Antiproliferative and Enzyme Docking Analysis of Engleromycin from Engleromyces goetzei

Abstract
:1. Introduction
2. Results and Discussion
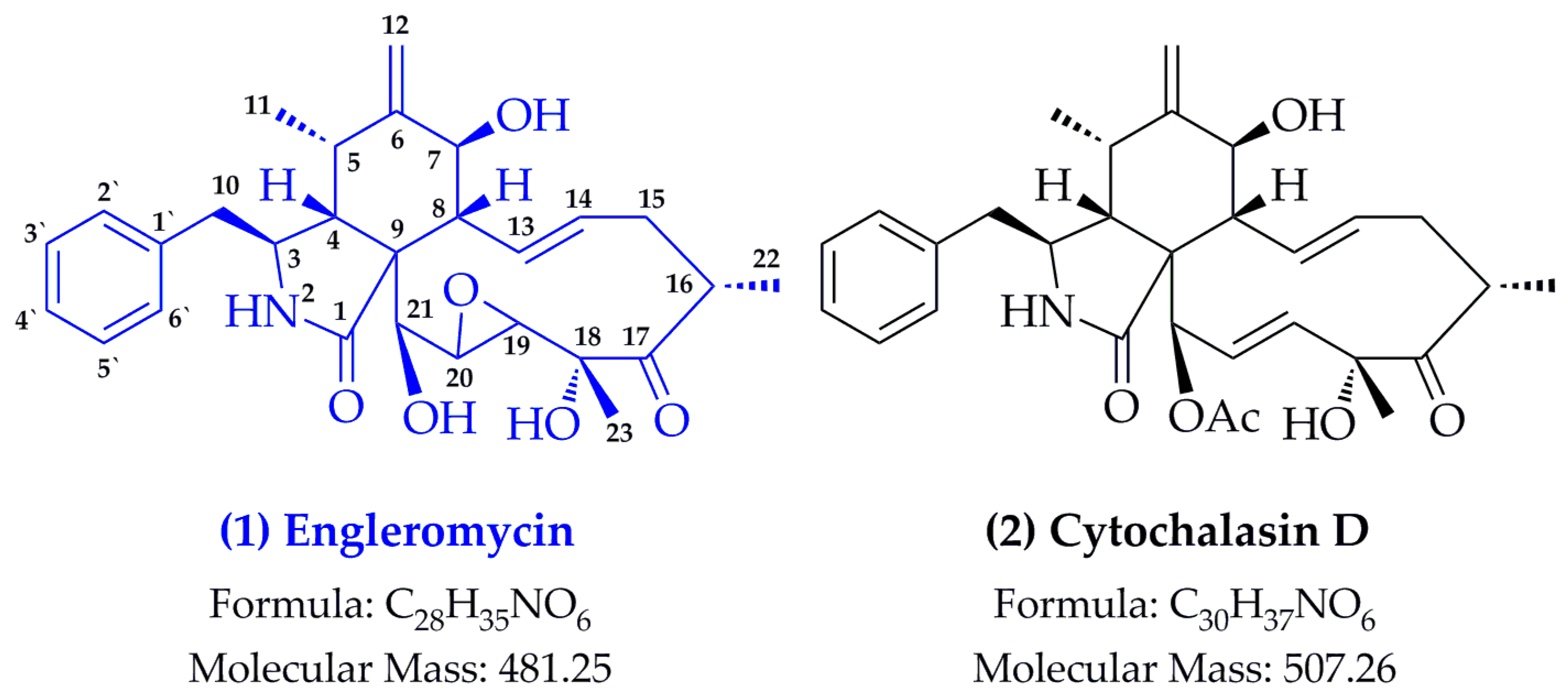
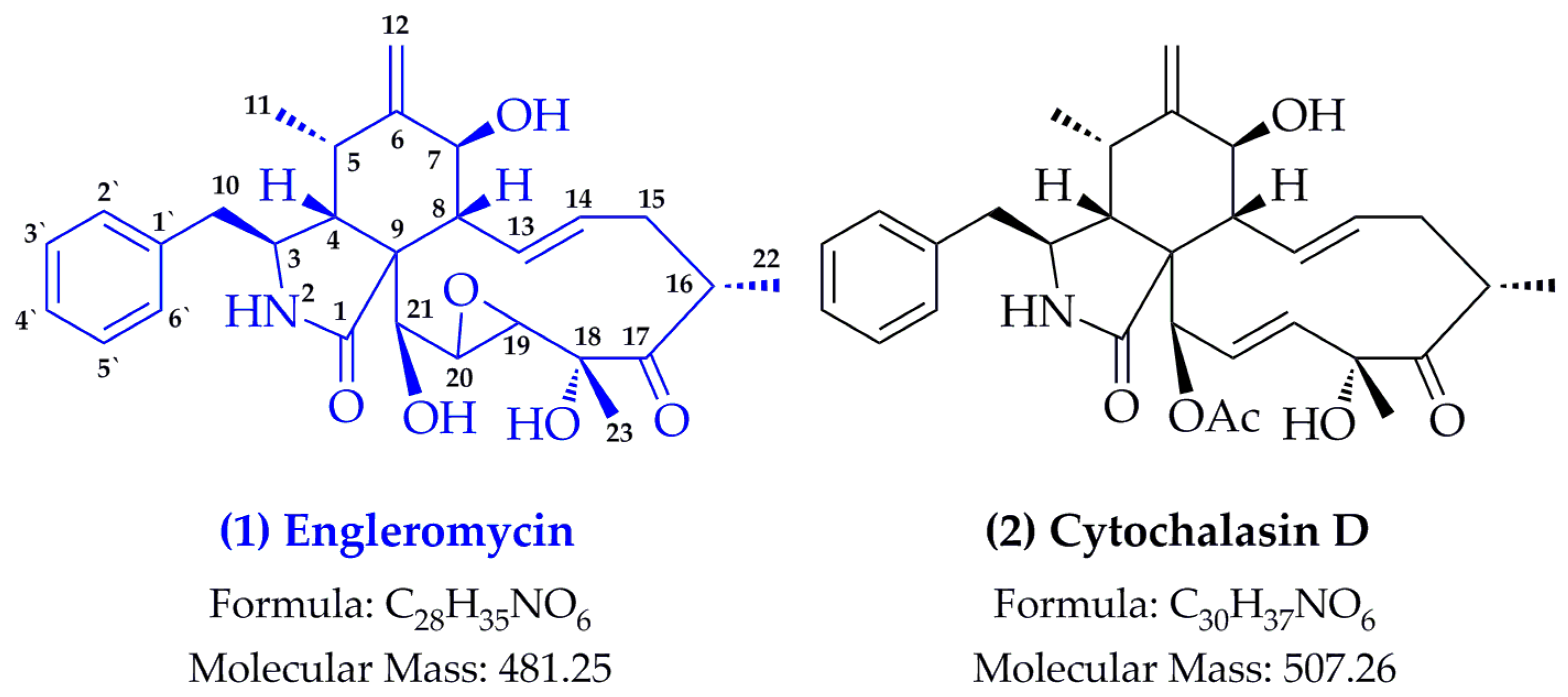
2.1. Structure Identification
2.2. Effects of Engleromycin on Cell Growth of SGC-7901, HT-29, HeLa, A549 and LO-2
2.3. Top II Inhibition Assay In Vitro
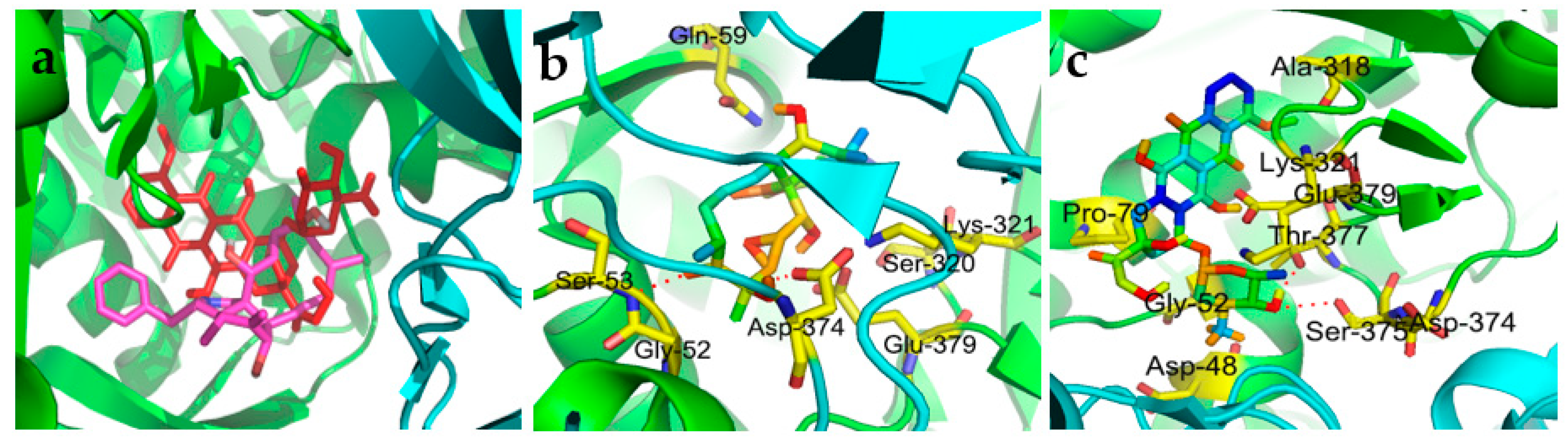
2.4. Molecular Docking Simulation
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Mushroom Material
3.3. Extraction and Isolation
3.4. Antiproliferative Assays
3.5. Top II inhibition Assay In Vitro
3.6. Molecular Docking
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 0, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, E.J.; Larsen, P.; Boll, P.M. Engleromycin, a new cytochalasan from Engleromyces goetzei hennings. Tetrahedron Lett. 1980, 21, 5079–5082. [Google Scholar] [CrossRef]
- Whalley, M.A.; Khalil, A.M.A.; Wei, T.Z.; Yao, Y.J.; Whalley, A.J.S. A new species of Engleromyces from China, a second species in the genus. Mycotaxon 2010, 112, 317–323. [Google Scholar] [CrossRef]
- Kokwaro, J.O. An African knowledge of ethnosystematics and its application to traditional medicine, with particular reference to the medicinal use of the fungus Engleromyces goetzei. Bothalia 1983, 14, 237–243. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Wang, F.; Li, Z.H.; Dong, Z.J.; Liu, J.K. New diterpenes from cultures of the fungus Engleromyces goetzii and their CETP inhibitory activity. Nat. Prod. Bioprospect. 2015, 5, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, L.; Li, G.T.; Li, Z.H.; Dong, Z.J.; Li, Y.; Liu, J.K. Identification and cytotoxic activities of two new trichothecenes and a new cuparane-type sesquiterpenoid from the cultures of the mushroom Engleromyces goetzii. Nat. Prod. Bioprospect. 2015, 5, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S.L.; Liu, X.X.; Wan, X.Y.; Chen, Y.T. Isolation and identification of cytochalasin D from Engleromyces goetzii. Acta Bot. Sin. 1978, 18, 248–252. [Google Scholar]
- Champoux, J.J. DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef]
- Hsieh, T. DNA topoisomerases. Curr. Opin. Cell Biol. 1990, 2, 461–463. [Google Scholar] [CrossRef]
- Wang, J.C. DNA topoisomerases: Why so many? J. Biol. Chem. 1991, 266, 6659–6662. [Google Scholar]
- Giles, G.I.; Sharma, R.P. Topoisomerase enzymes as therapeutic targets for cancer chemotherapy. Med. Chem. 2005, 1, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Nitiss, J.L. Targeting DNA topoisomerase II in cancer chemotherapy. Nat. Rev. Cancer 2009, 9, 338–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pommier, Y. Drugging topoisomerases: Lessons and challenges. ACS Chem. Biol. 2013, 8, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.K.; Escargueil, A.E.; Skladanowski, A. Catalytic topoisomerase II inhibitors in cancer therapy. Pharmacol. Ther. 2003, 99, 167–181. [Google Scholar] [CrossRef]
- Qin, Y.; Meng, L.; Hu, C.; Duan, W.; Zuo, Z.; Lin, L.; Zhang, X.; Ding, J. Gambogic acid inhibits the catalytic activity of human topoisomerase IIα by binding to its ATPase domain. Mol. Cancer Ther. 2007, 6, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K.; Tang, J.W.; Dong, Z.J.; Ding, Z.H.; Wang, X.H.; Liu, P.G. Neoengleromycin, a novel compound from Engleromyces goetzii. Helv. Chim. Acta 2002, 85, 1439–1442. [Google Scholar] [CrossRef]
- Zhan, Z.; Sun, H.; Wu, H.; Yue, J. Chemical components from the fungul Engleromyces goetzei. Acta Bot. Sin. 2003, 45, 248–252. [Google Scholar]
- Cooper, J.A. Effects of cytochalasin and phalloidin on actin. J. Cell Biol. 1987, 105, 1473–1478. [Google Scholar] [CrossRef] [Green Version]
- Yahara, I.; Harada, F.; Sekita, S.; Yoshihira, K.; Natori, S. Correlation between effects of 24 different cytochalasins on cellular structures and cellular events and those on actin in vitro. J. Cell Biol. 1982, 92, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Haidle, A.M.; Myers, A.G. An enantioselective, modular, and general route to the cytochalasins: Synthesis of L-696,474 and cytochalasin B. Proc. Natl. Acad. Sci. USA 2004, 101, 12048–12053. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, S.; Matsushima, T.; Kimura, T.; Minato, H.; Katagiri, K. Zygosporin A, a new antibiotic from Zygosporium masonnii. J. Antibiot. 1968, 21, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Farber, F.E.; Eberle, R. Effects of cytochalasin and alkaloid drugs on biological expression of herpes-simplex virus type 2 DNA. Exp. Cell Res. 1976, 103, 15–22. [Google Scholar] [CrossRef]
- Wagenaar, M.M.; Corwin, J.; Strobel, G.; Clardy, J. Three new cytochalasins produced by an endophytic fungus in the genus Rhinocladiella. J. Nat. Prod. 2000, 63, 1692–1695. [Google Scholar] [CrossRef]
- Malecki, J.M.; Bentke, A.; Ostrowska, B.; Laidler, P. Cytochalasin D, LY294002 and olomoucine synergize in promoting death of melanoma cells through activation of caspase-3 and apoptosis. Melanoma Res. 2010, 20, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Jasinska, A.; Strakova, Z.; Szmidt, M.; Fazleabas, A.T. Human chorionic gonadotropin and decidualization in vitro inhibits cytochalasin-D-induced apoptosis in cultured endometrial stromal fibroblasts. Endocrinology 2006, 147, 4112–4121. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.Y.; Li, Y.N.; Mei, W.L.; Dai, H.F.; Zhou, P.; Tan, G.H. Cytochalasin D, a tropical fungal metabolite, inhibits CT26 tumor growth and angiogenesis. Asian Pac. J. Trop. Med. 2012, 5, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Rubtsova, S.N.; Kondratov, R.V.; Kopnin, P.B.; Chumakov, P.M.; Kopnin, B.P.; Vasiliev, J.M. Disruption of actin microfilaments by cytochalasin D leads to activation of p53. FEBS Lett. 1998, 430, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Binder, M.; Tamm, C. The cytochalasans a new class of biologically active microbial metabolites. Angew. Chem. Int. Edit. 1973, 12, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, S.H.; Hirsch, J.G. Cytocalasin B: Inhibition of D-2-deoxyglucose transport into leukocytes and fibroblasts. Science 1972, 176, 1432–1434. [Google Scholar] [CrossRef] [PubMed]
- Prasain, J.K.; Ueki, M.; Stefanowicz, P.; Osada, H. Rapid screening and identification of cytochalasins by electrospray tandem mass spectrometry. J. Mass. Spectrom. 2002, 37, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Scherlach, K.; Boettger, D.; Remme, N.; Hertweck, C. The chemistry and biology of cytochalasans. Nat. Prod. Rep. 2010, 27, 869–886. [Google Scholar] [CrossRef] [PubMed]
- Nakopoulou, L.; Lazaris, A.C.; Kavantzas, N.; Alexandrou, P.; Athanassiadou, P.; Keramopoulos, A.; Davaris, P. DNA topoisomerase II-alpha immunoreactivity as a marker of tumor aggressiveness in invasive breast cancer. Pathobiology 2000, 68, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, T.; Bates, A. Topo2008: DNA topoisomerases in biology and medicine. Nucleic Acids Res. 2009, 37, 659–660. [Google Scholar] [CrossRef] [PubMed]
- Jun, K.Y.; Park, S.E.; Liang, J.L.; Jahng, Y.; Kwon, Y. Benzo[b]tryptanthrin inhibits MDR1, topoisomerase activity, and reverses adriamycin resistance in breast cancer cells. Chem. Med. Chem 2015, 10, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Rhee, H.K.; Park, H.J.; Lee, S.K.; Lee, C.O.; Choo, H.Y. Synthesis, cytotoxicity, and DNA topoisomerase II inhibitory activity of benzofuroquinolinediones. Bioorg. Med. Chem. 2007, 15, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Wang, Y.; Ma, L.; Ding, M.; Li, T.; Nie, Y.; Gu, Z. Peiminine serves as an adriamycin chemosensitizer in gastric cancer by modulating the EGFR/FAK pathway. Oncol. Rep. 2018, 39, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Tian, Y.Q.; Wu, J.L.; Li, N.; Guo, M.Q. Antiproliferative activities of Amaryllidaceae alkaloids from Lycoris radiata targeting DNA topoisomerase I. Sci. Rep. 2016, 6, 38284–38293. [Google Scholar] [CrossRef]
- Wang, W.; Rupasinghe, S.G.; Schuler, M.A.; Gonzalez de Mejia, E. Identification and characterization of topoisomerase II inhibitory peptides from soy protein hydrolysates. J. Agric. Food Chem. 2008, 56, 6267–6277. [Google Scholar] [CrossRef]
- Christodoulou, M.S.; Calogero, F.; Baumann, M.; Garcia-Argaez, A.N.; Pieraccini, S.; Sironi, M.; Dapiaggi, F.; Bucci, R.; Broggini, G.; Gazzola, S.; et al. Boehmeriasin A as new lead compound for the inhibition of topoisomerases and SIRT2. Eur. J. Med. Chem. 2015, 92, 766–775. [Google Scholar] [CrossRef]
- Rani, S.; Ajeet, A.; Kumar, A. Designing of Sulfanilamide/Sulfacetamide derivatives as human Topoisomerase II Inhibitor: A Docking Approach. Am. J. Pharmacol. Sci. 2014, 2, 42–46. [Google Scholar] [CrossRef]
- Jadhav, A.K.; Karuppayil, S.M. Molecular docking studies on thirteen fluoroquinolines with human topoisomerase II a and b. In Silico Pharmacol. 2017, 5, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Ruthenburg, A.J.; Bechis, S.K.; Verdine, G.L. Nucleotide-dependent domain movement in the ATPase domain of a human type IIA DNA topoisomerase. J. Biol. Chem. 2005, 280, 37041–37047. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |

{kind=link}
{kind=link}
| Compound | IC50 (µM) a | ||||
|---|---|---|---|---|---|
| SGC-7901 | HT-29 | HeLa | A549 | LO-2 | |
| Engleromycin | 26.77 ± 1.69 | 7.73 ± 0.18 | 7.00 ± 0.12 | 3.14 ± 0.03 | 3.76 ± 0.29 |
| Doxorubicin | 0.74 ± 0.03 | 0.24 ± 0.02 | 0.27 ± 0.02 | 0.14 ± 0.01 | 0.58 ± 0.03 |
| Compound | Inhibition of Top II (µM) | Energy Score (kcal/mol) |
|---|---|---|
| Engleromycin | 35.43 ± 1.85 | −16.62 |
| Doxorubicin | 4.92 ± 0.28 | −19.85 |
| Etoposide [3] | 78.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, G.; Ma, H.; Guo, M. Antiproliferative and Enzyme Docking Analysis of Engleromycin from Engleromyces goetzei. Molecules 2019, 24, 166. https://doi.org/10.3390/molecules24010166
Zhang Y, Chen G, Ma H, Guo M. Antiproliferative and Enzyme Docking Analysis of Engleromycin from Engleromyces goetzei. Molecules. 2019; 24(1):166. https://doi.org/10.3390/molecules24010166
Chicago/Turabian StyleZhang, Yongli, Guilin Chen, Hong Ma, and Mingquan Guo. 2019. "Antiproliferative and Enzyme Docking Analysis of Engleromycin from Engleromyces goetzei" Molecules 24, no. 1: 166. https://doi.org/10.3390/molecules24010166
APA StyleZhang, Y., Chen, G., Ma, H., & Guo, M. (2019). Antiproliferative and Enzyme Docking Analysis of Engleromycin from Engleromyces goetzei. Molecules, 24(1), 166. https://doi.org/10.3390/molecules24010166