Optimization of Oligosaccharide Production from Leuconostoc lactis Using a Response Surface Methodology and the Immunostimulating Effects of These Oligosaccharides on Macrophage Cells


Abstract
:1. Introduction
2. Results and Discussion
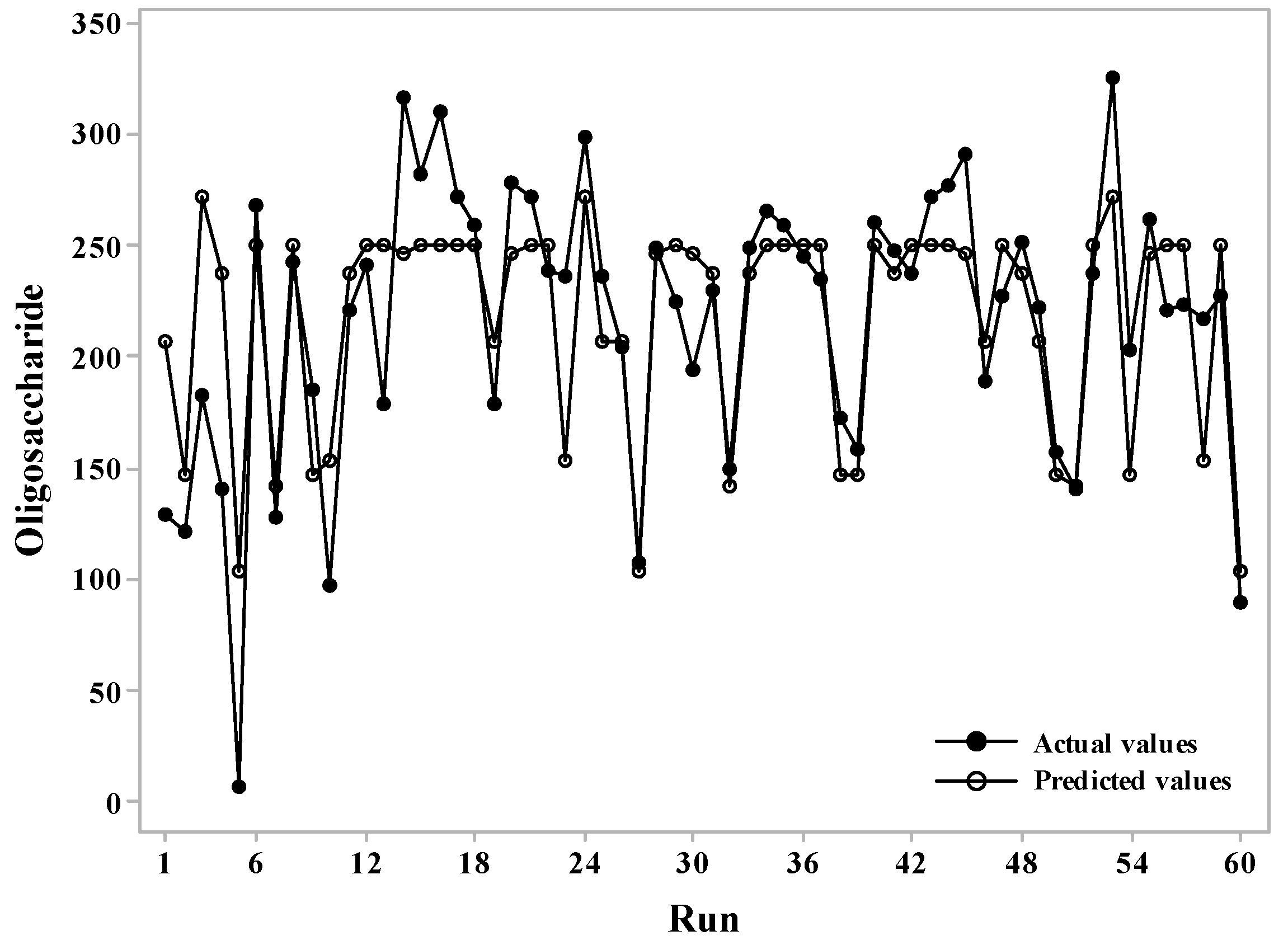
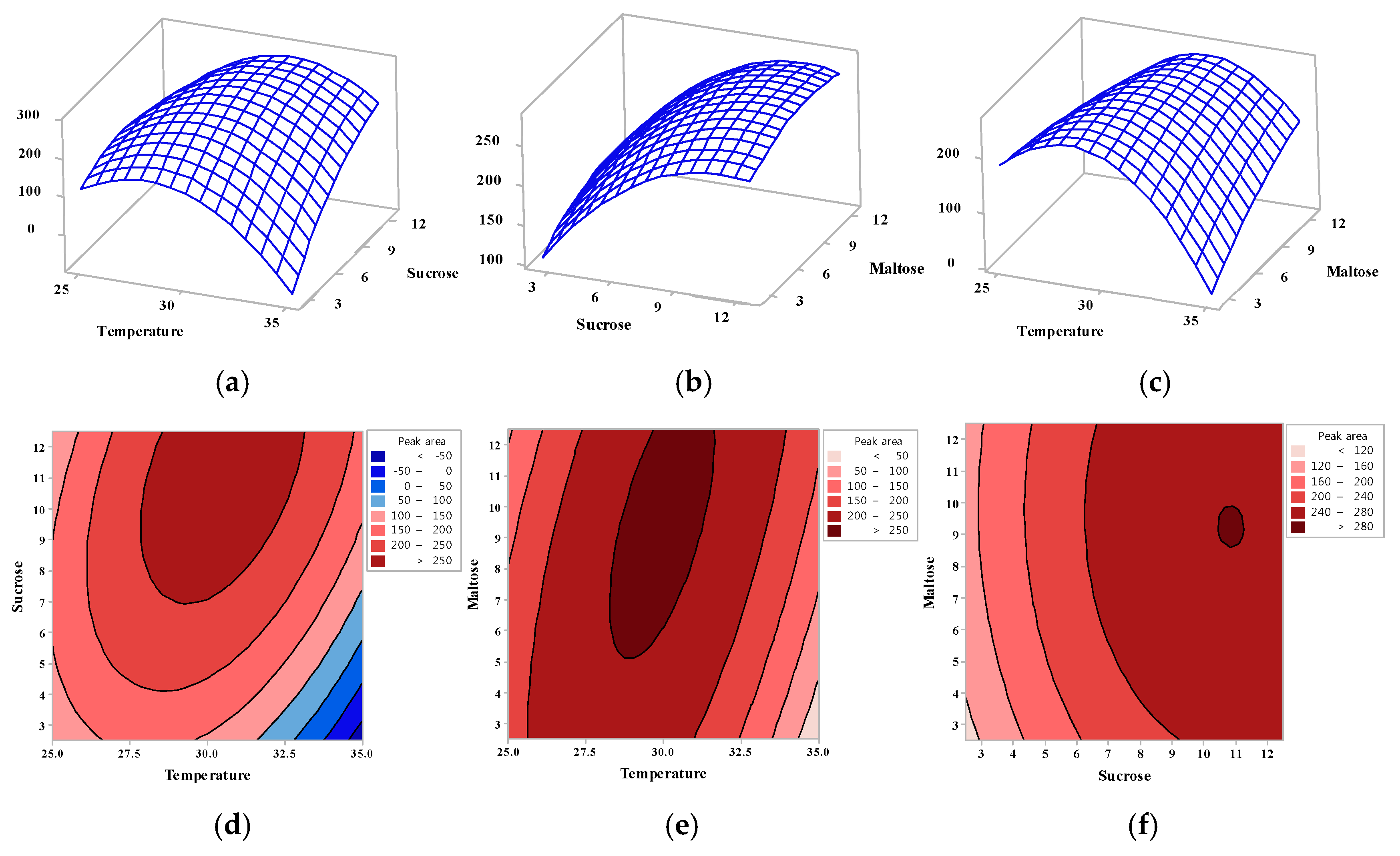
2.1. Optimization of Oligosaccharide Production Using a Statistical Technique
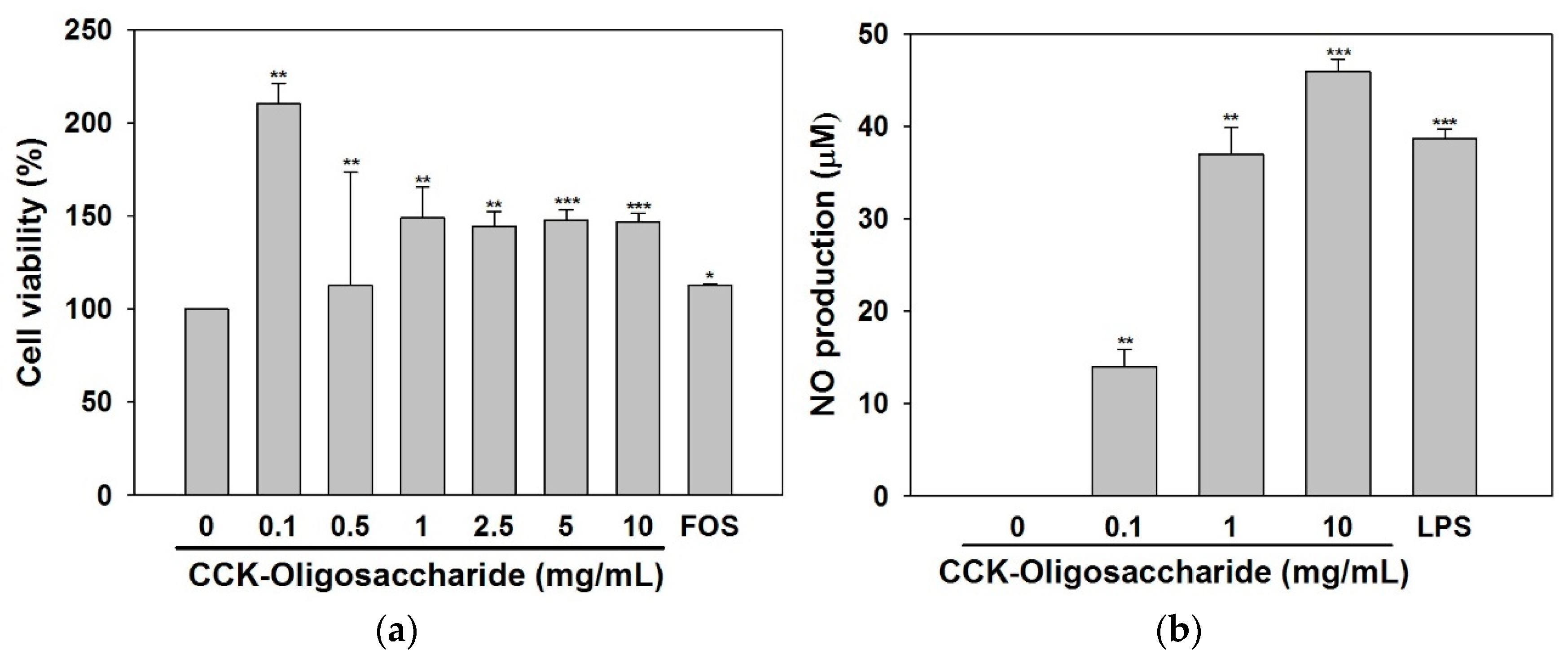
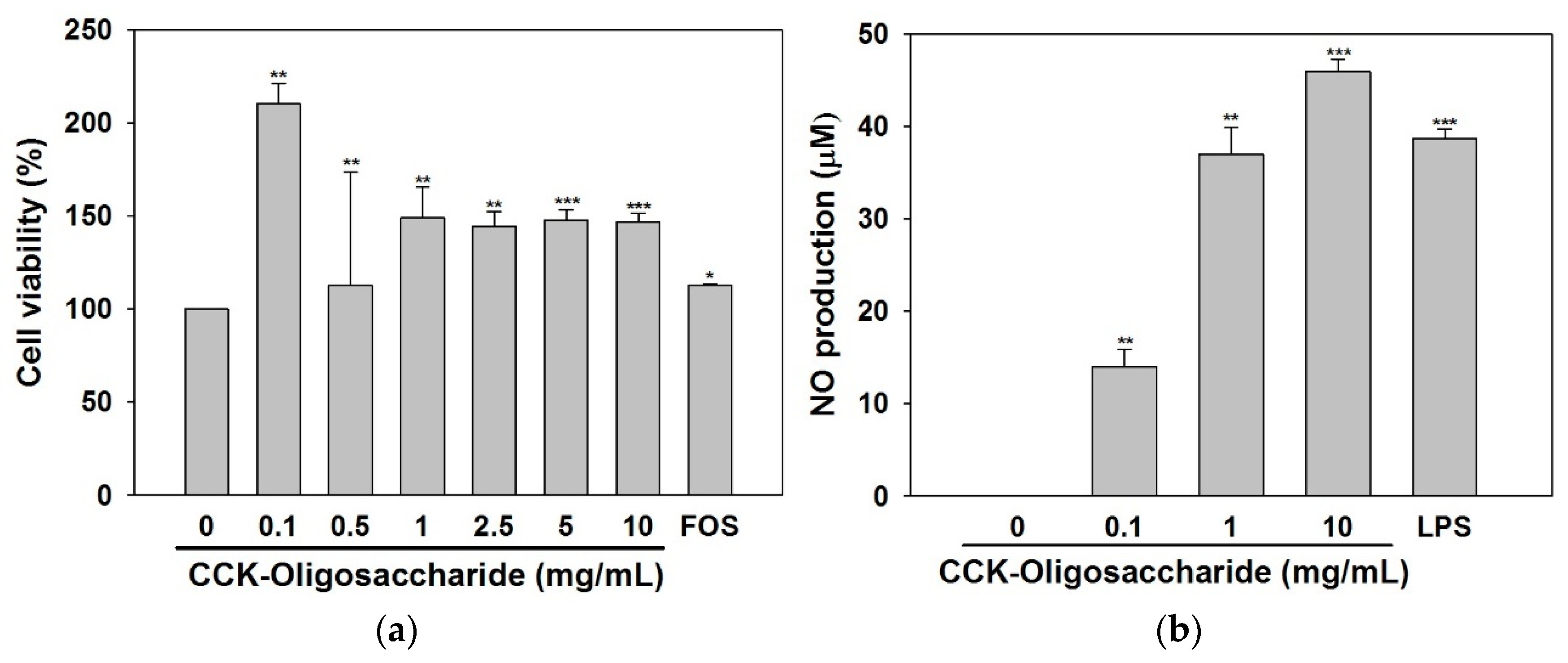
2.2. Effect of Oligosaccharides on RAW264.7 Cell Viability and NO Secretion
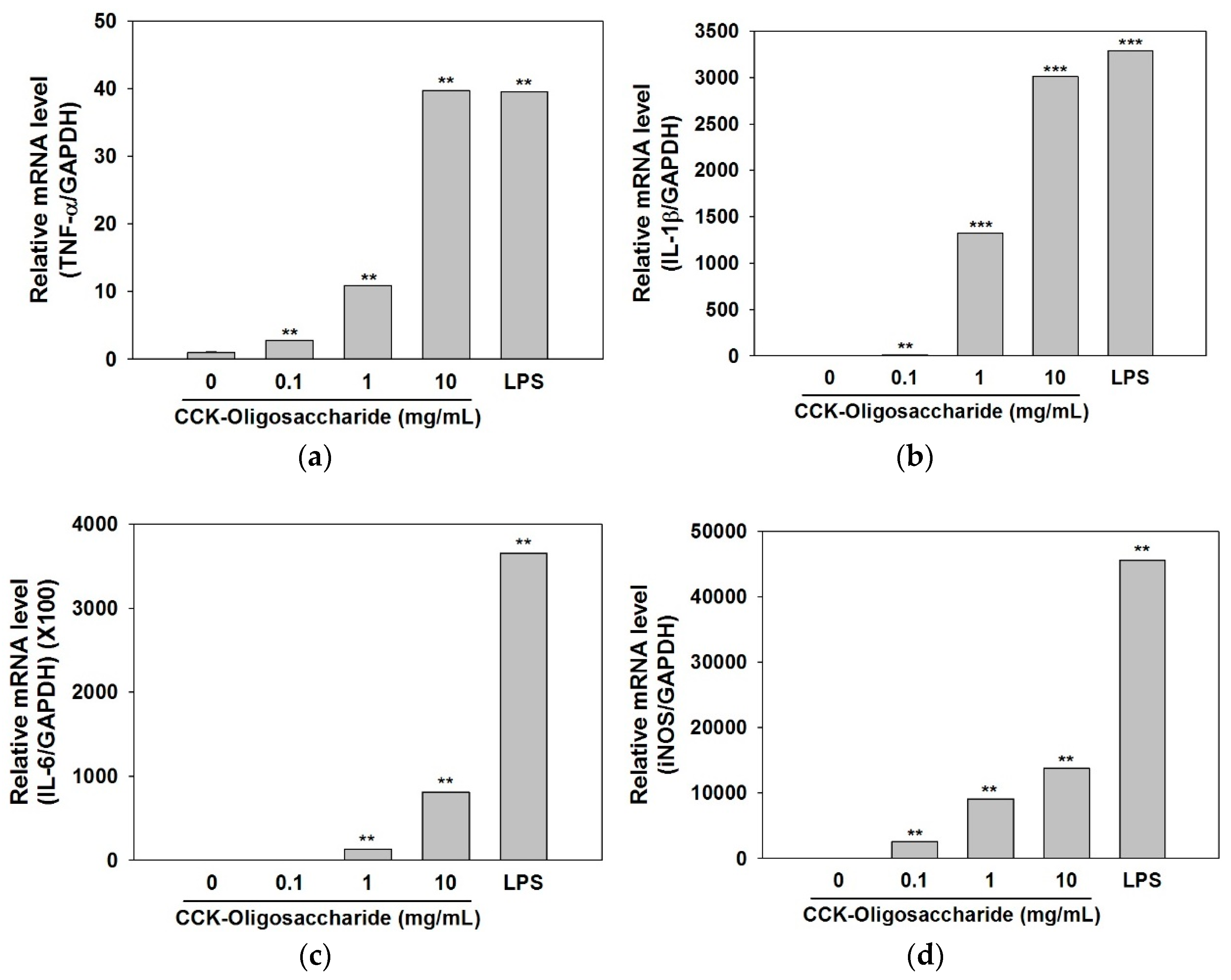
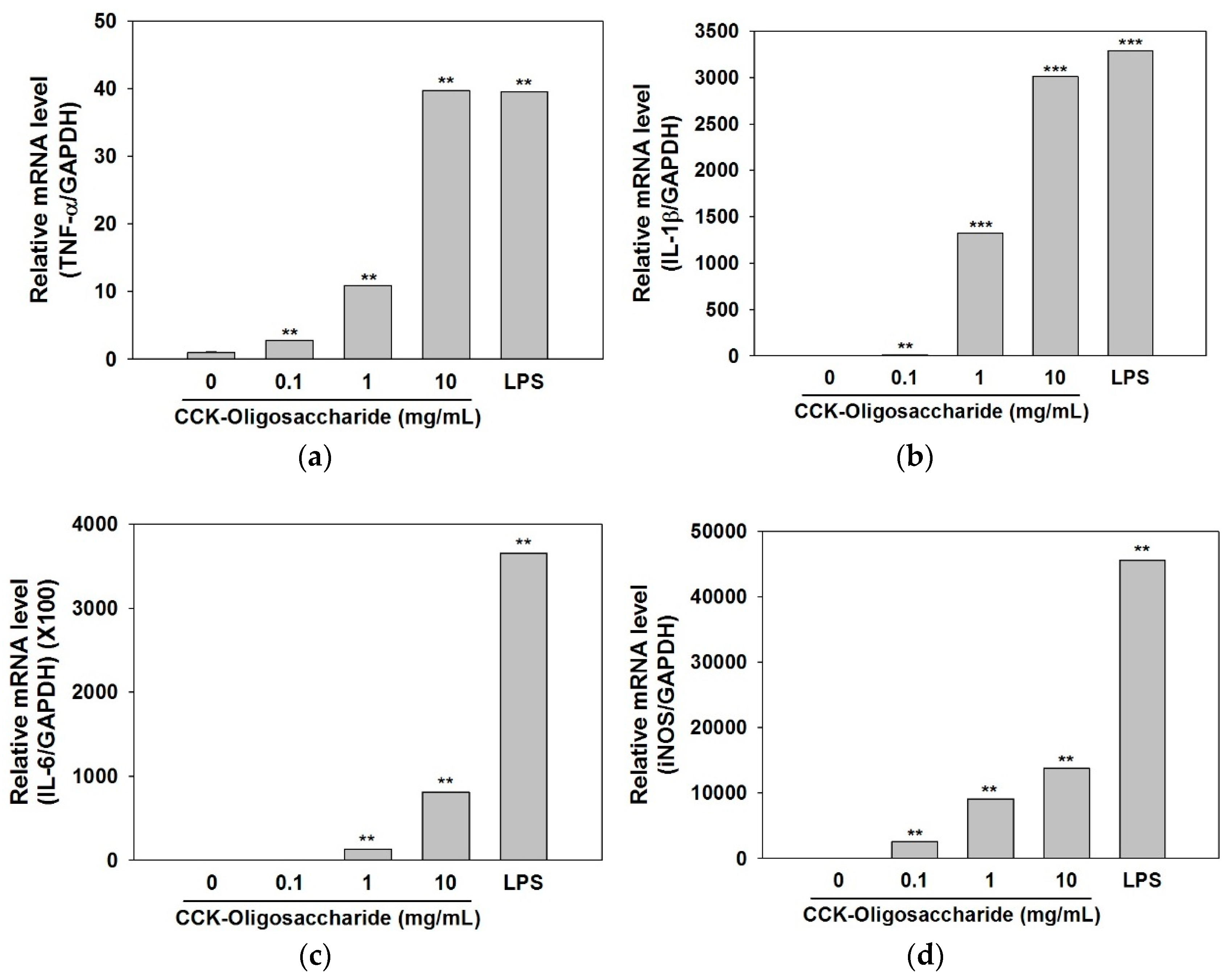
2.3. Effect of Oligosaccharides on Cytokine and iNOS Secretion
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. Response Surface Methodology Using Central Composite Design
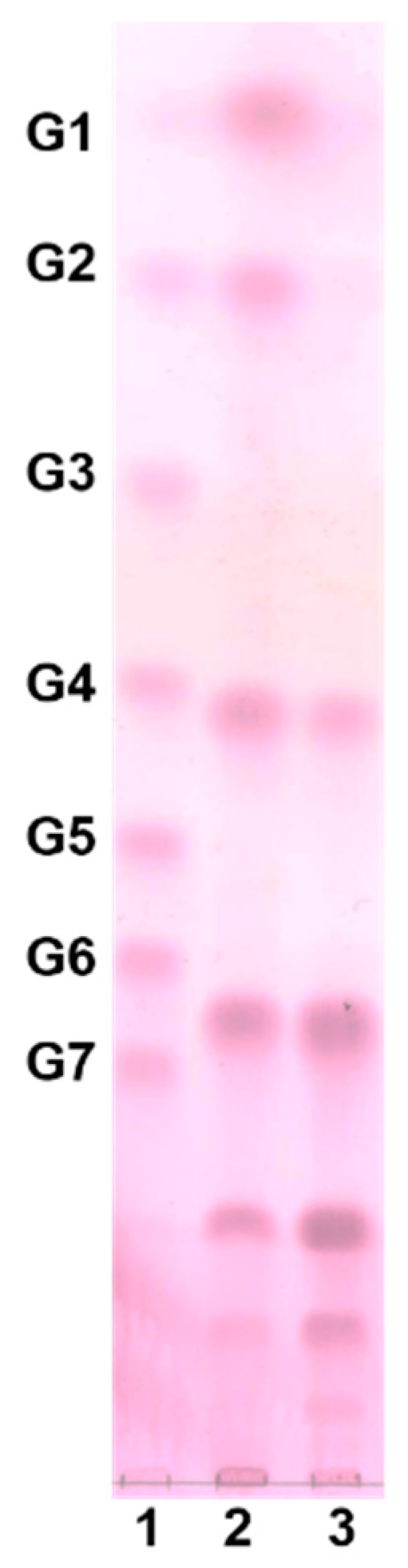
3.3. Analysis of Oligosaccharides by HPAEC-PAD
3.4. Purification of Oligosaccharides
3.5. Cell-Based Immunostimulatory Effect Test
3.5.1. Cell Culture
3.5.2. Cell Viability
3.5.3. NO Assay
3.5.4. Expression of Cytokines
3.5.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johansson, S.; Diehl, B.; Christakopoulos, P.; Austin, S.; Vafiadi, C. Oligosaccharide synthesis in fruit juice concentrates using a glucansucrase from Lactobacillus reuteri 180. Food Bioprod. Process. 2016, 98, 201–209. [Google Scholar] [CrossRef]
- Song, L.; Miao, M.; Jiang, B.; Xu, T.; Cui, S.W.; Zhang, T. Leuconostoc citreum SK24.002 glucansucrase: Biochemical characterisation and de novo synthesis of α-glucan. Int. J. Biol. Macromol. 2016, 91, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Baruah, R.; Goyal, A. Hyper glucansucrase, glucan and oligosaccharide producing novel Weissella cibaria RBA12 isolated from Pummelo (Citrus maxima). Ann. Microbiol. 2015, 65, 2301–2310. [Google Scholar] [CrossRef]
- Salim, А.S.; Bivolarski, V.P.; Vasileva, T.A.; Iliev, I.N. Enzymatic synthesis of fructo-oligosaccharides by recombinant levansucrase from Leuconostoc mesenteroides Lm17. Bulg. Chem. Commun. 2017, 49, 259–264. [Google Scholar]
- Argüello Morales, M.A.; Remaud-Simeon, M.; Willemot, R.M.; Vignon, M.R.; Monsan, P. Novel oligosaccharides synthesized from sucrose donor and cellobiose acceptor by alternansucrase. Carbohydr. Res. 2001, 331, 403–411. [Google Scholar] [CrossRef]
- Purama, R.K.; Goyal, A. Screening and optimization of nutritional factors for higher dextransucrase production by Leuconostoc mesenteroides NRRL B-640 using statistical approach. Bioresour. Technol. 2008, 99, 7108–7114. [Google Scholar] [CrossRef] [PubMed]
- Kanimozhi, J.; Moorthy, I.G.; Sivashankar, R.; Sivasubramanian, V. Optimization of dextran production by Weissella cibaria NITCSK4 using Response Surface Methodology-Genetic Algorithm based technology. Carbohydr. Polym. 2017, 174, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Daoudi, L.; Azaola, A. Optimization of galacto-oligosaccharide production by Bifidobacterium infantis RW-8120 using response surface methodology. J. Ind. Microbiol. Biotechnol. 2002, 29, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Côté, G.; Robyt, J. Acceptor reactions of alternansucrase from Leuconostoc mesenteroides NRRL B-1355. Carbohydr. Res. 1982, 111, 127–142. [Google Scholar] [CrossRef]
- Robyt, J.F.; Eklund, S.H. Relative, quantitative effects of acceptors in the reaction of Leuconostoc mesenteroides B-512F dextransucrase. Carbohydr. Res. 1983, 121, 279–286. [Google Scholar] [CrossRef]
- Lee, S.; Park, Y.-S. Oligosaccharide production by Leuconostoc lactis CCK940 which has glucansucrase activity. Food Eng. Prog. 2017, 21, 383–390. [Google Scholar] [CrossRef]
- Leardi, R. Experimental design in chemistry: A tutorial. Anal. Chim. Acta 2009, 652, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Babbar, N.; Dejonghe, W.; Sforza, S.; Elst, K. Enzymatic pectic oligosaccharides (POS) production from sugar beet pulp using response surface methodology. J. Food Sci. Technol. 2017, 54, 3707–3715. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Chen, Y.K.; Chang, H.C.; Lin, S.Y. Immunomodulatory effects of xylooligosaccharides. Food Sci. Technol. Res. 2012, 18, 195–199. [Google Scholar] [CrossRef]
- Xu, X.; Wu, X.; Wang, Q.; Cai, N.; Zhang, H.; Jiang, Z.; Wan, M.; Oda, T. Immunomodulatory effects of alginate oligosaccharides on murine macrophage RAW264.7 cells and their structure-activity relationships. J. Agric. Food Chem. 2014, 62, 3168–3176. [Google Scholar] [CrossRef] [PubMed]
- Majumder, A.; Goyal, A. Enhanced production of exocellular glucansucrase from Leuconostoc dextranicum NRRL B-1146 using response surface method. Bioresour. Technol. 2008, 99, 3685–3691. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jang, J.K.; Park, Y.S. Optimization of oligosaccharide production using response surface methodology and its immunostimulating effect on RAW264.7 macrophage cell. In Proceedings of the 2018 KoSFoST International Symposium and Annual Meeting, Busan, Korea, 27–29 June 2018. [Google Scholar]
- Cavaillon, J. Dossier cytokines and macrophages. Biomed. Pharmacother. 1994, 48, 445–453. [Google Scholar] [CrossRef]
- Cheong, K.L.; Meng, L.Z.; Chen, X.Q.; Wang, L.Y.; Wu, D.T.; Zhao, J.; Li, S.P. Structural elucidation, chain conformation and immuno-modulatory activity of glucogalactomannan from cultured Cordyceps sinensis fungus UM01. J. Funct. Foods 2016, 25, 174–185. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.A.; Liao, H.; Chesney, J.; Fingerle-Rowson, G.; Baugh, J.; David, J.; Bucala, R. Macrophage migration inhibitory factor (MIF) sustains macrophage proinflammatory function by inhibiting p53: Regulatory role in the innate immune response. Proc. Natl. Acad. Sci. USA 2002, 99, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Maresz, K.; Lee, P.-S.; Wu, J.C.; Ho, C.-T.; Popko, J.; Mehta, D.S.; Stohs, S.J.; Badmaev, V. Inhibition of TNF-α, IL-1α, and IL-1β by pretreatment of human monocyte-derived macrophages with menaquinone-7 and cell activation with TLR agonists in vitro. J. Med. Food 2016, 19, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.S.; Kim, W.J.; Kim, S.M.; Park, J.K.; Lee, S.M.; Kim, S.O.; Synytsya, A.; Park, Y. Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens. Int. Immunopharmacol. 2010, 10, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Yu, A.R.; Choi, I.W.; Hong, H.D.; Lee, K.W.; Choi, H.D. Immunostimulatory effects and characterization of a glycoprotein fraction from rice bran. Int. Immunopharmacol. 2013, 17, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.; Zum Büschenfelde, K.H.M.; Rose-John, S. The function of the soluble IL-6 receptor in vivo. Immunol. Lett. 1996, 54, 177–184. [Google Scholar] [CrossRef]
- Palmer, R.M.J.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Chou, C.J.; Chen, C.F. Camptothecin suppresses nitric oxide biosynthesis in RAW264.7 macrophages. Life Sci. 2001, 69, 625–635. [Google Scholar] [CrossRef]
- Seo, W.G.; Pae, H.O.; Oh, G.S.; Chai, K.Y.; Yun, Y.G.; Kwon, T.O.; Chung, H.T. Inhibitory effect of ethyl acetate fraction from Cudrania tricuspidata on the expression of nitric oxide synthase gene in RAW264.7 macrophages stimulated with interferon-γ and lipopolysaccharide. Gen. Pharmacol. Vasc. Syst. 2001, 35, 21–28. [Google Scholar] [CrossRef]
- Jeon, C.B.; Kim, Y.H.; Batsuren, D.; Tunsag, J.; Nho, C.W.; Pan, C.-H.; Lee, J.K. Immune-stimulating effects of Polygonum aviculare L. extract on macrophages. Yakhak Hoeji 2013, 57, 394–399. [Google Scholar]
Sample Availability: Not available. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Coded Variable Levels | Oligosaccharide Production (Relative Peak Area) | ||
|---|---|---|---|---|
| Temperature, °C (X1) | Sucrose, % (X2) | Maltose, % (X3) | ||
| 1 | 27.5 (−1) | 5.0 (−1) | 10.0 (+1) | 212.9 ± 13.1 * |
| 2 | 32.5 (+1) | 5.0 (−1) | 5.0 (−1) | 145.1 ± 20.5 ** |
| 3 | 30 (0) | 12.5 (+2) | 7.5 (0) | 312.6 ± 19.4 * |
| 4 | 27.5 (−1) | 10.0 (+1) | 10.0 (+1) | 248.9 ± 1.1 ** |
| 5 | 35 (+2) | 7.5 (0) | 7.5 (0) | 97.8 ± 12.8 |
| 6 | 30 (0) | 7.5 (0) | 7.5 (0) | 270.3 ± 1.9 *** |
| 7 | 30 (0) | 2.5 (−2) | 7.5 (0) | 139.0 ± 10.7 ** |
| 8 | 30 (0) | 7.5 (0) | 7.5 (0) | 248.1 ± 26.7 ** |
| 9 | 32.5 (+1) | 5.0 (−1) | 10.0 (+1) | 186.8 ± 14.8 ** |
| 10 | 25 (−2) | 7.5 (0) | 7.5 (0) | 227.0 ± 13.5 * |
| 11 | 27.5 (−1) | 10.0 (+1) | 5.0 (−1) | 234.1 ± 15.7 ** |
| 12 | 30 (0) | 7.5 (0) | 7.5 (0) | 244.7 ± 19.5 ** |
| 13 | 30 (0) | 7.5 (0) | 2.5 (−2) | 231.1 ± 10.9 * |
| 14 | 32.5 (+1) | 10.0 (+1) | 10.0 (+1) | 304.1 ± 18.6 * |
| 15 | 30 (0) | 7.5 (0) | 7.5 (0) | 259.6 ± 23.0 ** |
| 16 | 30 (0) | 7.5 (0) | 7.5 (0) | 233.5 ± 17.0 * |
| 17 | 30 (0) | 7.5 (0) | 7.5 (0) | 245.0 ± 23.4 ** |
| 18 | 30 (0) | 7.5 (0) | 12.5 (+2) | 252.3 ± 12.9 ** |
| 19 | 27.5 (−1) | 5.0 (−1) | 5.0 (−1) | 183.8 ± 7.6 * |
| 20 | 32.5 (+1) | 10.0 (+1) | 5.0 (−1) | 270.3 ± 12.1 * |
| Variables | DF | Adj SS | Adj MS | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 9 | 151,743 | 16,860.3 | 10.78 | 0.000 |
| Linear | 3 | 61,162 | 20,387.5 | 13.03 | 0.000 |
| Quadratic | 3 | 77,970 | 25,990.1 | 16.61 | 0.000 |
| Cross-product | 3 | 12,610 | 4203.4 | 2.69 | 0.056 |
| X1 | 1 | 7750 | 7750.0 | 4.95 | 0.031 |
| X2 | 1 | 50,631 | 50,631.1 | 32.37 | 0.000 |
| X3 | 1 | 2781 | 2781.3 | 1.78 | 0.188 |
| X12 | 1 | 75,158 | 75,157.8 | 48.04 | 0.000 |
| X22 | 1 | 10,678 | 10,678.1 | 6.83 | 0.012 |
| X32 | 1 | 1653 | 1653.5 | 1.06 | 0.309 |
| X1X2 | 1 | 7159 | 7159.0 | 4.58 | 0.037 |
| X1X3 | 1 | 5436 | 5436.4 | 3.48 | 0.068 |
| X2X3 | 1 | 15 | 15.0 | 0.01 | 0.922 |
| Residual | 50 | 78,217 | 1564.3 | - | - |
| Lack of fit | 5 | 13,947 | 2789.5 | 1.95 | 0.104 |
| Pure error | 45 | 64,270 | 1428.2 | - | - |
| Cor Total | 59 | 229,960 |
| Gene | Sequence | |
|---|---|---|
| GAPDH | Forward | 5′―ATC CCA TCA CCA TCT TCC AG―3′ |
| Reverse | 5′―CCT GCT TCA CCA CCT TCT TG―3′ | |
| TNF-α | Forward | 5′―ATG AGC ACA GAA AGC ATG ATC CG―3′ |
| Reverse | 5′―CCA AAG TAG ACC TGC CCG GAC TC―3′ | |
| IL-1β | Forward | 5′―ATG GCA ACT GTT CCT GAA CTC AAC T―3′ |
| Reverse | 5′―CAG GAC AGG TAT AGA TTC TTT CCT T―3′ | |
| IL-6 | Forward | 5′―CAA GAG ACT TCC ATC CAG TTG C―3′ |
| Reverse | 5′―TTG CCG AGT TCT CAA AGT GAC―3′ | |
| iNOS | Forward | 5′―AAT GGC AAC ATC AGG TCG GCC ATC ACT―3′ |
| Reverse | 5′―GCT GTG TGT CAC AGA AGT CTC GAA CTC―3′ | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Park, G.-G.; Jang, J.-K.; Park, Y.-S. Optimization of Oligosaccharide Production from Leuconostoc lactis Using a Response Surface Methodology and the Immunostimulating Effects of These Oligosaccharides on Macrophage Cells. Molecules 2018, 23, 2118. https://doi.org/10.3390/molecules23092118
Lee S, Park G-G, Jang J-K, Park Y-S. Optimization of Oligosaccharide Production from Leuconostoc lactis Using a Response Surface Methodology and the Immunostimulating Effects of These Oligosaccharides on Macrophage Cells. Molecules. 2018; 23(9):2118. https://doi.org/10.3390/molecules23092118
Chicago/Turabian StyleLee, Sulhee, Gwi-Gun Park, Jae-Kweon Jang, and Young-Seo Park. 2018. "Optimization of Oligosaccharide Production from Leuconostoc lactis Using a Response Surface Methodology and the Immunostimulating Effects of These Oligosaccharides on Macrophage Cells" Molecules 23, no. 9: 2118. https://doi.org/10.3390/molecules23092118
APA StyleLee, S., Park, G.-G., Jang, J.-K., & Park, Y.-S. (2018). Optimization of Oligosaccharide Production from Leuconostoc lactis Using a Response Surface Methodology and the Immunostimulating Effects of These Oligosaccharides on Macrophage Cells. Molecules, 23(9), 2118. https://doi.org/10.3390/molecules23092118