Crinipellins A and I, Two Diterpenoids from the Basidiomycete Fungus Crinipellis rhizomaticola, as Potential Natural Fungicides

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. In Vivo Antifungal Activity of the Culture Filtrate of the IUM00035 Isolate
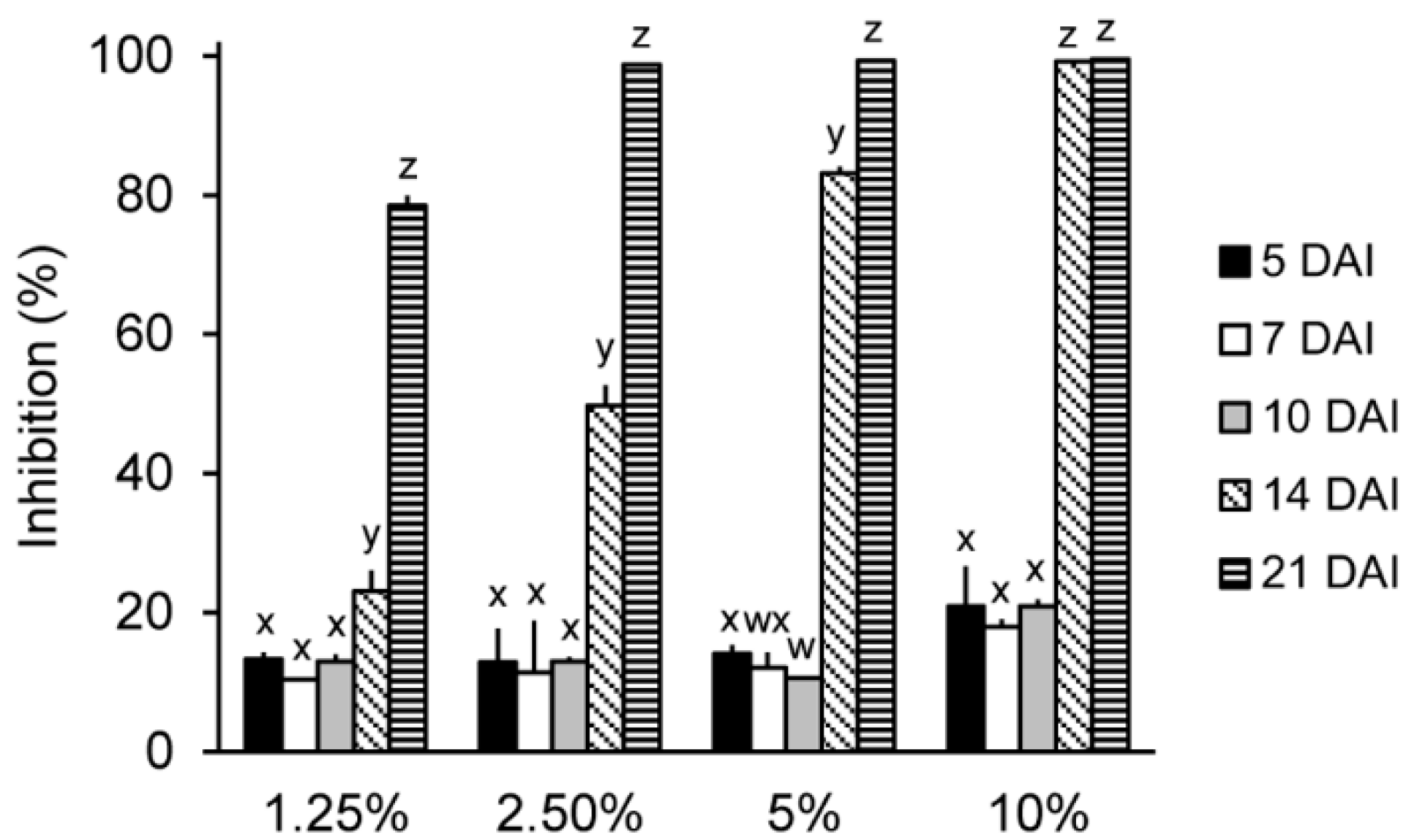
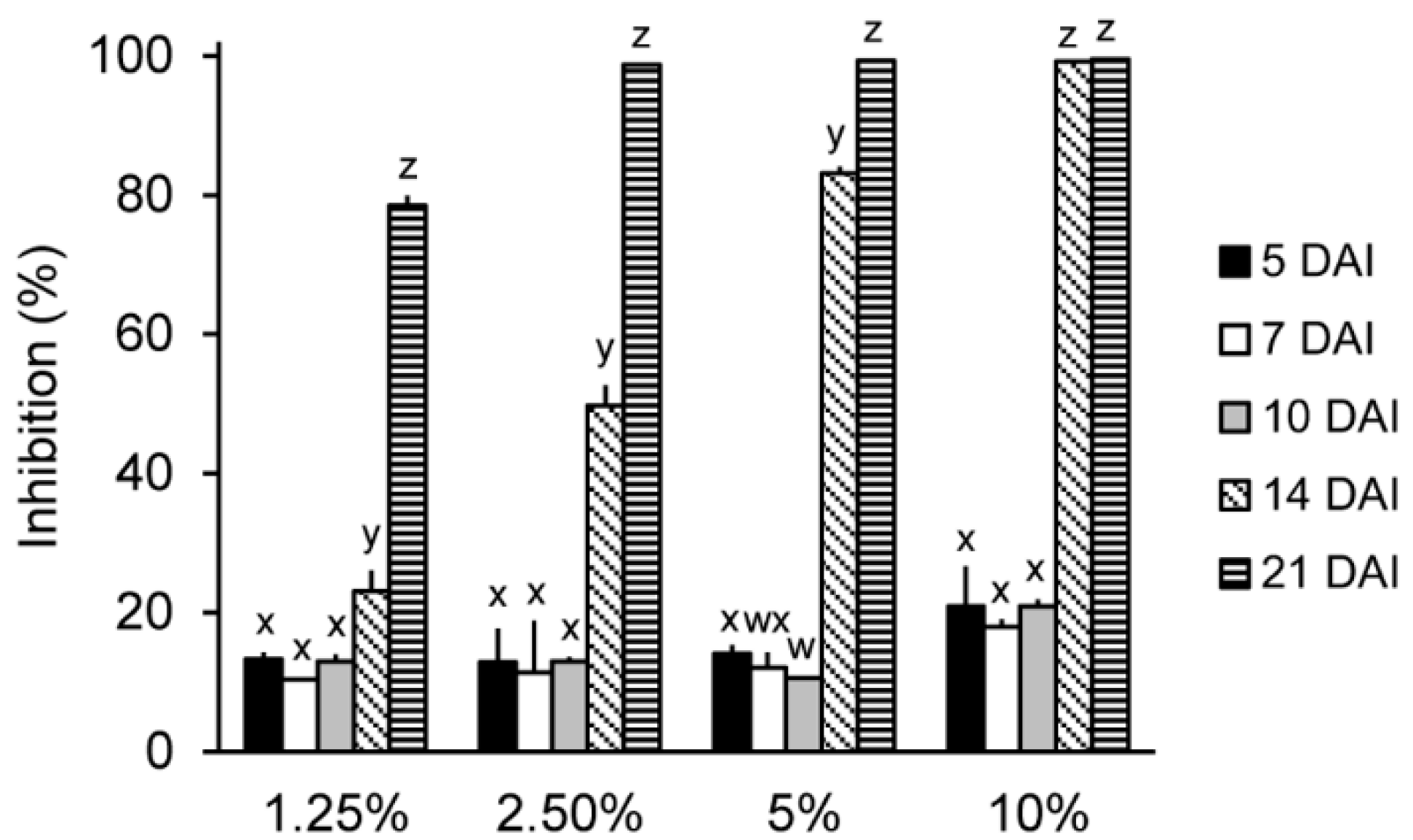
2.2. In Vitro Antifungal Activity of the IUM00035 Culture According to Incubation Period
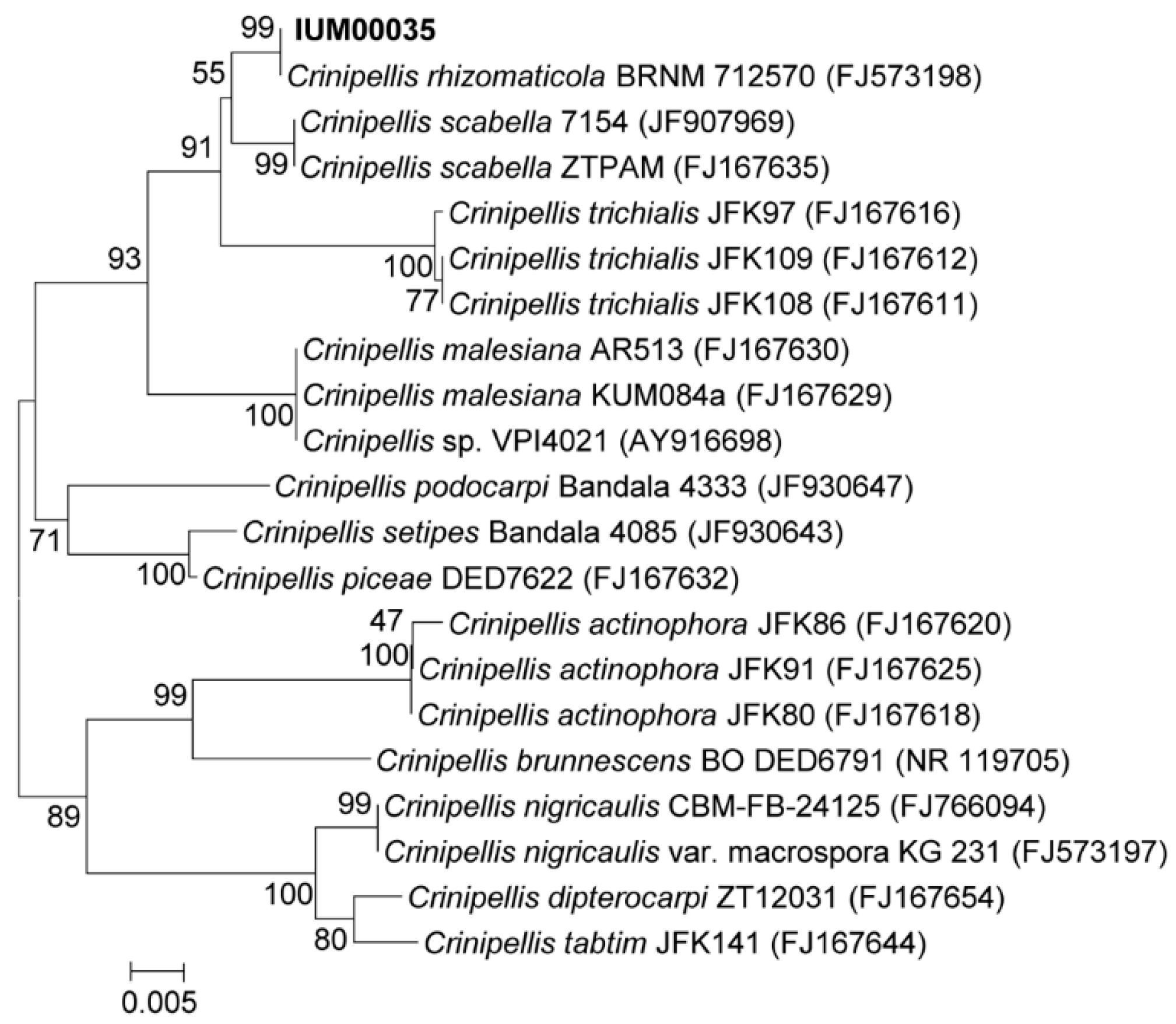
2.3. Molecular Identification of the IUM00035 Isolate
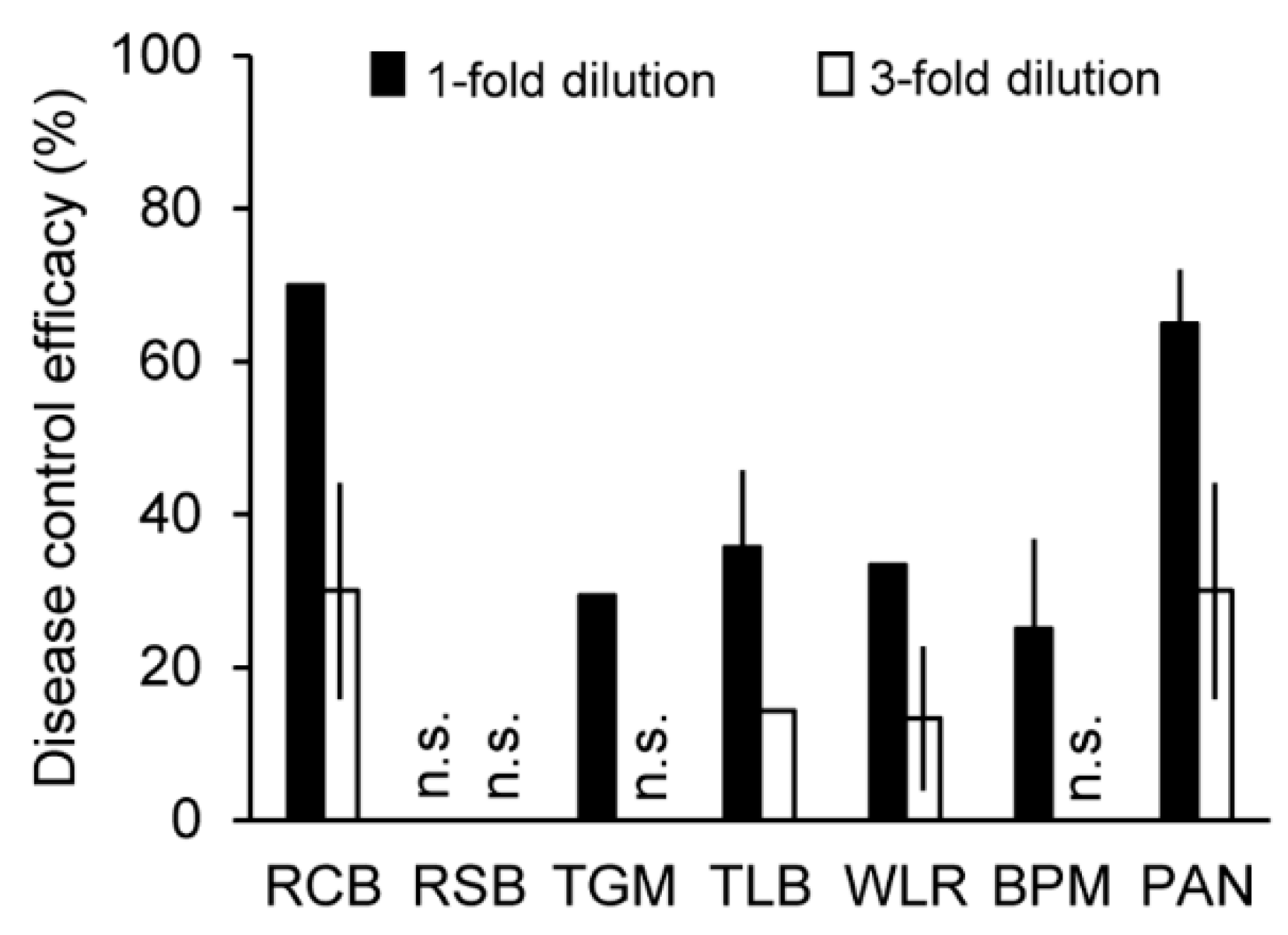
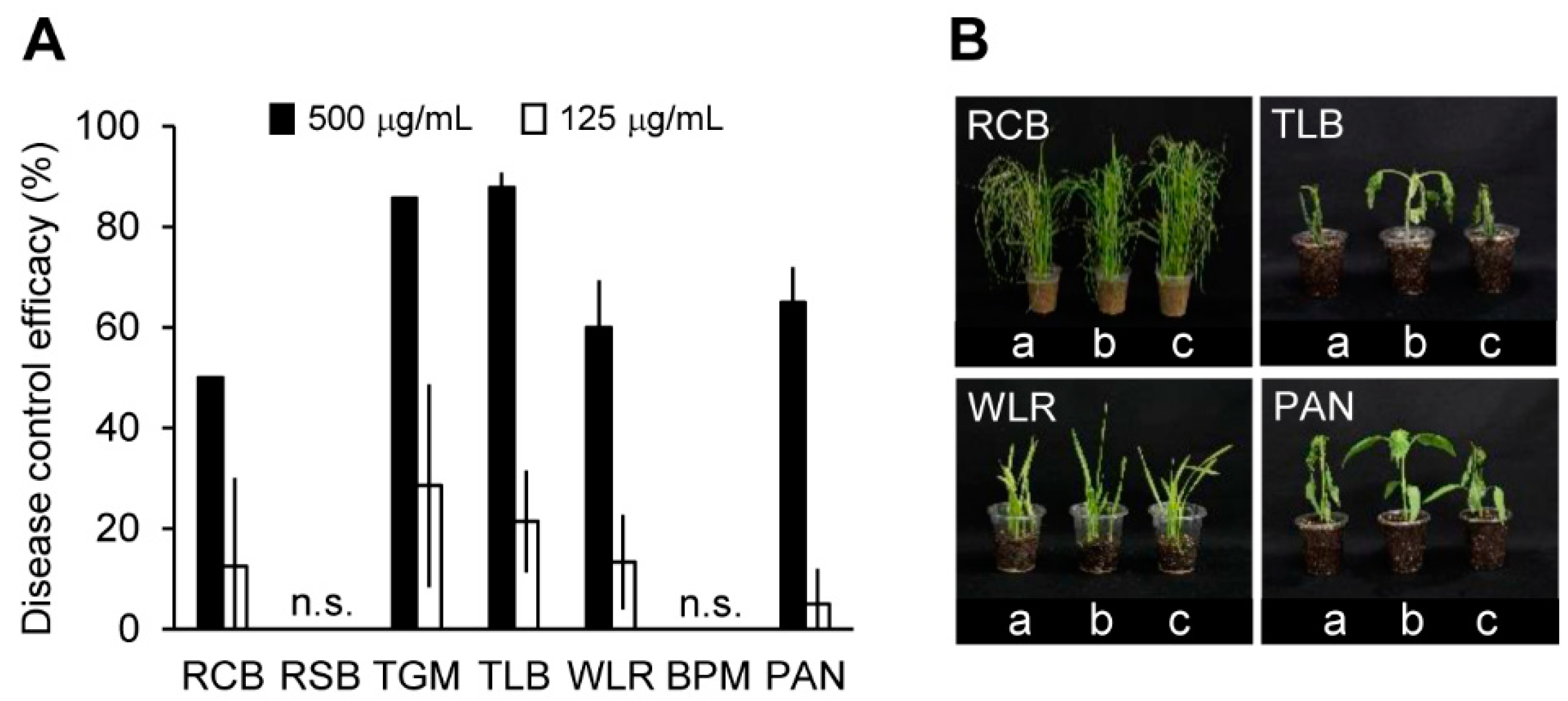
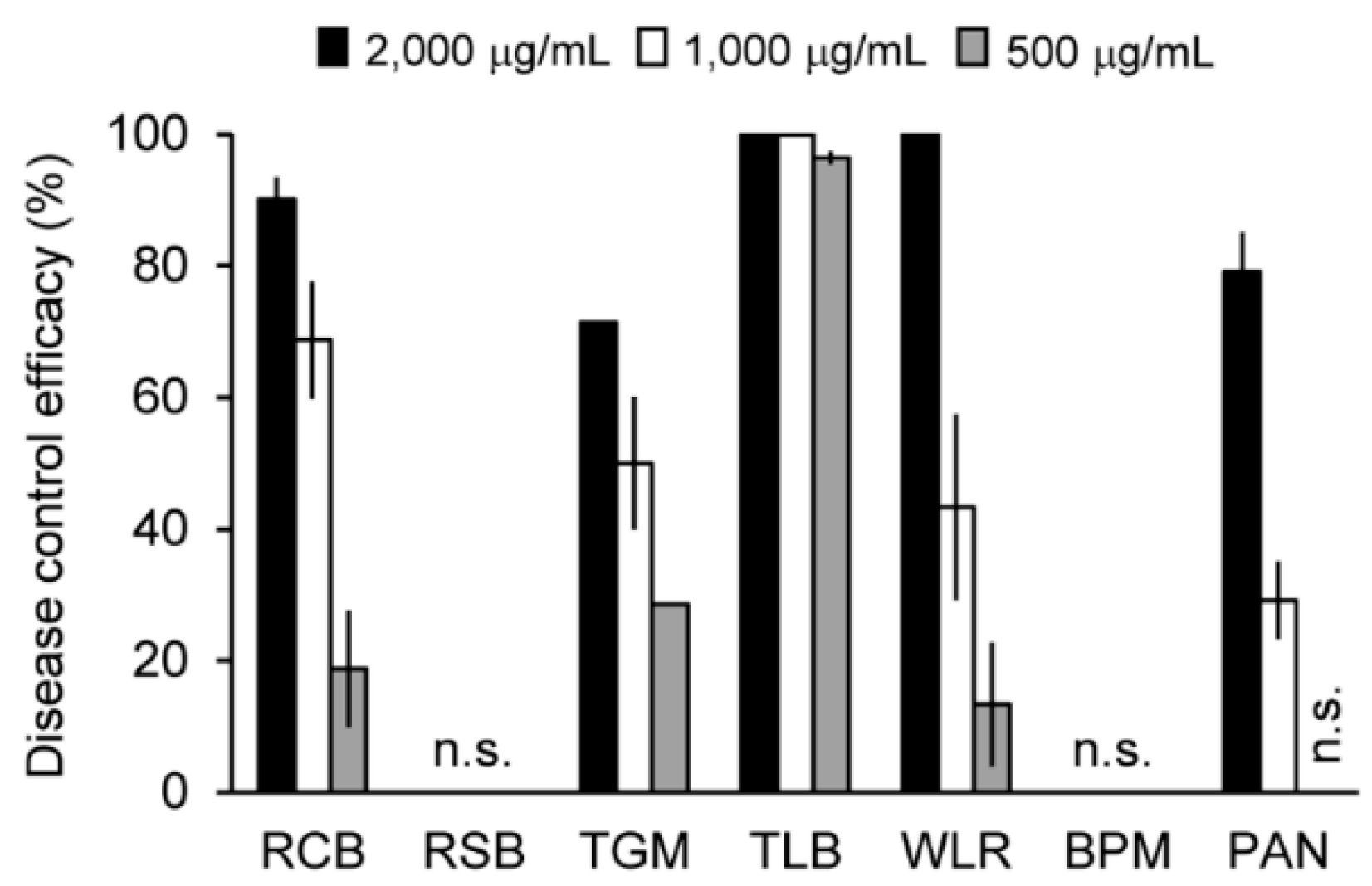
2.4. In Vivo Antifungal Activity of the Solvent Extracts of the C. rhizomaticola Culture Filtrate
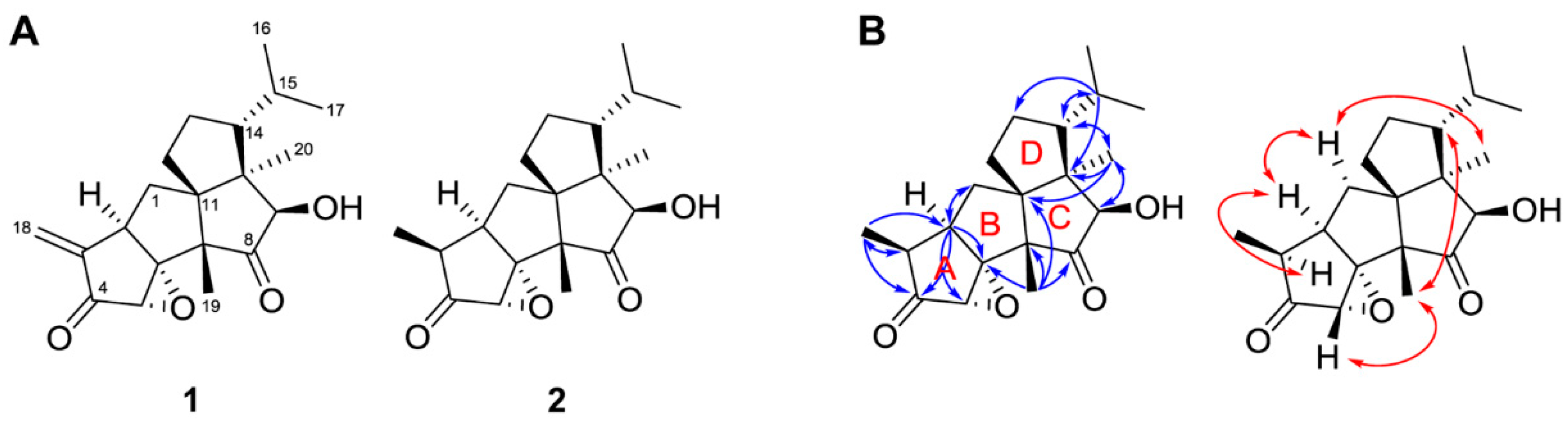
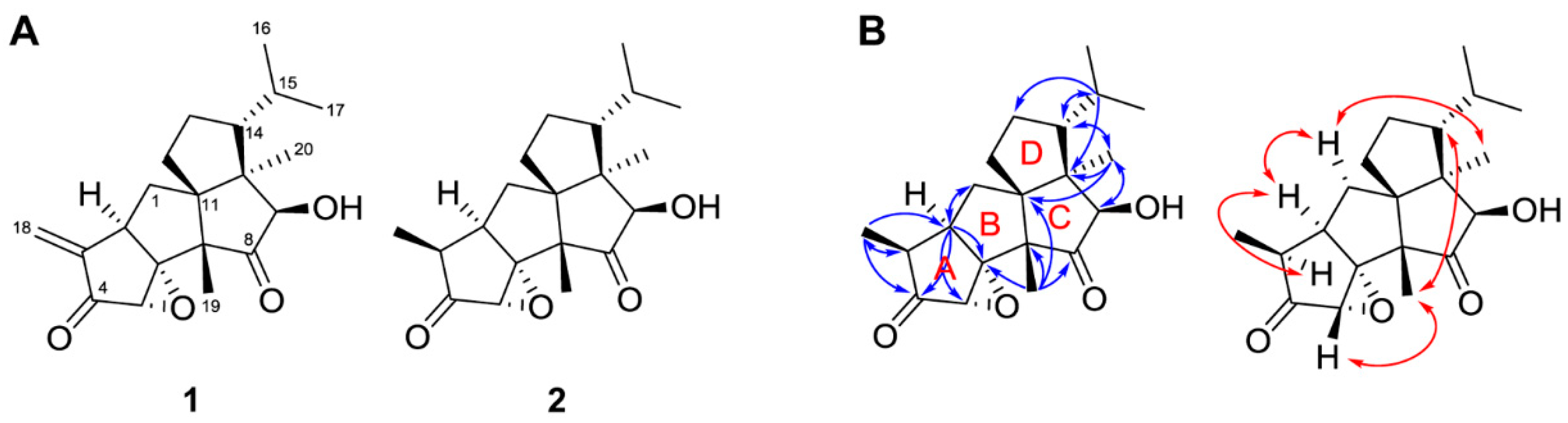
2.5. Isolation and Structure Determination of the Antifungal Compounds
2.6. Minimum Inhibitory Concentrations (MICs) of Crinipellins A and I
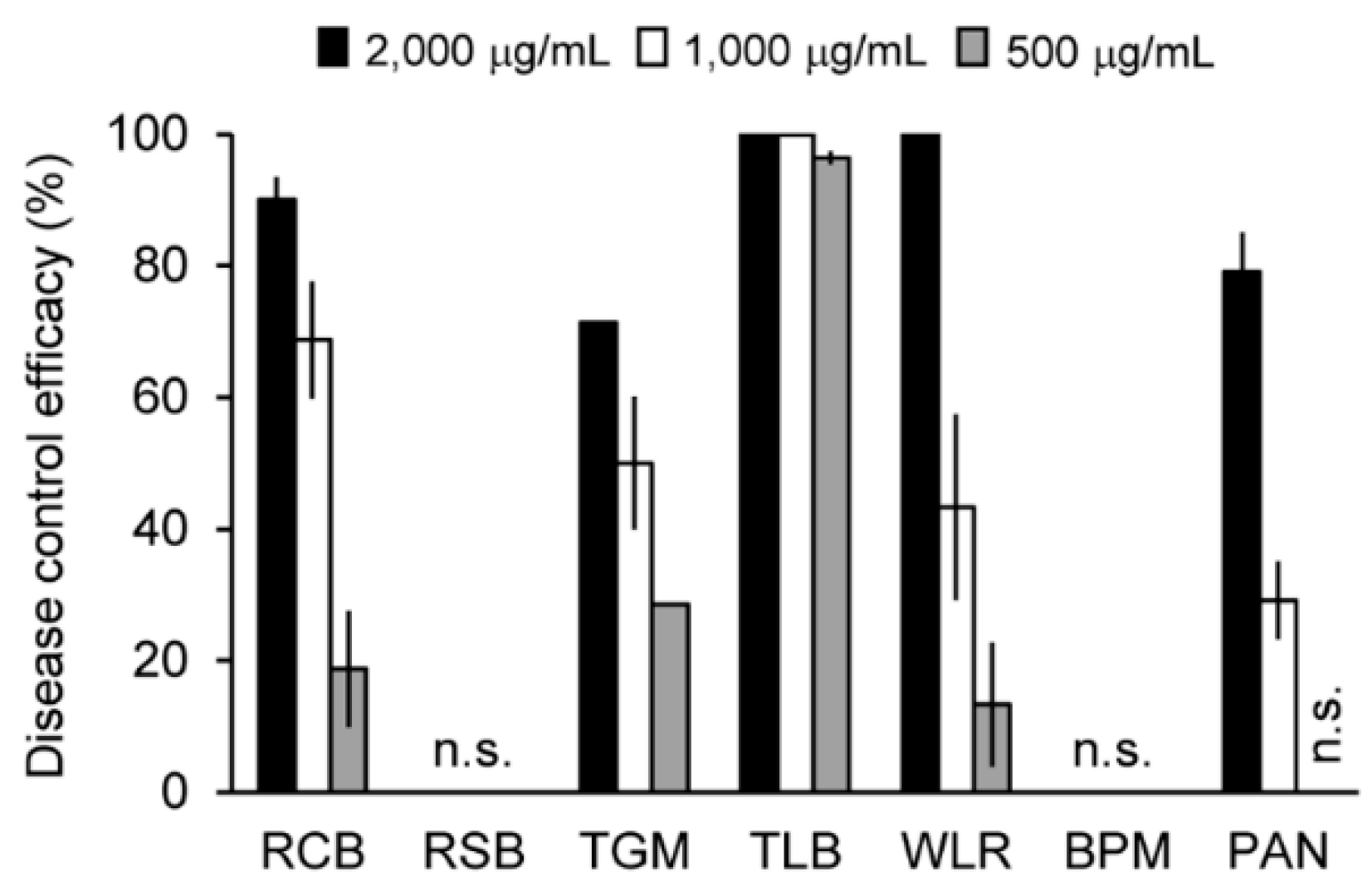
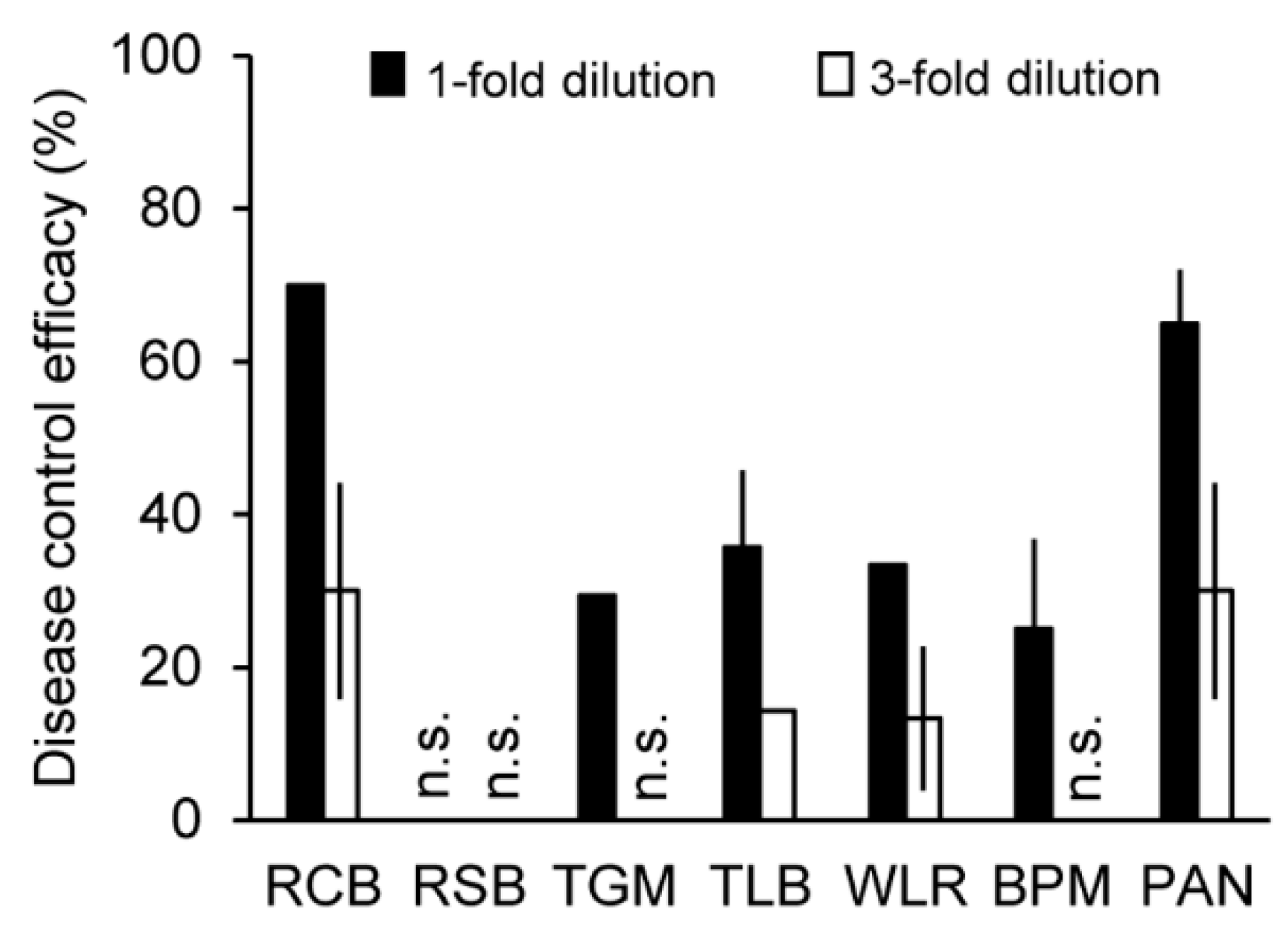
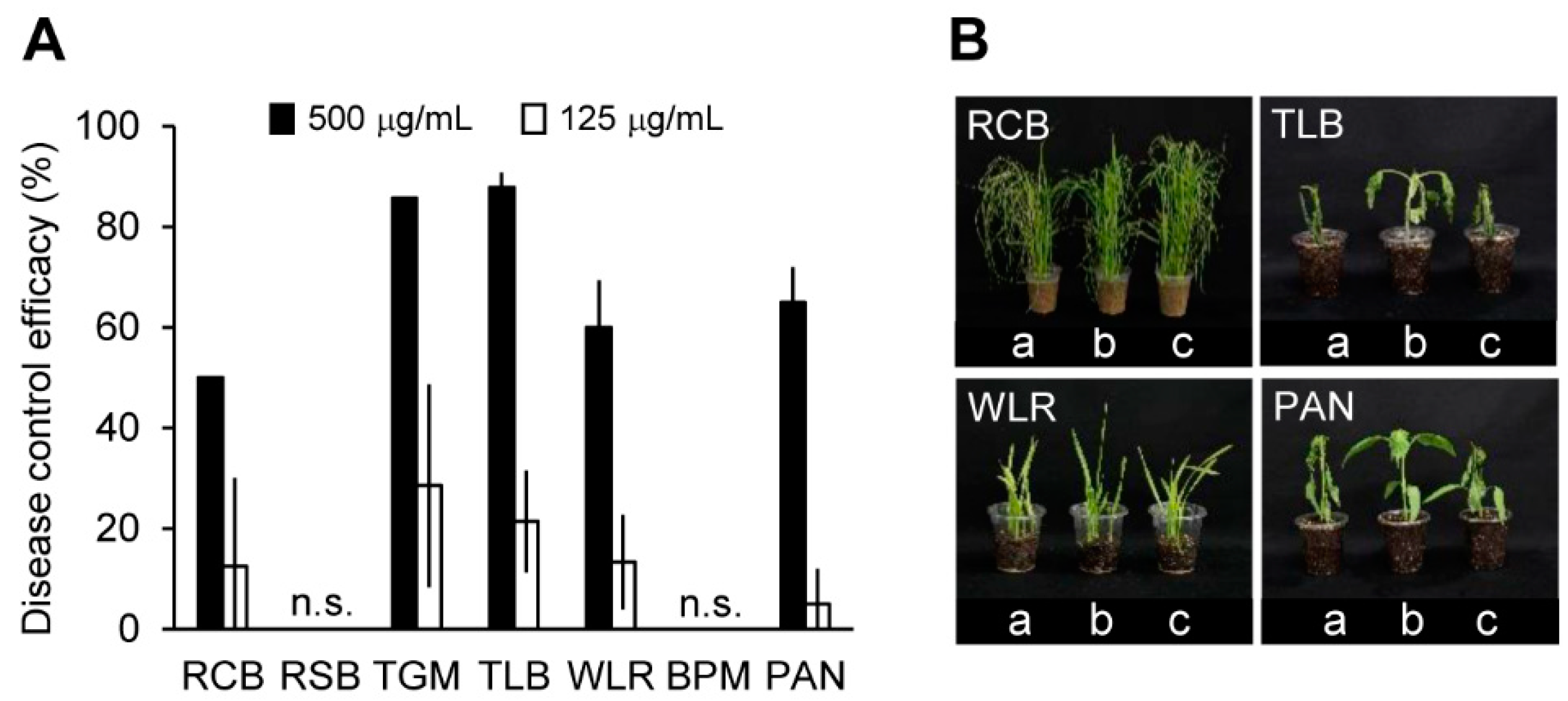
2.7. Suppressive Effects of Crinipellin A (1) on Plant Diseases Caused by Plant-Pathogenic Fungi
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Mushrooms and Culture Conditions
3.3. Genomic DNA Extraction and Phylogenetic Analysis
3.4. Isolation of Antifungal Compounds
3.4.1. Crinipellin A (1)
3.4.2. Crinipellin I (2)
3.5. Optimization of the Incubation Time for the Production of Antifungal Substances
3.6. In Vitro Antifungal Activity Assay against Plant Pathogens
3.7. Disease Control Efficacy Test against Plant Pathogens
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Hüter, O.F. Use of natural products in the crop protection industry. Phytochem. Rev. 2011, 10, 185–194. [Google Scholar] [CrossRef]
- Centrell, C.L.; Dayan, F.E.; Duke, S.O. Natural products as sources for new pesticides. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Lindequist, U.; Niedermeyer, T.H.; Jülich, W.D. The pharmacological potential of mushrooms. Evid-Based Complement. Altern. Med. 2005, 2, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Poucheret, P.; Fons, F.; Rapior, S. Biological and pharmacological activity of higher fungi: 20-year retrospective analysis. Cryptogam. Mycol. 2006, 27, 311–333. [Google Scholar]
- Wasser, S.P. Medicinal mushroom science: History, current status, future trends, and unsolved problems. Int. J. Med. Mushrooms 2010, 12, 1–16. [Google Scholar] [CrossRef]
- Hatvani, N. Antibacterial effect of the culture fluid of Lentinus edodes mycelium grown in submerged liquid culture. Int. J. Antimicrob. Agents 2001, 17, 71–74. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T.B. Eryngin, a novel antifungal peptide from fruiting bodies of the edible mushroom Pleurotus eryngii. Peptides 2004, 25, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E. Biological activities of the polysaccharides produced in submerged culture of two edible Pleurotus ostreatus mushrooms. BioMed Res. Int. 2012, 2012, 565974. [Google Scholar]
- Lorenzen, K.; Anke, T. Basidiomycetes as a source for new bioactive natural products. Curr. Org. Chem. 1998, 2, 329–364. [Google Scholar]
- Zhou, Y.; Bian, Q.; Yang, P.; Wang, L.; Li, S.; Sun, X.; Wang, M.; Wang, M.; Zhong, J. Catalytic asymmetric syntheses of (-)-oudemanisin A and its diastereomer. Tetrahedron 2017, 28, 969–973. [Google Scholar] [CrossRef]
- Anke, T.; August, P.R.; Bertels, S.; Bindseil, K.U.; Blondelle, S.E.; Fischer, G.; Floss, H.G.; Freitag, R.; Frormann, S.; Geschwill, K.; et al. Strobilurins and Oudemansins. In Drug Discovery from Nature, 1st ed.; Anke, T., Strglich, W., Eds.; Springer: Berlin, Germany, 1999; Volume 18, pp. 320–323. ISBN 3-540-64844-5. [Google Scholar]
- Imtiaj, A.; Lee, T.S. Screening of antibacterial and antifungal activities from Korean wild mushrooms. World J. Agric. Sci. 2007, 3, 316–321. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 177. ISBN 978-0-85199-826-8. [Google Scholar]
- Antonín, V.; Ryoo, R.; Shin, H.D. Marasmioid and gymnopoid fungi of the Republic of Korea. 1. Three interesting species of Crinipellis (Basidiomycota, Marasmiaceae). Mycotaxon 2009, 108, 429–440. [Google Scholar] [CrossRef]
- Piers, E.; Renaud, J. Total synthesis of the tetraquinane diterpenoid (±)-crinipellin B. J. Org. Chem. 1993, 58, 11–13. [Google Scholar] [CrossRef]
- Kang, T.; Song, S.B.; Kim, W.Y.; Kim, B.G.; Lee, H.Y. Total synthesis of (-)-crinipellin A. J. Am. Chem. Soc. 2014, 136, 10274–10276. [Google Scholar] [CrossRef] [PubMed]
- Anke, T.; Heim, J.; Knoch, F.; Mocek, U.; Steffan, B.; Steglich, W. Crinipellins, the first natural products with a tetraquinane skeleton. Angew. Chem. Int. Ed. 1985, 24, 709–711. [Google Scholar] [CrossRef]
- Rohr, M.; Oleinikov, K.; Jung, M.; Sandjo, L.P.; Opatz, T.; Erkel, G. Anti-inflammatory tetraquinane diterpenoids from a Crinipellis species. Bioorg. Med. Chem. 2017, 25, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Kupka, J.; Anke, T.; Oberwinkler, F.; Schramm, G.; Steglich, W. Antibiotics from basidiomycetes. VII. Cripellin, a new antibiotic from the basidiomycetous fungus Cripellis Stipitaria (Fr.) PAT. J. Antibiot. 1979, 32, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Shen, Y.M. Four novel diterpenoids from Crinipellis sp. 113. Helv. Chim. Acta 2010, 93, 2151–2157. [Google Scholar] [CrossRef]
- Oh, M.; Son, H.; Choi, G.J.; Lee, C.; Kim, J.-C.; Kim, H.; Lee, Y.-W. Transcription factor ART1 mediates starch hydrolysis and mycotoxin production in Fusarium graminearum and F. verticillioides. Mol. Plant. Pathol. 2016, 17, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.T.; Kim, H.; Tran, V.K.; Dang, Q.L.; Nguyen, H.T.; Kim, H.; Kim, I.S.; Choi, G.J.; Kim, J.C. In vitro antibacterial activity of selected medicinal plants traditionally used in Vietnam against human pathogenic bacteria. BMC Complement. Altern. Med. 2016, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Han, J.W.; Shim, S.H.; Jang, K.S.; Choi, Y.H.; Kim, H.; Choi, G.J. In vivo disease control efficacy of isoquinoline alkaloids isolated from Corydalis ternata against wheat leaf rust and pepper anthracnose. J. Microbiol. Biotechnol. 2018, 28, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Han, J.W.; Shim, S.H.; Jang, K.S.; Choi, Y.H.; Dang, L.Q.; Kim, H.; Choi, G.J. In vivo assessment of plant extracts for control of plant diseases: A sesquiterpene ketolactone isolated from Curcuma zedoaria suppresses wheat leaf rust. J. Environ. Sci. Health Part B 2018, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compound 1 and 2 are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | 2 |
|---|---|---|
| δH, (J in Hz) | δH, (J in Hz) | |
| 1 | 2.51, dd (14.3, 7.5); 1.39, dd (14.2, 13.0) | 2.23, dd (14.0, 6.6); 1.20 dd (14.4, 13.7) |
| 2 | 3.08, dd (12.5, 7.7) | 2.69 dt (13.7, 7.0) |
| 3 | - | 2.55 p (7.3) |
| 5 | 3.46, s | 3.29, s |
| 9 | 4.40, s | 4.42, d (2.7) |
| 12 | 1.87, dt (9.9, 7.1); 1.59, m | 1.88, m; 1.63, m |
| 13 | 1.59, m | 1.63, m |
| 14 | 1.36, m | 1.37, dd (9.3, 2.5) |
| 15 | 2.09, pd (7.0, 2.8) | 2.12, qd (6.8, 2.7) |
| 16 | 0.85, d (7.0) | 0.89, d (6.9) |
| 17 | 0.82, d (6.8) | 0.86, d (6.9) |
| 18 | 5.48, d (1.4); 6.13, d (1.8) | 1.03, d (6.3) |
| 19 | 1.02, s | 1.02, s |
| 20 | 1.31, s | 1.30, s |
| Position | δC, Type | δH, (J in Hz) | HMBC |
|---|---|---|---|
| 1 | 33.6, CH2 | 2.18, dd (13.9, 6.8); 1.21, m | C-2, 3, 6, 7, 10, 11 |
| 2 | 40.4, CH | 2.74, dt (13.9, 7.2) | C-1, 4, 5, 6 |
| 3 | 42.0, CH | 2.45, p (7.4) | C-2, 4, 18 |
| 4 | 212.4, CO | - | |
| 5 | 58.4, CH | 3.29, s | C-4, 6 |
| 6 | 79.4, C | - | |
| 7 | 51.7, C | - | |
| 8 | 216.6, CO | - | |
| 9 | 86.1, CH | 4.38, d (2.8) | C-8, 10, 20 |
| 10 | 54.6, C | ||
| 11 | 61.5, C | ||
| 12 | 33.2, CH2 | 1.89, ddd (12.3, 8.8, 3.9); 1.59, m | C-7, 10, 11, 13 |
| 13 | 23.5, CH2 | 1.59, m | |
| 14 | 52.7, CH | 1.38, td (9.5, 2.5) | C-9, 10, 15, 16, 17, 20 |
| 15 | 29.0, CH | 2.09, pd (6.9, 2.5) | C-10, 13, 14, 16, 17 |
| 16 | 25.2, CH3 | 0.84, d (5.4) | C-14, 15, 17 |
| 17 | 19.8, CH3 | 0.83, d (5.6) | C-16 |
| 18 | 9.5, CH3 | 0.94, d (7.4) | C-2, 3, 4 |
| 19 | 15.4, CH3 | 0.92, s | C-6, 7, 8, 11 |
| 20 | 16.2, CH3 | 1.21, s | C-9, 10, 11, 14 |
| Plant-Pathogenic Fungi | MIC (µg/mL) | |
|---|---|---|
| 1 | 2 | |
| Alternaria porri | 125 | 250 |
| Botrytis cinerea | 31 | >250 |
| Colletotrichum coccodes | 1 | 250 |
| Fusarium oxysporum | 125 | >250 |
| Magnaporthe oryzae | 8 | 250 |
| Phytophthora infestans | 31 | 250 |
| Rhizoctonia solani | 125 | >250 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.W.; Oh, M.; Lee, Y.J.; Choi, J.; Choi, G.J.; Kim, H. Crinipellins A and I, Two Diterpenoids from the Basidiomycete Fungus Crinipellis rhizomaticola, as Potential Natural Fungicides. Molecules 2018, 23, 2377. https://doi.org/10.3390/molecules23092377
Han JW, Oh M, Lee YJ, Choi J, Choi GJ, Kim H. Crinipellins A and I, Two Diterpenoids from the Basidiomycete Fungus Crinipellis rhizomaticola, as Potential Natural Fungicides. Molecules. 2018; 23(9):2377. https://doi.org/10.3390/molecules23092377
Chicago/Turabian StyleHan, Jae Woo, Mira Oh, Yu Jeong Lee, Jaehyuk Choi, Gyung Ja Choi, and Hun Kim. 2018. "Crinipellins A and I, Two Diterpenoids from the Basidiomycete Fungus Crinipellis rhizomaticola, as Potential Natural Fungicides" Molecules 23, no. 9: 2377. https://doi.org/10.3390/molecules23092377
APA StyleHan, J. W., Oh, M., Lee, Y. J., Choi, J., Choi, G. J., & Kim, H. (2018). Crinipellins A and I, Two Diterpenoids from the Basidiomycete Fungus Crinipellis rhizomaticola, as Potential Natural Fungicides. Molecules, 23(9), 2377. https://doi.org/10.3390/molecules23092377