, 4, 387–394. [Google Scholar] [CrossRef]
- Wang, G.D.; Li, Q.J.; Luo, B.; Chen, X.Y. Ex Planta Phytoremediation of Trichlorophenol and Phenolic Allelochemicals Via an Engineered Secretory Laccase. Nat. Biotechnol. 2004, 7, 893. [Google Scholar] [CrossRef]
- Balasubramanian, V.K.; Rai, K.M.; Thu, S.W.; Mei, M.H.; Mendu, V. Genome-Wide Identification of Multifunctional Laccase Gene Family in Cotton (Gossypium spp.); Expression and Biochemical Analysis During Fiber Development. Sci. Rep. 2016, 6, 34309. [Google Scholar] [CrossRef]
- Wang, J.; Feng, J.; Jia, W.; Fan, P.; Bao, H.; Li, S.; Li, Y. Genome-Wide Identification of Sorghum Bicolor Laccases Reveals Potential Targets for Lignin Modification. Front. Plant Sci. 2017, 8, 714. [Google Scholar] [CrossRef]
- Lafayette, P.R.; Eriksson, K.E.; Dean, J.F. Nucleotide Sequence of a Cdna Clone Encoding an Acidic Laccase from Sycamore Maple (Acer pseudoplatanus L.). Plant Physiol. 1995, 2, 667–668. [Google Scholar] [CrossRef]
- Kiefermeyer, M.C.; Gomord, V.; O’Connell, A.; Halpin, C.; Faye, L. Cloning and Sequence Analysis of Laccase-Encoding Cdna Clones from Tobacco. Gene 1996, 1–2, 205–207. [Google Scholar] [CrossRef]
- Gavnholt, B.; Larsen, K.; Rasmussen, S.K. Isolation and Characterisation of Laccase Cdnas from Meristematic and Stem Tissues of Ryegrass (Lolium Perenne). Plant Sci. 2002, 6, 873–885. [Google Scholar] [CrossRef]
- Bligny, R.; Gaillard, J.; Douce, R. Excretion of Laccase by Sycamore (Acer pseudoplatanus L.) Cells. Effects of a Copper Deficiency. Biochemical Journal. 1983, 2, 583–588. [Google Scholar]
- Tezuka, K.; Hayashi, M.; Ishihara, H.; Onozaki, K.; Nishimura, M.; Takahashi, N. Occurrence of Heterogeneity of N-Linked Oligosaccharides Attached to Sycamore (Acer pseudoplatanus L.) Laccase after Excretion. Biochem. Mol. Biol. Int. 1993, 3, 395–402. [Google Scholar]
- Ranocha, P.; Mcdougall, G.; Hawkins, S.; Sterjiades, R.; Borderies, G.; Stewart, D.; Cabanesmacheteau, M.; Boudet, A.M.; Goffner, D. Biochemical Characterization, Molecular Cloning and Expression of Laccases—A Divergent Gene Family—in Poplar. FEBS J. 1999, 259, 485–495. [Google Scholar] [CrossRef]
- Turlapati, P.V.; Kim, K.W.; Davin, L.B.; Lewis, N.G. The Laccase Multigene Family in Arabidopsis Thaliana: Towards Addressing the Mystery of Their Gene Function(S). Planta 2011, 3, 439–470. [Google Scholar] [CrossRef] [PubMed]
- Mccaig, B.C.; Meagher, R.B.; Dean, J.F.D. Gene Structure and Molecular Analysis of the Laccase-Like Multicopper Oxidase (Lmco) Gene Family in Arabidopsis Thaliana. Planta 2005, 5, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Caparrós-Ruiz, D.; Fornalé, S.; Civardi, L.; Puigdomènech, P.; Rigau, J. Isolation and Characterisation of a Family of Laccases in Maize. Plant Sci. 2006, 2, 217–225. [Google Scholar] [CrossRef]
- Cesarino, I.; Araújo, P.; Mayer, J.L.S.; Vicentini, R.; Berthet, S.; Demedts, B.; Vanholme, B.; Boerjan, W.; Mazzafera, P. Expression of Soflac, a New Laccase in Sugarcane, Restores Lignin Content but Not S:G Ratio of Arabidopsis Lac17 Mutant. J. Exp. Bot. 2013, 6, 1769–1781. [Google Scholar] [CrossRef]
- Zhang, K.; Lu, K.; Qu, C.; Liang, Y.; Wang, R.; Chai, Y.; Li, J. Gene Silencing of Bntt10 Family Genes Causes Retarded Pigmentation and Lignin Reduction in the Seed Coat of Brassica Napus. PLoS ONE 2013, 4, e61247. [Google Scholar] [CrossRef]
- Wang, Y.; Le, B.P.; Antelme, S.; Soulhat, C.; Gineau, E.; Dalmais, M.; Bendahmane, A.; Morin, H.; Mouille, G.; Lapierre, C. Laccase 5 Is Required for Lignification of the Brachypodium Distachyon Culm. Plant Physiol. 2015, 1, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.Y.; Lee, C.; Hwang, S.G.; Park, Y.C.; Lim, H.L.; Jang, C.S. Overexpression of the Oschi1 Gene, Encoding a Putative Laccase Precursor, Increases Tolerance to Drought and Salinity Stress in Transgenic Arabidopsis. Gene 2014, 1, 98–105. [Google Scholar] [CrossRef]
- Sato, Y.; Bao, W.; Sederoff, R.; Whetten, R. Molecular Cloning and Expression of Eight Laccase Cdnas in Loblolly Pine (Pinus taeda). J. Plant Res. 2001, 2, 147–155. [Google Scholar] [CrossRef]
- O’Malley, D.M.; Ross, W.; Bao, W.; Chen, C.; Sederoff, R.R. The Role of Laccase in Lignification. Plant J. 1993, 5, 751–757. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. Mcscanx: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 7, e49. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Paterson, A.H.; Bowers, J.E.; Chapman, B.A. Ancient Polyploidization Predating Divergence of the Cereals, and Its Consequences for Comparative Genomics. Proc. Natl. Acad. Sci. USA 2004, 26, 9903–9908. [Google Scholar] [CrossRef]
- Toledoortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis Basic/Helix-Loop-Helix Transcription Factor Family. Plant Cell 2003, 8, 1749–1770. [Google Scholar] [CrossRef]
- Guo, Y.; Gan, S. Atnap, a Nac Family Transcription Factor, Has an Important Role in Leaf Senescence. Plant J. 2006, 4, 601–612. [Google Scholar] [CrossRef]
- Shen, H.; Cao, K.; Wang, X. Atbzip16 and Atbzip68, Two New Members of Gbfs, Can Interact with Other G Group Bzips in Arabidopsis Thaliana. BMB Rep. 2008, 2, 132–138. [Google Scholar] [CrossRef]
- Zhou, J.; Lee, C.; Zhong, R.; Ye, Z.H. Myb58 and Myb63 Are Transcriptional Activators of the Lignin Biosynthetic Pathway During Secondary Cell Wall Formation in Arabidopsis. Plant Cell 2009, 1, 248–266. [Google Scholar] [CrossRef]
- Lu, S.; Li, Q.; Wei, H.; Chang, M.J.; Tunlayaanukit, S.; Kim, H.; Liu, J.; Song, J.; Sun, Y.H.; Yuan, L. Ptr-Mir397a Is a Negative Regulator of Laccase Genes Affecting Lignin Content in Populus Trichocarpa. Proc. Natl. Acad. Sci. USA 2013, 26, 10848–10853. [Google Scholar] [CrossRef]
- Abdelghany, S.E.; Pilon, M. Microrna-Mediated Systemic Down-Regulation of Copper Protein Expression in Response to Low Copper Availability in Arabidopsis. J. Biol. Chem. 2008, 23, 15932–15945. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Yu, Y.; Wang, C.Y.; Li, Z.Y.; Liu, Q.; Xu, J.; Liao, J.Y.; Wang, X.J.; Qu, L.H.; Chen, F. Overexpression of Microrna Osmir397 Improves Rice Yield by Increasing Grain Size and Promoting Panicle Branching. Nat. Biotechnol. 2013, 9, 848. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Zhang, S.; Yu, Y.; Luo, Y.-C.; Liu, Q.; Ju, C.; Zhang, Y.-C.; Qu, L.-H.; Lucas, W.J.; Wang, X. Mir397b Regulates Both Lignin Content and Seed Number in Arabidopsis Via Modulating a Laccase Involved in Lignin Biosynthesis. Plant Biotechnol. J. 2015, 8, 1132–1142. [Google Scholar] [CrossRef]
- Bottcher, A.; Cesarino, I.; Santos, A.B.; Vicentini, R.; Mayer, J.L.; Vanholme, R.; Morreel, K.; Goeminne, G.; Moura, J.C.; Nobile, P.M. Lignification in Sugarcane: Biochemical Characterization, Gene Discovery, and Expression Analysis in Two Genotypes Contrasting for Lignin Content. Plant Physiol. 2013, 4, 1539. [Google Scholar] [CrossRef]
- Jin, L.F.; Liu, Y.Z.; Yin, X.X.; Peng, S.A. Transcript Analysis of Citrus Mirna397 and Its Target Lac7 Reveals a Possible Role in Response to Boron Toxicity. Acta Physiol. Plant. 2016, 1, 18. [Google Scholar] [CrossRef]
- Shen, C.; Huang, Y.Y.; He, C.T.; Zhou, Q.; Chen, J.X.; Tan, X.; Mubeen, S.; Yuan, J.G.; Yang, Z.Y. Comparative Analysis of Cadmium Responsive Micrornas in Roots of Two Ipomoea Aquatica Forsk. Cultivars with Different Cadmium Accumulation Capacities. Plant Physiol. Biochem. 2016, 111, 329–339. [Google Scholar] [CrossRef]
- Bo, P.; Hopkins, R.; Rask, L.; Meijer, J. Differential Wound Induction of the Myrosinase System in Oilseed Rape (Brassica napus): Contrasting Insect Damage with Mechanical Damage. Plant Sci. 2005, 3, 715–722. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped Blast and Psi-Blast: A New Generation of Protein Database Search. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Mitchell, A.; Chang, H.Y.; Daugherty, L.; Fraser, M.; Hunter, S.; Lopez, R.; Mcanulla, C.; Mcmenamin, C.; Nuka, G.; Pesseat, S. The Interpro Protein Families Database: The Classification Resource after 15 Years. Nucleic Acids Res. 2015, 43, 213–221. [Google Scholar] [CrossRef]
- Voorrips, R.E. Mapchart: Software for the Graphical Presentation of Linkage Maps and Qtls. J. Hered. 2002, 1, 77–78. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; Von Heijne, G. Predicting Subcellular Localization of Proteins Based on Their N-Terminal Amino Acid Sequence. J. Mol. Biol. 2000, 4, 1005–1016. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. Gsds 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 8, 1296. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. Meme Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. Pssrnaminer: A Plant Short Small Rna Regulatory Cascade Analysis Server. Nucleic Acids Res. 2008, 36, W114–W118. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows–Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Daehwan, K.; Langmead, B.; Salzberg, S.L. Hisat: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 4, 357–360. [Google Scholar]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. Stringtie EnablesImproved Reconstruction of a Transcriptome from Rna-Seq Reads. Nat. Biotechnol. 2015, 3, 290–295. [Google Scholar] [CrossRef]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. Hemi: A Toolkit for Illustrating Heatmaps. PLoS ONE 2014, 11, e111988. [Google Scholar] [CrossRef]
- Lalitha, S. Primer Premier 5. Biotech Softw. Internet Rep. 2000, 1, 270–272. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAC | Gene ID | Predicted Subcellular Location | Chr | Position | Genomic Sequences Length (bp) | cDNA Length (bp) | Protein Sequences Length (aa) | MW (kDa) | pI |
|---|---|---|---|---|---|---|---|---|---|
| BnLAC1-1 | BnaC08g17780D | Secretory | C08 | 21234694:21237623 | 2929 | 1737 | 578 | 65.20 | 9.09 |
| BnLAC1-2 | BnaA08g22770D | Secretory | A08 | 16433641:16436613 | 2972 | 1740 | 579 | 65.23 | 9.09 |
| BnLAC2 | BnaC04g54790D | Secretory | C04 | 2100118: 2102571 | 2453 | 1737 | 578 | 64.46 | 9.53 |
| BnLAC3-1 | BnaA04g17380D | Secretory | A04 | 14120708:14122939 | 2231 | 1713 | 570 | 64.07 | 9.48 |
| BnLAC3-2 | BnaC04g41010D | Secretory | C04 | 41756886:41759146 | 2260 | 1713 | 570 | 64.12 | 9.59 |
| BnLAC3-3 | BnaA05g12170D | Secretory | A05 | 6998148: 7000182 | 2034 | 1632 | 543 | 60.91 | 9.41 |
| BnLAC3-4 | BnaC04g14580D | Secretory | C04 | 11956954:11959010 | 2056 | 1551 | 516 | 57.83 | 9.63 |
| BnLAC4-1 | BnaA05g06610D | Secretory | A05 | 3604302: 3607497 | 3195 | 1677 | 558 | 61.54 | 9.36 |
| BnLAC4-2 | BnaC04g07220D | Secretory | C04 | 5406354: 5409465 | 3111 | 1677 | 558 | 61.54 | 9.36 |
| BnLAC4-3 | BnaA04g21810D | Secretory | A04 | 16546132:16549220 | 3088 | 1680 | 559 | 61.79 | 9.41 |
| BnLAC4-4 | BnaC04g45660D | Secretory | C04 | 45245585:45248722 | 3137 | 1683 | 560 | 61.88 | 9.41 |
| BnLAC5-1 | BnaC04g47080D | Secretory | C04 | 46166799:46169078 | 2279 | 1710 | 569 | 63.00 | 8.57 |
| BnLAC5-2 | BnaA05g05410D | Secretory | A05 | 2791689: 2794263 | 2574 | 1725 | 574 | 63.65 | 8.92 |
| BnLAC5-3 | BnaC04g04810D | Secretory | C04 | 3510057: 3512615 | 2558 | 1725 | 574 | 63.52 | 8.85 |
| BnLAC5-4 | BnaA04g29320D | Secretory | A04 | 1313319: 1320433 | 7114 | 1722 | 573 | 63.47 | 8.57 |
| BnLAC6-1 | BnaA04g27180D | Secretory | A04 | 19140832:19142936 | 2104 | 1710 | 569 | 63.62 | 8.19 |
| BnLAC6-2 | BnaC04g50890D | Secretory | C04 | 48319588:48321669 | 2081 | 1743 | 580 | 65.01 | 8.7 |
| BnLAC7-1 | BnaA05g29170D | Secretory | A05 | 20475705:20479975 | 4270 | 1668 | 555 | 61.16 | 9.11 |
| BnLAC7-2 | BnaCnng24340D | Secretory | Cnn | 22790373:22794992 | 4619 | 1707 | 568 | 62.68 | 9.1 |
| BnLAC8 | BnaC02g03650D | Secretory | C02 | 1747091: 1749618 | 2527 | 1542 | 513 | 57.09 | 8.88 |
| BnLAC9-1 | BnaC03g00490D | Secretory | C03 | 235782: 239591 | 3809 | 1704 | 567 | 62.86 | 6.1 |
| BnLAC9-2 | BnaC03g00480D | Secretory | C03 | 229247: 232215 | 2968 | 1779 | 592 | 65.59 | 6.67 |
| BnLAC9-3 | BnaA03g00500D | Secretory | A03 | 177682: 180567 | 2885 | 1734 | 577 | 63.99 | 6.83 |
| BnLAC10-1 | BnaC02g03710D | Secretory | C02 | 1776151: 1778498 | 2347 | 1680 | 559 | 61.18 | 9.46 |
| BnLAC10-2 | BnaAnng13970D | Secretory | Ann | 15018444:15020836 | 2392 | 1692 | 563 | 61.62 | 9.46 |
| BnLAC11-1 | BnaA10g26680D | Secretory | A10 | 16966690:16969252 | 2562 | 1686 | 561 | 62.54 | 8.99 |
| BnLAC11-2 | BnaC02g03260D | Secretory | C02 | 1549295: 1551691 | 2396 | 1683 | 560 | 62.25 | 8.49 |
| BnLAC11-3 | BnaAnng18410D | Secretory | Ann | 19628923:19631267 | 2344 | 1683 | 560 | 62.14 | 8.75 |
| BnLAC11-4 | BnaCnng02950D | Mitochondrion | Cnn | 2437726: 2440207 | 2481 | 1731 | 576 | 64.31 | 8.88 |
| BnLAC12-1 | BnaAnng01310D | Secretory | Ann | 804829: 807062 | 2233 | 1698 | 565 | 62.58 | 9.52 |
| BnLAC12-2 | BnaA10g25010D | Secretory | A10 | 16204889:16207264 | 2375 | 1698 | 565 | 62.65 | 9.1 |
| BnLAC12-3 | BnaC02g02320D | Secretory | C02 | 1036921: 1039163 | 2242 | 1695 | 564 | 62.50 | 9.49 |
| BnLAC12-4 | BnaC09g49940D | Secretory | C09 | 48045040:48047342 | 2302 | 1698 | 565 | 62.58 | 9.23 |
| BnLAC13-1 | BnaA10g23590D | Secretory | A10 | 15550049:15551986 | 1937 | 1704 | 567 | 63.05 | 6.65 |
| BnLAC13-2 | BnaC09g48310D | Secretory | C09 | 47172850:47174803 | 1953 | 1701 | 566 | 62.86 | 6.65 |
| BnLAC14-1 | BnaC09g47160D | Secretory | C09 | 46664083:46666581 | 2498 | 1731 | 576 | 65.13 | 9.67 |
| BnLAC14-2 | BnaA10g22590D | Secretory | A10 | 15188457:15190992 | 2535 | 1743 | 580 | 65.74 | 9.74 |
| BnLAC15-1 | BnaC02g38340D | Secretory | C02 | 41316880:41322634 | 5754 | 1692 | 563 | 63.60 | 9.01 |
| BnLAC15-2 | BnaAnng08030D | Secretory | Ann | 8068829: 8073341 | 4512 | 1680 | 559 | 63.42 | 9.04 |
| BnLAC15-3 | BnaA06g30430D | Secretory | A06 | 20553666:20556168 | 2502 | 1683 | 560 | 63.26 | 9.09 |
| BnLAC16 | BnaC09g34170D | Secretory | C09 | 37560731:37563386 | 2655 | 1713 | 570 | 63.07 | 9.18 |
| BnLAC17-1 | BnaA03g09140D | Secretory | A03 | 4114497: 4116527 | 2030 | 1722 | 573 | 63.71 | 9.28 |
| BnLAC17-2 | BnaC03g11450D | Secretory | C03 | 5566603: 5568690 | 2087 | 1722 | 573 | 63.82 | 9.32 |
| BnLAC17-3 | BnaA02g06580D | Secretory | A02 | 3141348: 3143631 | 2283 | 1719 | 572 | 63.47 | 9.27 |
| BnLAC17-4 | BnaA10g12900D | Secretory | A10 | 10509237:10513003 | 3766 | 1722 | 573 | 63.59 | 9.28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ping, X.; Wang, T.; Lin, N.; Di, F.; Li, Y.; Jian, H.; Wang, H.; Lu, K.; Li, J.; Xu, X.; et al. Genome-Wide Identification of the LAC Gene Family and Its Expression Analysis Under Stress in Brassica napus. Molecules 2019, 24, 1985. https://doi.org/10.3390/molecules24101985
Ping X, Wang T, Lin N, Di F, Li Y, Jian H, Wang H, Lu K, Li J, Xu X, et al. Genome-Wide Identification of the LAC Gene Family and Its Expression Analysis Under Stress in Brassica napus. Molecules. 2019; 24(10):1985. https://doi.org/10.3390/molecules24101985
Chicago/Turabian StylePing, Xiaoke, Tengyue Wang, Na Lin, Feifei Di, Yangyang Li, Hongju Jian, Hao Wang, Kun Lu, Jiana Li, Xinfu Xu, and et al. 2019. "Genome-Wide Identification of the LAC Gene Family and Its Expression Analysis Under Stress in Brassica napus" Molecules 24, no. 10: 1985. https://doi.org/10.3390/molecules24101985
APA StylePing, X., Wang, T., Lin, N., Di, F., Li, Y., Jian, H., Wang, H., Lu, K., Li, J., Xu, X., & Liu, L. (2019). Genome-Wide Identification of the LAC Gene Family and Its Expression Analysis Under Stress in Brassica napus. Molecules, 24(10), 1985. https://doi.org/10.3390/molecules24101985