Natural Cyclic Peptides as an Attractive Modality for Therapeutics: A Mini Review


Abstract
1. Introduction
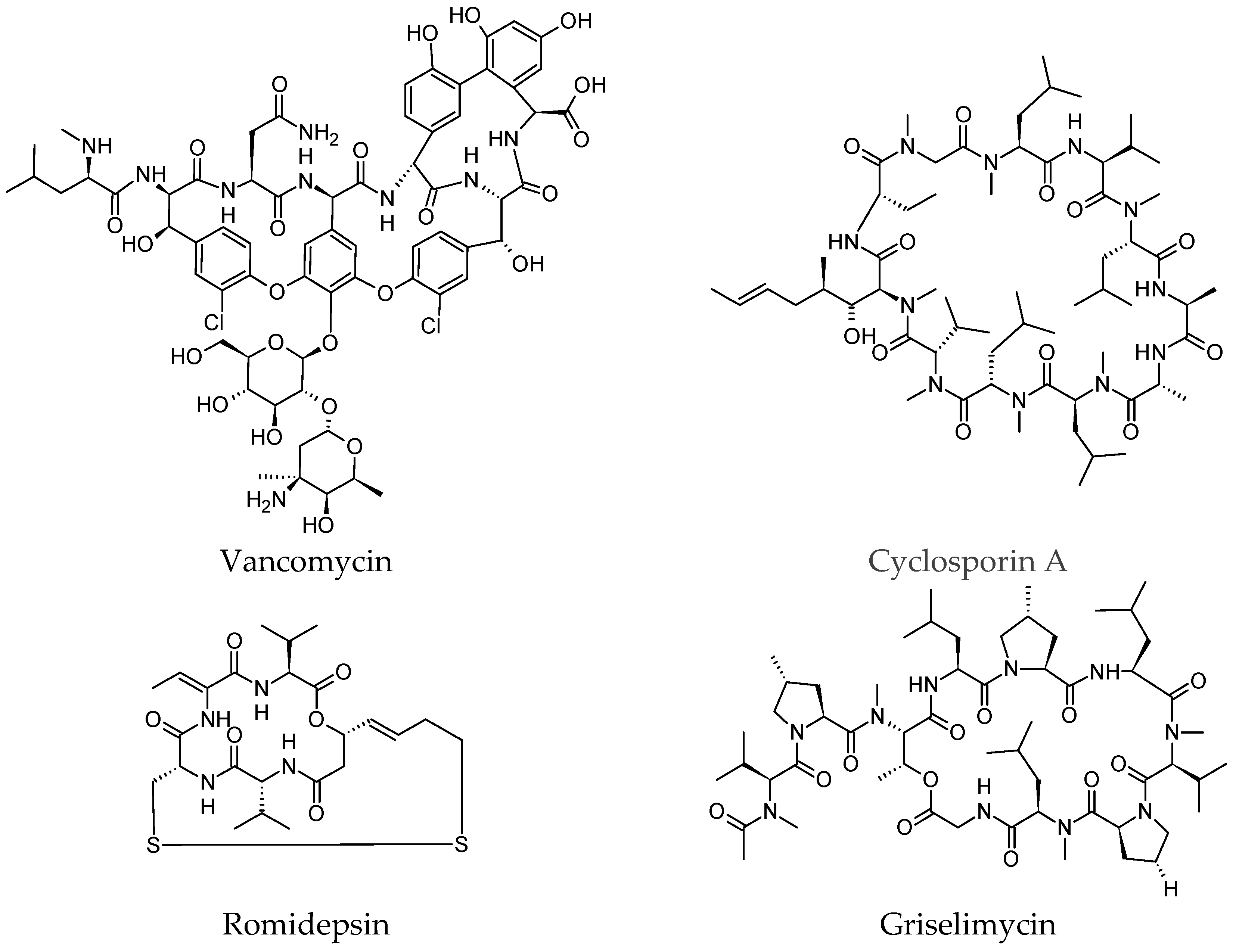
2. Examples of Previously Isolated Cyclic Peptides Used as Therapeutic Agents
3. Natural Cyclic Peptides and Analogues in Clinical Trials
4. Recently Reported Naturally Occurring Bioactive Cyclopeptides Covering 2014–2018
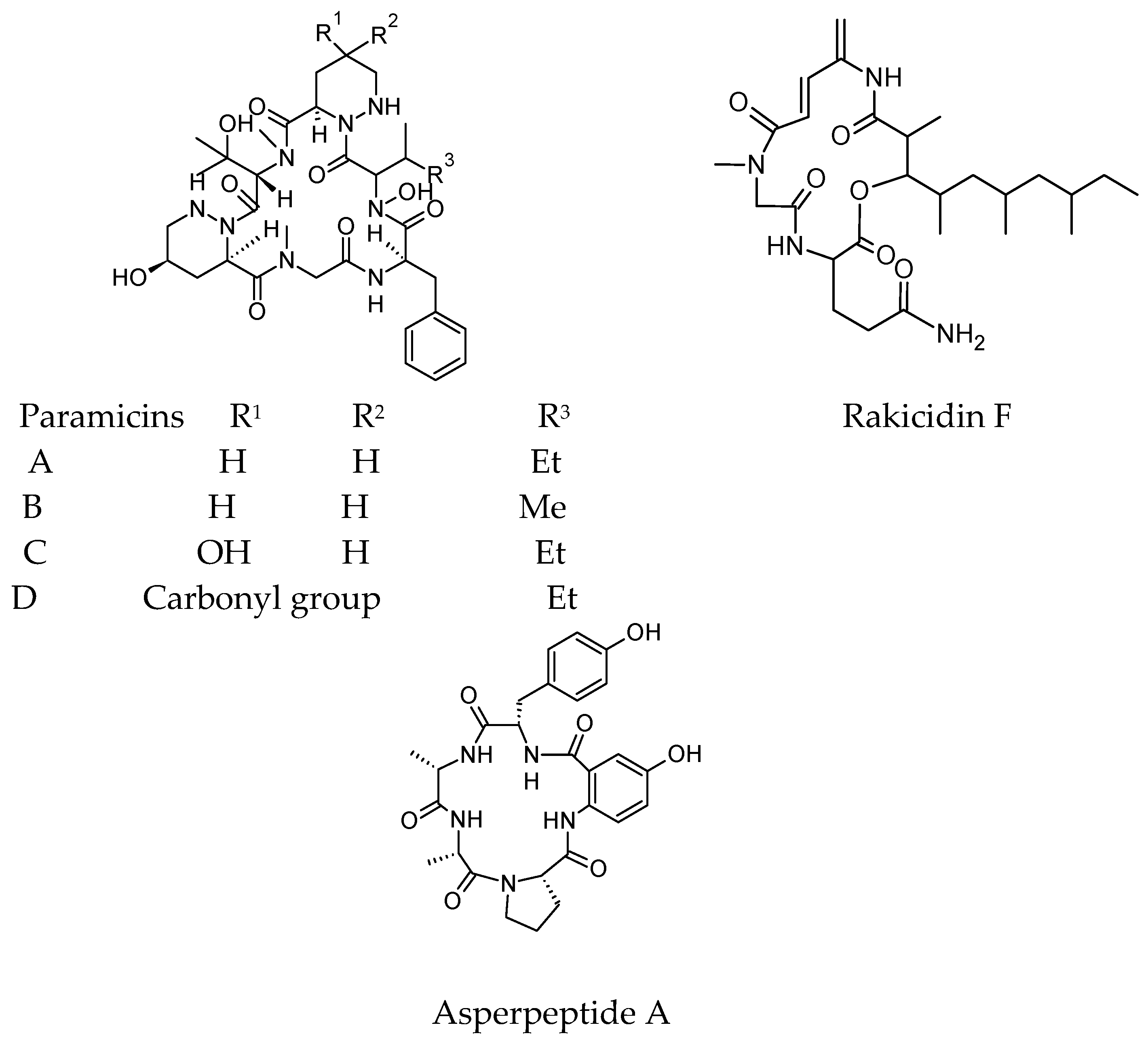
4.1. Antibacterial Cyclic Peptides
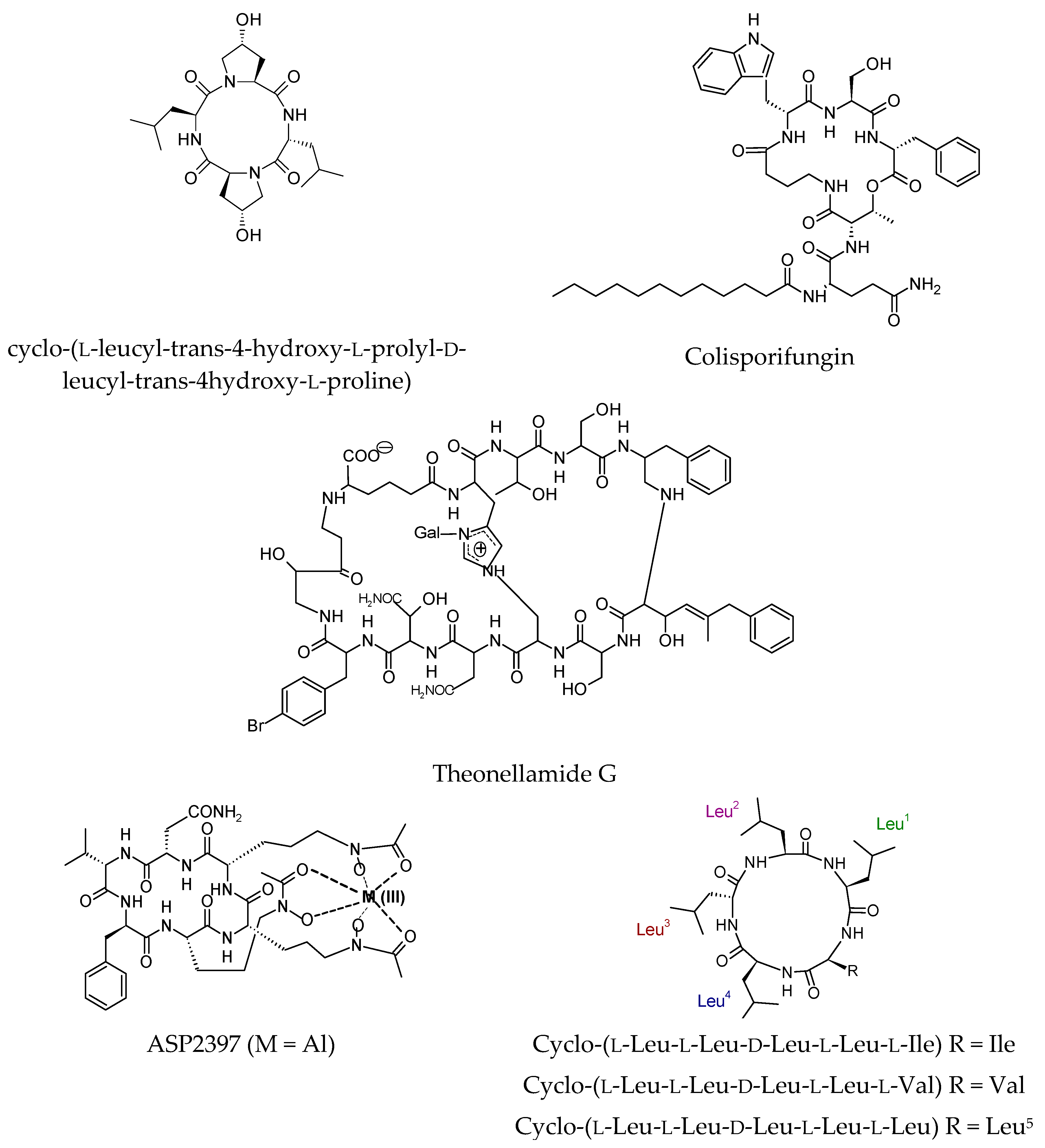
4.2. Antifungal Cyclic Peptides
4.3. Anticancer Cyclic Peptides
4.4. Recently Antiviral Isolated Cyclic Peptides
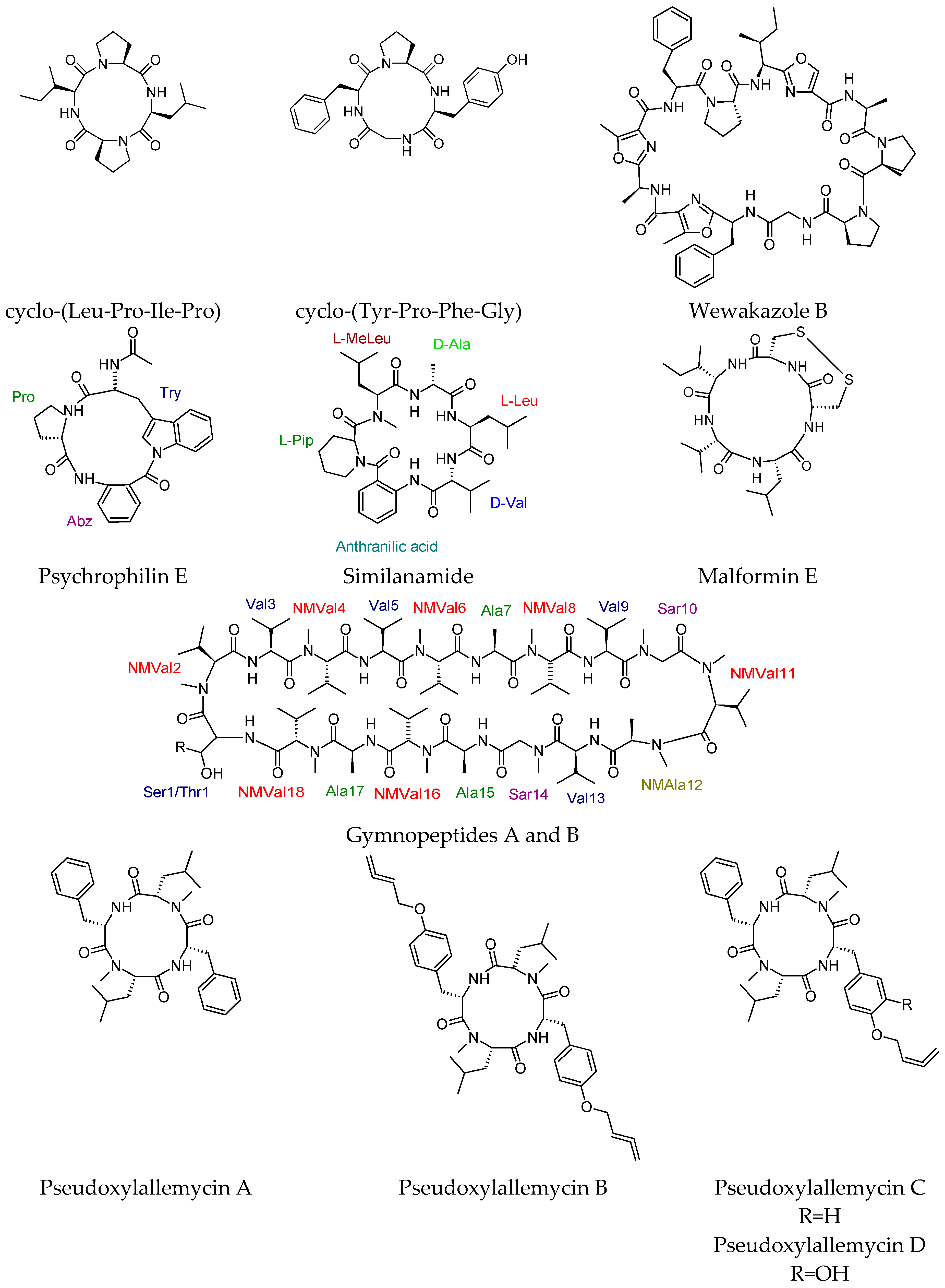
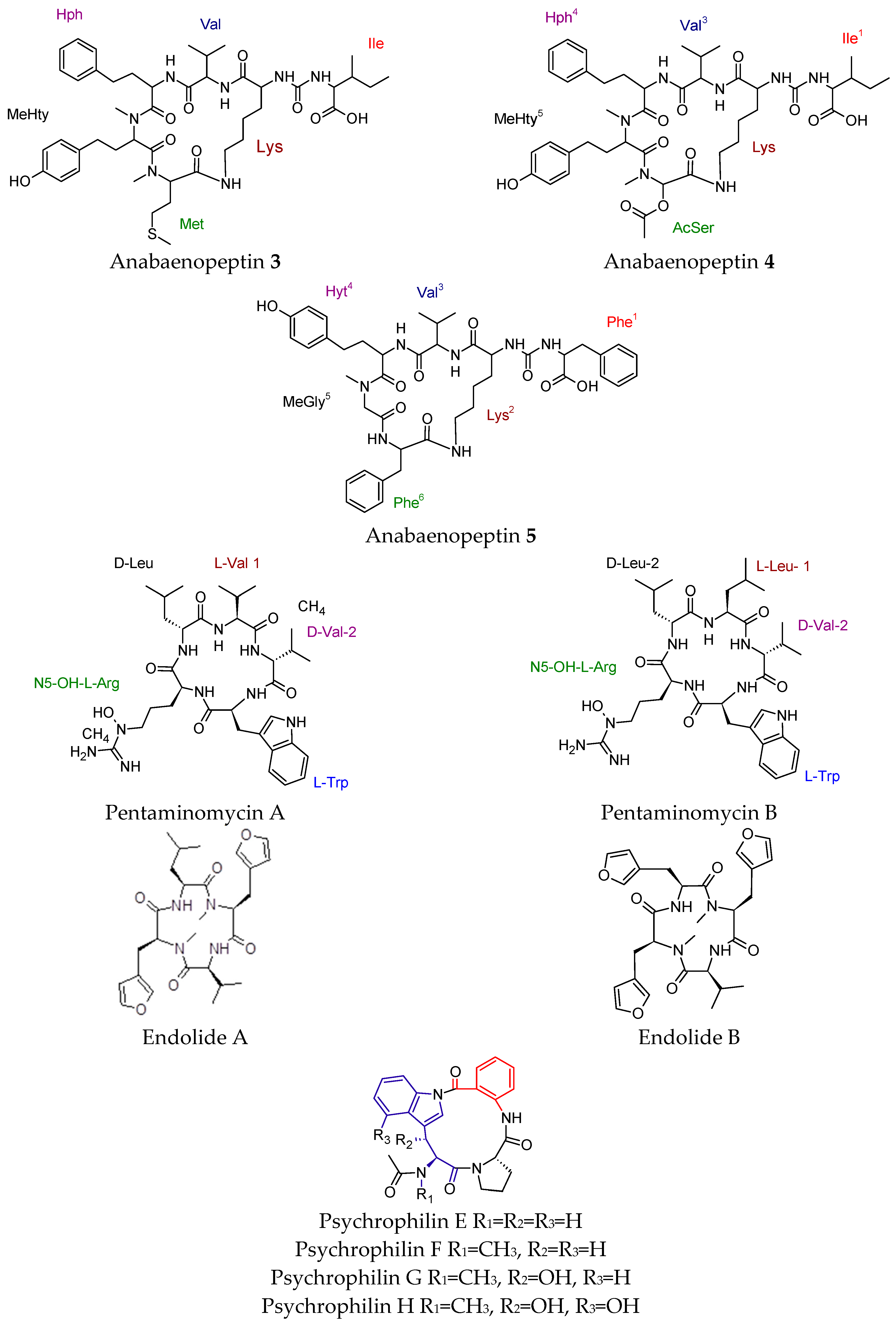
4.5. Cyclic Peptides with Diverse Biological Activities
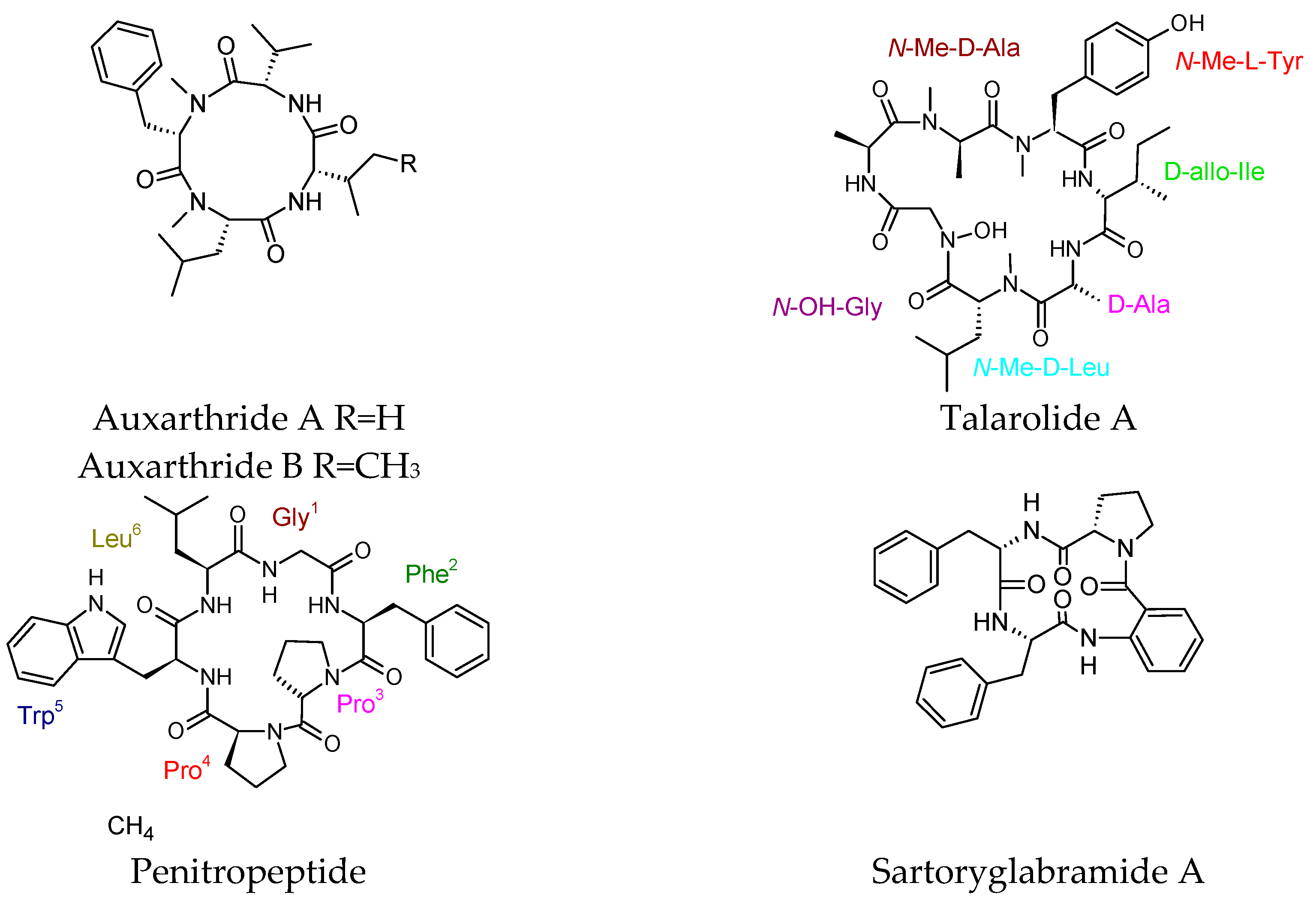
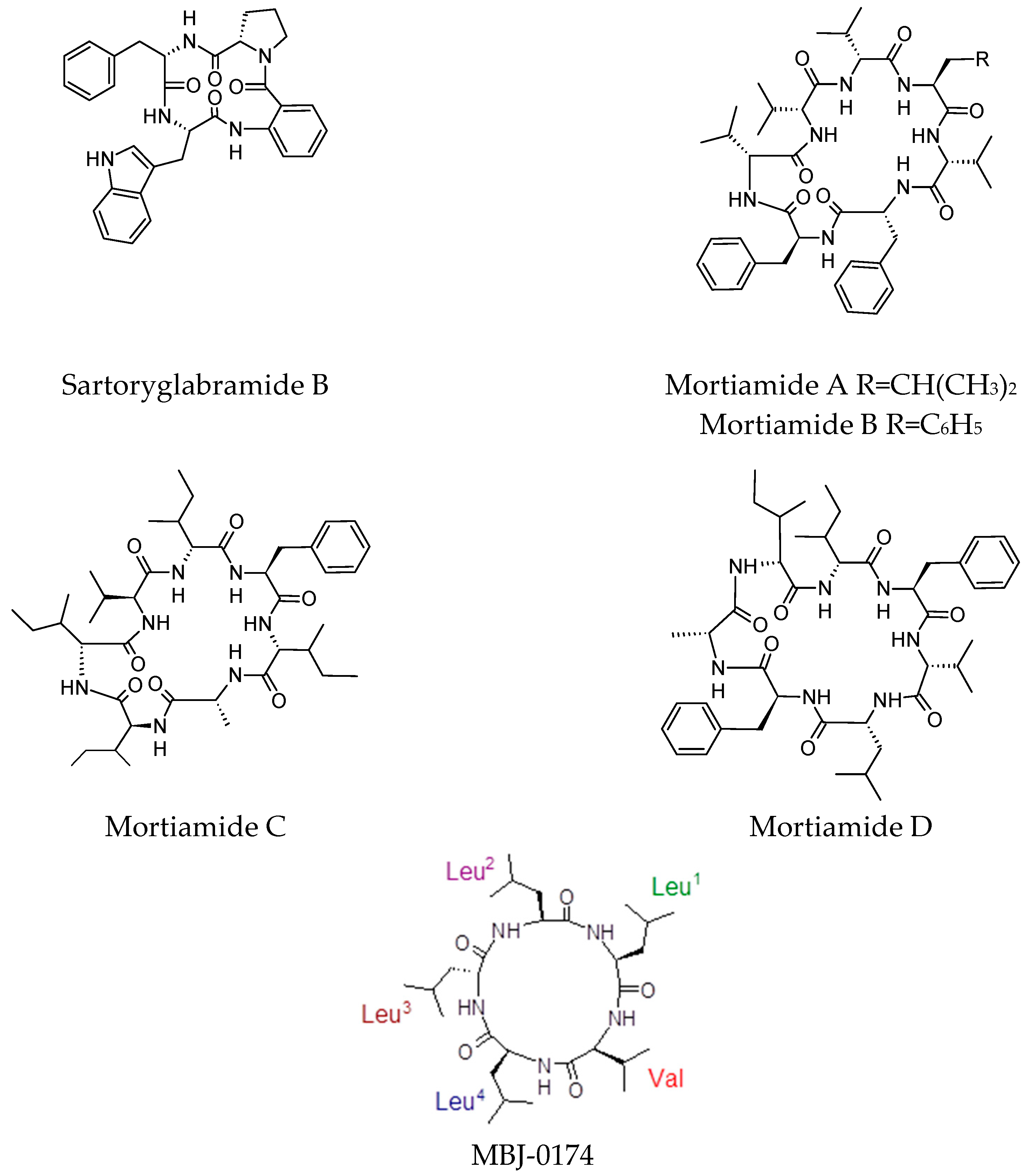
4.6. Cyclic Peptides with No Reported Biological Activity
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Waldhoer, M.; Bartlett, S.E.; Whistler, J.L. Opioid receptors. Annu. Rev. Biochem. 2004, 73, 953–990. [Google Scholar] [CrossRef] [PubMed]
- Neefjes, J.; Ovaa, H. A peptide’s perspective on antigen presentation to the immune system. Nat. Chem. Biol. 2013, 9, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Pontiroli, A.E. Peptide hormones: Review of current and emerging uses by nasal delivery. Adv. Drug Deliv. Rev. 1998, 29, 81–87. [Google Scholar] [CrossRef]
- Burgen, A.S.V.; Roberts, G.C.K.; Feeney, J. Binding of flexible ligands to macromolecules. Nature 1975, 253, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Ehrmann, M.; Clausen, T. Proteolysis as a regulatory mechanism. Annu. Rev. Genet. 2004, 38, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The future of peptide-based drugs. Chem. Biol. Drug 2013, 81, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Driggers, E.M.; Hale, S.P.; Lee, J.; Terrett, N.K. The exploration of macrocycles for drug discovery—An underexploited structural class. Nat. Rev. Drug Discov. 2008, 7, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, A.; Deyle, K.; Heinis, C. Cyclic peptide therapeutics: Past, present and future. Curr. Opin. Chem. Biol. 2017, 38, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Choi, M.C.; Seo, C.H.; Park, Y. Therapeutic properties and biological benefits of marine-derived anticancer peptides. Int. J. Mol. Sci. 2018, 19, 919. [Google Scholar] [CrossRef] [PubMed]
- Edman, P. Chemistry of amino acids and peptides. Annu. Rev. Biochem. 1959, 28, 69–96. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.A.; Bourne, G.T.; Smythe, M.L. Exploring privileged structures: The combinatorial synthesis of cyclic peptides. J. Comput. Aided Mol. Des. 2002, 16, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.H. Cyclic peptides as therapeutic agents and biochemical tools. Biomol. Ther. (Seol.) 2012, 20, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Fan, J.; Liu, J.; He, L.; He, K. Applications of cyclic peptide nanotubes. Prog. Chem. 2010, 22, 648–653. [Google Scholar]
- Sun, L.; Fan, Z.; Wang, Y.; Huang, Y.; Schmidt, M.; Zhang, M. Tunable synthesis of self-assembled cyclic peptide nanotubes and nanoparticles. Soft Matter 2015, 11, 3822–3832. [Google Scholar] [CrossRef] [PubMed]
- Tiangtrong, P.; Thamwattana, N.; Baowan, D. Modelling water molecules inside cyclic peptide nanotubes. Appl. Nanosci. 2016, 6, 345–357. [Google Scholar] [CrossRef]
- Tan, N.-H.; Zhou, J. Plant cyclopeptides. Chem. Rev. 2006, 106, 840–895. [Google Scholar] [CrossRef] [PubMed]
- Taevernier, L.; Wynendaele, E.; De Vreese, L.; Burvenich, C.; De Spiegeleer, B. The mycotoxindefinition reconsidered towards fungal cyclic depsipeptides. J. Environ. Sci. Health Part C 2016, 34, 114–135. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A. Medicinal significance of naturally occurring cyclotetrapeptides. J. Nat. Med. 2016, 70, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, X.; Kim, S.J.; Zhang, W. Antimycin-type depsipeptides: Discovery, biosynthesis, chemical synthesis, and bioactivities. Nat. Prod. Rep. 2016, 33, 1146–1165. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A. Three new cyclotetrapeptides isolated from Streptomyces sp. 447. Nat. Prod. Res. 2017, 31, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Matasyoh, J.C. Endophytes as producers of peptides: An overview about the recently discovered peptides from endophytic microbes. Nat. Prod. Bioprospect. 2014, 4, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Andavan, G.S.B.; Lemmens-Gruber, R. Cyclodepsipeptides from marine sponges: Natural agents for drug research. Mar. Drugs 2010, 8, 810–834. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, X.; Wang, B.-G. Secondary metabolites from the marine algal-derived endophtic fungi: Chemical diversity and biological activity. Planta Med. 2016, 82, 832–842. [Google Scholar] [PubMed]
- Falanga, A.; Nigro, E.; De biasi, M.G.; Daniele, A.; Morelli, G.; Galdiero, S.; Scudiero, O. Cyclic peptides as novel therapeutic microbicides engineering of human defensing mimetics. Molecules 2017, 22, 1217. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine Peptides: Bioactivities and Applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- McGivern, J.G. Ziconotide: A review of its pharmacology and use in the treatment of pain. Neuropsych. Dis. Treat. 2007, 3, 69–85. [Google Scholar] [CrossRef]
- Webster, L.R. The relationship between the mechanisms of action and safety profiles of intrathecal morphine and ziconotide: A review of the literature. Pain Med. 2015, 16, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Volk, K.J.; Lee, M.S.; Pucci, M.; Handwerger, S. Binding studies of vancomycin to the cytoplasmic peptidoglycan precursors by affinity capillary electrophoresis. J. Anal. Chem. 1994, 66, 2412–2416. [Google Scholar] [CrossRef]
- Li, B.; Brown, K.V.; Wenke, J.C.; Guelcher, S.A. Sustained release of vancomycin from polyurethane scaffolds inhibits infection of bone wounds in a rat femoral segmental defect model. J. Control. Release 2010, 145, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Holland, T.L.; Arnold, C.; Fowler, V.G. Clinical management of Staphylococcus aureus Bacteremia: A review. JAMA 2014, 312, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Levine, D.P. Vancomycin: A History. Clin. Infect. Dis. 2006, 42, S5–S12. [Google Scholar] [CrossRef] [PubMed]
- Faron, M.L.; Ledeboer, N.A.; Buchan, B.W. Resistance mechanisms, epidemiology, and approaches to screening for Vancomycin-Resistant Enterococcus in the health care setting. J. Clin. Microbiol. 2016, 54, 2436–2447. [Google Scholar] [CrossRef] [PubMed]
- Sieradzki, K.; Roberts, R.B.; Haber, S.W.; Tomasz, A. The development of vancomycin resistance in a patient with Methicillin-Resistant Staphylococcus aureus Infection. N. Engl. J. Med. 1999, 340, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Yarlagadda, V.; Sarkar, P.; Samaddar, S.; Haldar, J. A vancomycin derivative with a pyrophosphate-binding group: A strategy to combat Vancomycin-Resistant Bacteria. Angew. Chem. 2016, 128, 7967–7971. [Google Scholar] [CrossRef]
- Rüegger, A.; Kuhn, M.; Lichti, H.; Loosli, H.R.; Huguenin, R.; Quiquerez, C.; Von Wartburg, A. Cyclosporin A, ein immunsuppressiv wirksamer Peptidmetabolit aus Trichoderma polysporum (LINK ex PERS.) Rifai. Helv. Chim. Acta 1976, 59, 1075–1092. [Google Scholar] [CrossRef] [PubMed]
- Campana, C.; Regazzi, M.B.; Buggia, I.; Molinaro, M. Clinically significant drug interactions with cyclosporin: An update. Clin. Pharmacokinet. 1996, 30, 141–179. [Google Scholar] [CrossRef] [PubMed]
- Li, K.W.; Wu, J.; Xing, W.; Simon, J.A. Total synthesis of the antitumor depsipeptide FR-901, 228. J. Am. Chem. Soc. 1996, 118, 7237–7238. [Google Scholar] [CrossRef]
- Barbarotta, L.; Hurley, K. Romidepsin for the treatment of peripheral T-Cell lymphoma. J. Adv. Pract. Oncol. 2015, 6, 22–36. [Google Scholar] [PubMed]
- Terlain, B.; Thomas, J.P. Structure of griselimycin, polypeptide antibiotic extracted Streptomyces cultures. I. identification of the products liberated by hydrolysis. Bull. Soc. Chim. Fr. 1971, 6, 2349–2356. [Google Scholar] [PubMed]
- Kling, A.; Lukat, P.; Almeida, D.V.; Bauer, A.; Fontaine, E.; Sordello, S.; Zaburannyi, N.; Herrmann, J.; Wenzel, S.C.; Konig, C.; et al. Targeting DnaN for tuberculosis therapy using novel griselimycins. Science 2015, 348, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Holzgrabe, U. New griselimycins for treatment of tuberculosis. Chem. Biol. 2015, 22, 981–982. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.L. The role of vancomycin in the treatment paradigm. Clin. Infect. Dis. 2006, 42, S51–S57. [Google Scholar] [CrossRef] [PubMed]
- Hammes, W.P.; Neuhaus, F.C. On the mechanism of action of Vancomycin: Inhibition of peptidoglycan synthesis in Gaffkya homari. Antimicrob. Agents Chemother. 1974, 6, 722–728. [Google Scholar] [CrossRef]
- Chinen, J.; Buckley, R.H. Transplantation immunology: Solid organ and bone marrow. J. Allergy Clin. Immunol. 2010, 125, S324–S335. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Koyasu, S. Mechanisms of action of cyclosporine. Immunopharmacology 2000, 47, 119–125. [Google Scholar] [CrossRef]
- Biasutto, L.; Azzolini, M.; Szabò, I.; Zoratti, M. The mitochondrial permeability transition pore in AD 2016: An update. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2515–2530. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, I.; Matsubara, H.; Hanari, N.; Mori, M.; Nishimori, T.; Yoneyama, Y.; Akutsu, Y.; Sakata, H.; Matsushita, K.; Sekiet, N.; et al. Histone deacetylase inhibitor FK.228 activates tumor suppressor Prdx1 with apoptosis induction in esophageal cancer cells. Clin. Cancer Res. 2005, 11, 7945–7952. [Google Scholar] [CrossRef] [PubMed]
- Suárez, Y.; González, L.; Cuadrado, A.; Berciano, M.; Lafarga, M.; Muñoz, A. Kahalalide F, a new marine-derived compound, induces oncosis in human prostate and breast cancer cells. Mol. Cancer Ther. 2003, 2, 863–872. [Google Scholar] [PubMed]
- Janmaat, M.L.; Rodriguez, J.A.; Jimeno, J.; Kruyt, F.A.E.; Giaccone, G. Kahalalide F induces necrosis-like cell death that involves depletion of ErbB3 and inhibition of Akt signaling. Mol. Pharmacol. 2005, 68, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Pardo, B.; Paz-Ares, L.; Tabernero, J.; Ciruelos, E.; García, M.; Salazar, R.; López, A.; Blanco, M.; Nieto, A.; Jimeno, J.; et al. Phase I clinical and pharmacokinetic study of kahalalide F administered weekly as a 1-hour infusion to patients with advanced solid tumors. Clin. Cancer Res. 2008, 14, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Martin-Algarra, S.; Espinosa, E.; Rubio, J.; Lopez Lopez, J.J.; Manzano, J.L.; Carrion, L.A.; Plazaola, A.; Tanovic, A.; Paz-Ares, L. Phase II study of weekly kahalalide F in patients with advanced malignant melanoma. Eur. J. Cancer 2009, 45, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Salazar, R.; Cortes-Funes, H.; Casado, E.; Pardo, B.; Lopez-Martin, A.; Cuadra, C.; Tabernero, J.; Coronado, C.; Garcia, M.; Soto Matos-Pita, A.; et al. Phase I study of weekly kahalalide F as prolonged infusion in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2013, 72, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Serova, M.; de Gramont, A.; Bieche, I.; Riveiro, M.E.; Galmarini, C.M.; Aracil, M.; Jimeno, J.; Faivre, S.; Raymond, E. Predictive factors of sensitivity to elisidepsin, a novel kahalalide F-derived marine compound. Mar. Drugs 2013, 11, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.; Viteri, S.; Morán, T.; Coronado, C.; Dios, J.L.; Miguel-Lillo, B.; Fernández-García, E.M.; Rosell, R. Phase I, dose-escalating study of elisidepsin (Irvalec®), a plasma membrane-disrupting marine antitumor agent, in combination with erlotinib in patients with advanced malignant solid tumors. Investig. New Drugs 2016, 34, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Molina-Guijarro, J.M.; García, C.; Macías, Á.; García-Fernández, L.F.; Moreno, C.; Reyes, F.; Martínez-Leal, J.F.; Fernández, R.; Martínez, V.; Valenzuela, C.; et al. Elisidepsin interacts directly with glycosylceramides in the plasma membrane of tumor cells to induce necrotic cell death. PLoS ONE 2015, 10, e0140782. [Google Scholar] [CrossRef] [PubMed]
- Salazar, R.; Jones, R.J.; Oaknin, A.; Crawford, D.; Cuadra, C.; Hopkins, C.; Gil, M.; Coronado, C.; Soto-Matos, A.; Cullell-Young, M.; et al. A phase I and pharmacokinetic study of elisidepsin (PM02734) in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2012, 70, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Geoerger, B.; Estlin, E.J.; Aerts, I.; Kearns, P.; Gibson, B.; Corradini, N.; Doz, F.; Lardelli, P.; Miguel, B.D.; Soto, A.; et al. A phase I and pharmacokinetic study of plitidepsin in children with advanced solid tumours: Innovative therapies for children with cancer (ITCC) study. Eur. J. Cancer 2012, 48, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.A.; Giroux, D.J.; Jaeckle, K.A.; Panella, T.J.; Dakhil, S.R.; Schold, S.C. Phase II study of didemnin B in central nervous system tumors: A southwest oncology group study. Investig. New Drugs 1998, 16, 331–332. [Google Scholar] [CrossRef]
- Dorr, F.A.; Kuhn, J.G.; Phillips, J.; von Hoff, D.D. Phase I clinical and pharmacokinetic investigation of didemnin B, a cyclic depsipeptide. Eur. J. Cancer Clin. Oncol. 1988, 24, 1699–1706. [Google Scholar] [CrossRef]
- Kucuk, O.; Young, M.L.; Habermann, T.M.; Wolf, B.C.; Jimeno, J.; Cassileth, P.A. Phase II trial of didemnin B in previously treated non-Hodgkin’s lymphoma: An eastern cooperative oncology group (ECOG) study. Am. J. Clin. Oncol. 2000, 23, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Mittelman, A.; Chun, H.G.; Puccio, C.; Coombe, N.; Lansen, T.; Ahmed, T. Phase II clinical trial of didemnin B in patients with recurrent or refractory anaplastic astrocytoma or glioblastoma multiforme (NSC 325319). Investig. New Drugs 1999, 17, 179–182. [Google Scholar] [CrossRef]
- Nuijen, B.; Bouma, M.; Manada, C.; Jimeno, J.M.; Schellens, J.H.; Bult, A.; Beijnen, J.H. Pharmaceutical development of anticancer agents derived from marine sources. Anticancer Drugs 2000, 11, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Adrio, J.; Cuevas, C.; Manzanares, I.; Joullie, M.M. Total synthesis and biological evaluation of tamandarin B analogues. J. Org. Chem. 2007, 72, 5129–5138. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Álvarez, S.; Pardal, E.; Sánchez-Nieto, D.; Navarro, M.; Caballero, M.D.; Mateos, M.V.; Martín, A. Plitidepsin: Design, development, and potential place in therapy. Drug Des. Dev. Ther. 2017, 11, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, R.; Hirsch, C.; Sesin, D.; Flor, J.; Chartrain, M.; Fromtling, R.; Harris, G.; Salvatore, M.; Liesch, J.; Yudin, K. Pharmaceuticals from cultured algae. J. Ind. Microbiol. 1990, 5, 113–123. [Google Scholar] [CrossRef]
- Smith, C.D.; Zhang, X.; Mooberry, S.L.; Patterson, G.M.; Moore, R.E. Cryptophycin: A new antimicrotubule agent active against drug-resistant cells. Cancer Res. 1994, 54, 3779–3784. [Google Scholar] [PubMed]
- Lu, K.; Dempsey, J.; Schultz, R.M.; Shih, C.; Teicher, B.A. Cryptophycin-induced hyperphosphorylation of Bcl-2, cell cycle arrest and growth inhibition in human H460 NSCLC cells. Cancer Chemother. Pharmacol. 2001, 47, 170–178. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, G.; del Campo, J.; Mellado, B.; Izquierdo, M.A.; Minarik, T.; Cirri, L.; Marini, L.; Perez-Gracia, J.L.; Scambia, G. A multicenter phase II study of the cryptophycin analog LY355703 in patients with platinum-resistant ovarian cancer. Int. J. Gynecol. Cancer 2006, 16, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Edelman, M.J.; Gandara, D.R.; Hausner, P.; Israel, V.; Thornton, D.; DeSanto, J.; Doyle, L.A. Phase 2 study of cryptophycin 52 (LY355703) in patients previously treated with platinum-based chemotherapy for advanced non-small cell lung cancer. Lung Cancer 2003, 39, 197–199. [Google Scholar] [CrossRef]
- Hashizume, H.; Sawa, R.; Yamashita, K.; Nishimura, Y.; Igarashi, M. Structure and antibacterial activities of new cyclic peptide antibiotics, pargamicins B, C and D, from Amycolatopsis sp. ML1-hF4. J. Antibiot. 2017, 70, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Kitani, S.; Ueguchi, T.; Igarashi, Y.; Leetanasaksakul, K.; Thamchaipenet, A.; Nihira, T. Rakicidin F, a new antibacterial cyclic depsipeptide from a marine sponge-derived Streptomyces sp. J. Antibiot. 2018, 71, 139. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shao, C.L.; Fu, X.M.; Kong, C.J.; She, Z.G.; Wang, C.Y. Lumazine Peptides Penilumamides B–D and the cyclic pentapeptide asperpeptide A from a Gorgonian-derived Aspergillus sp. Fungus. J. Nat. Prod. 2014, 77, 1601–1606. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, J.; Luo, C.; Ding, W.; Cox, D.G. A new cyclopeptide with antifungal activity from the co-culture broth of two marine mangrove fungi. Nat. Prod. Res. 2014, 28, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Ortíz-López, F.J.; Monteiro, M.C.; González-Menéndez, V.; Tormo, J.R.; Genilloud, O.; Bills, G.F.; Vicente, F.; Zhang, C.; Roemer, T.; Singh, S.B.; et al. Cyclic colisporifungin and linear cavinafungins, antifungal lipopeptides isolated from Colispora cavincola. J. Nat. Prod. 2015, 78, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.T.A.; Shaala, L.A.; Mohamed, G.A.; Badr, J.M.; Bamanie, F.H.; Ibrahim, S.R.M. Theonellamide G, a potent antifungal and cytotoxic bicyclic glycopeptide from the Red Sea marine sponge Theonella swinhoei. Mar. Drugs 2014, 12, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Semreen, M.H.; El-Gamal, M.I.; Abdin, S.; Alkhazraji, H.; Kamal, L.; Hammada, S.; El-Awady, F.; Waleed, D.; Kourbaj, L. Recent updates of marine antimicrobial peptides. Saudi Pharm. J. 2018, 26, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Yoshimura, S.; Masaki, T.; Takase, S.; Ohsumi, K.; Hashimoto, M.; Furukawa, S.; Fujie, A. ASP2397: A novel antifungal agent produced by Acremonium persicinum MF-347833. J. Antibiot. 2017, 70, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Golz, C.; Strohmann, C.; Spiteller, M. Three cyclic pentapeptides and a cyclic lipopeptide produced by endophytic Fusarium decemcellulare LG53. RSC Adv. 2016, 6, 54092–54098. [Google Scholar] [CrossRef]
- Gao, C.-H.; Chen, Y.-N.; Pan, L.-X.; Lei, F.; Long, B.; Hu, L.-Q.; Zhang, R.-C.; Ke, K.; Huang, R.-M. Two new cyclic tetrapeptides from deep-sea bacterium Bacillus amyloliquefaciens GAS 00152. J. Antibiot. 2014, 67, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.A.; Al-Lihaibi, S.S.; Alarif, W.M.; Abdel-Lateff, A.; Nogata, Y.; Washio, K.; Morikawa, M.; Okino, T. Wewakazole B, a cytotoxic cyanobactin from the Cyanobacterium Moorea producens collected in the Red Sea. J. Nat. Prod. 2016, 79, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Ebada, S.S.; Fischer, T.; Hamacher, A.; Du, F.Y.; Roth, Y.O.; Kassack, M.U.; Wang, B.G.; Roth, E.H. Psychrophilin E, a new cyclotripeptide, from co-fermentation of two marine alga-derived fungi of the genus Aspergillus. Nat. Prod. Res. 2014, 28, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Prompanya, C.; Fernandes, C.; Cravo, S.; Pinto, M.M.; Dethoup, T.; Silva, A.M.; Kijjoa, A. A new cyclic hexapeptide and a new isocoumarin derivative from the marine sponge-associated fungus Aspergillus similanensis KUFA 0013. Mar. Drugs 2015, 13, 1432–1450. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.M.; Liang, X.A.; Zhang, H.C.; Liu, R. Cytotoxic and antibiotic cyclic pentapeptide from an endophytic Aspergillus tamarii of Ficus carica. J. Agric. Food Chem. 2016, 64, 3789–3793. [Google Scholar] [CrossRef] [PubMed]
- Ványolós, A.; Dékány, M.; Kovács, B.; Krámos, B.; Bérdi, P.; Zupkó, I.; Hohmann, J.; Béni, Z. Gymnopeptides A and B, cyclic octadecapeptides from the mushroom Gymnopus fusipes. Org. Lett. 2016, 18, 2688–2691. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.H.; Wu, P.; Chi, Y.L.; Xu, L.X.; We, X.Y. Cyclopeptides from Amanita exitialis. Nat. Prod. Bioprospect. 2011, 1, 52–56. [Google Scholar] [CrossRef]
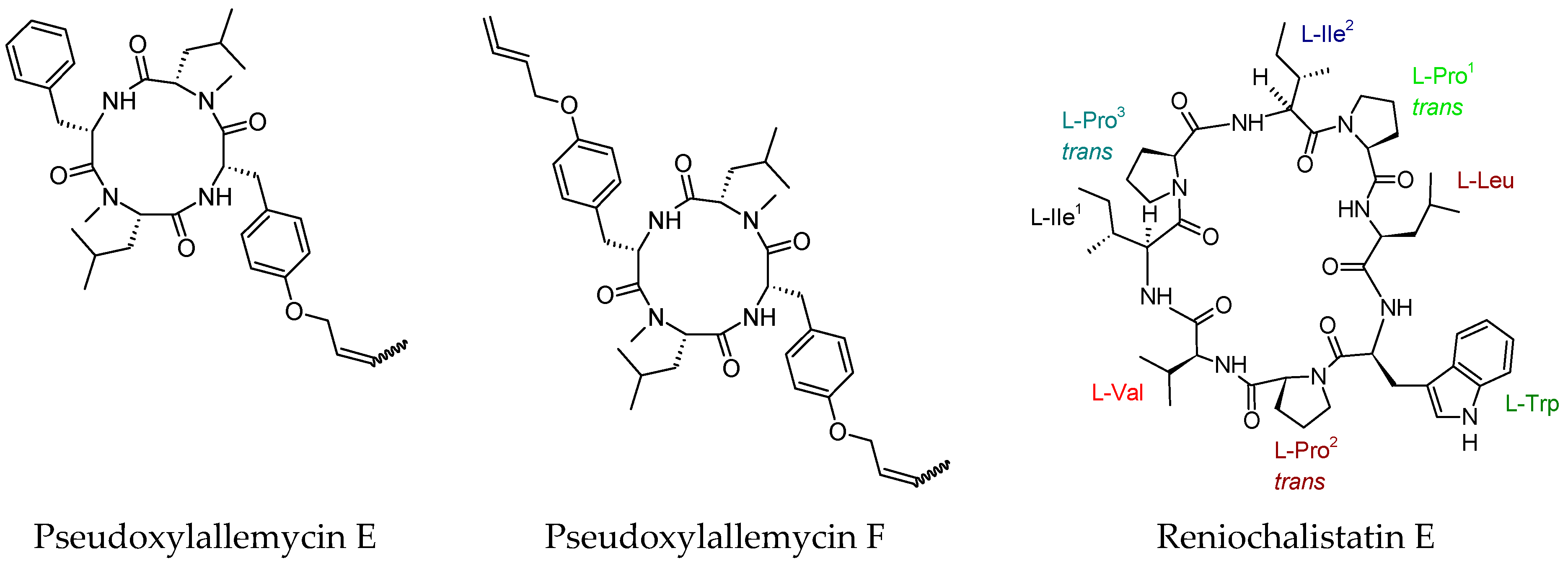
- Guo, H.; Kreuzenbeck, N.B.; Otani, S.; Garcia-Altares, M.; Dahse, H.M.; Weigel, C.; Aanen, D.K.; Hertweck, C.; Poulsen, M.; Beemelmanns, C. Pseudoxylallemycins A–F, cyclic tetrapeptides with rare allenyl modifications isolated from Pseudoxylaria sp. X802: A competitor of fungus-growing termite cultivars. Org. Lett. 2016, 18, 3338–3341. [Google Scholar] [CrossRef] [PubMed]
- Zhan, K.X.; Jiao, W.H.; Yang, F.; Li, J.; Wang, S.P.; Li, Y.S.; Han, B.N.; Lin, H.W. Reniochalistatins A–E, cyclic peptides from the marine sponge Reniochalina stalagmitis. J. Nat. Prod. 2014, 77, 2678–2684. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Min, C.C.; Teuscher, F.; Ebel, R.; Kakoschke, C.; Lin, W.; Wray, V.; Edrada-Ebel, R.; Proksch, P. Callyaerins A-F and H, new cytotoxic cyclic peptides from the Indonesian marine sponge Callyspongia aerizusa. Bioorg. Med. Chem. 2010, 18, 4947–4956. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Lee, J.H.; Won, H.; Park, S.-K.; Kim, H.M.; Shin, H.J.; Park, H.S.; Sim, C.J.; Kim, H.-K. Identification of novel acetylenic alcohols and a new dihydrothiopyranone from tropical sponge Reniochalina sp. Lipids 2009, 44, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.Y.; Jin, Y.; Yu, X.J.; Zhang, W.; Jin, M.F. Analysis of low polarity constituents of Reniochalina sp. by gas chromatography -mass spectrometry. Chin. J. Chromatogr. 2004, 22, 652–654. [Google Scholar]
- Ma, X.; Nong, X.H.; Ren, Z.; Wang, J.; Liang, X.; Wang, L.; Qi, S.H. Antiviral peptides from marine gorgonian-derived fungus Aspergillus sp. SCSIO 41501. Tetrahedron Lett. 2017, 58, 1151–1155. [Google Scholar] [CrossRef]
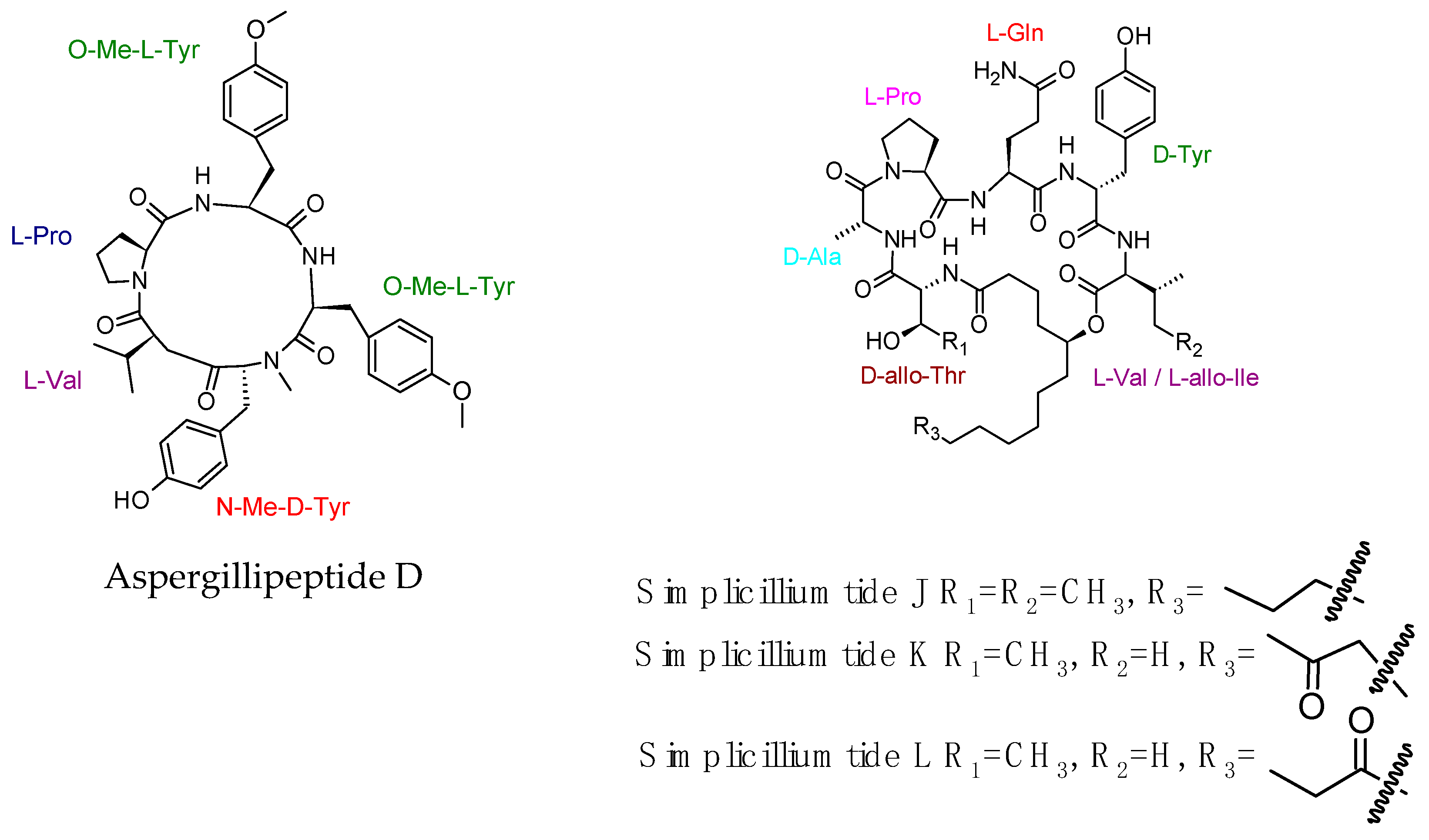
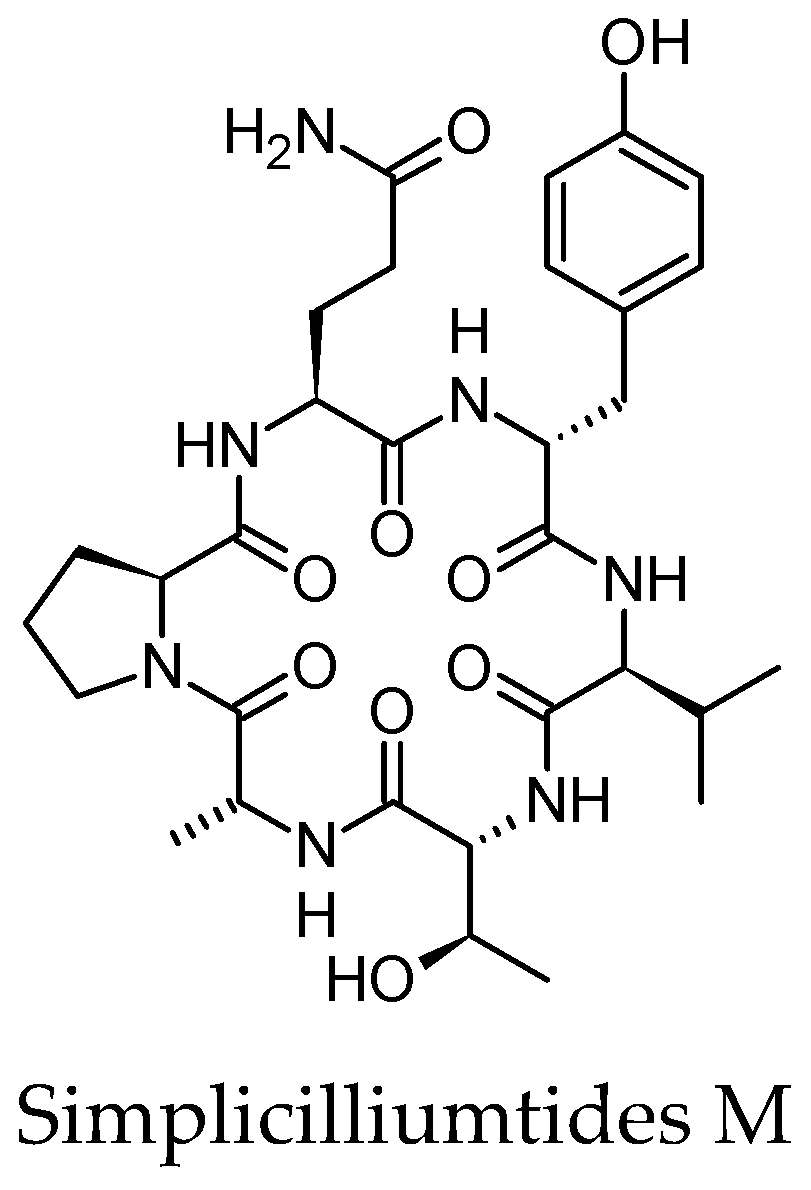
- Liang, X.; Nong, X.H.; Huang, Z.H.; Qi, S.H. Antifungal and antiviral cyclic peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. J. Agric. Food Chem. 2017, 65, 5114–5121. [Google Scholar] [CrossRef] [PubMed]
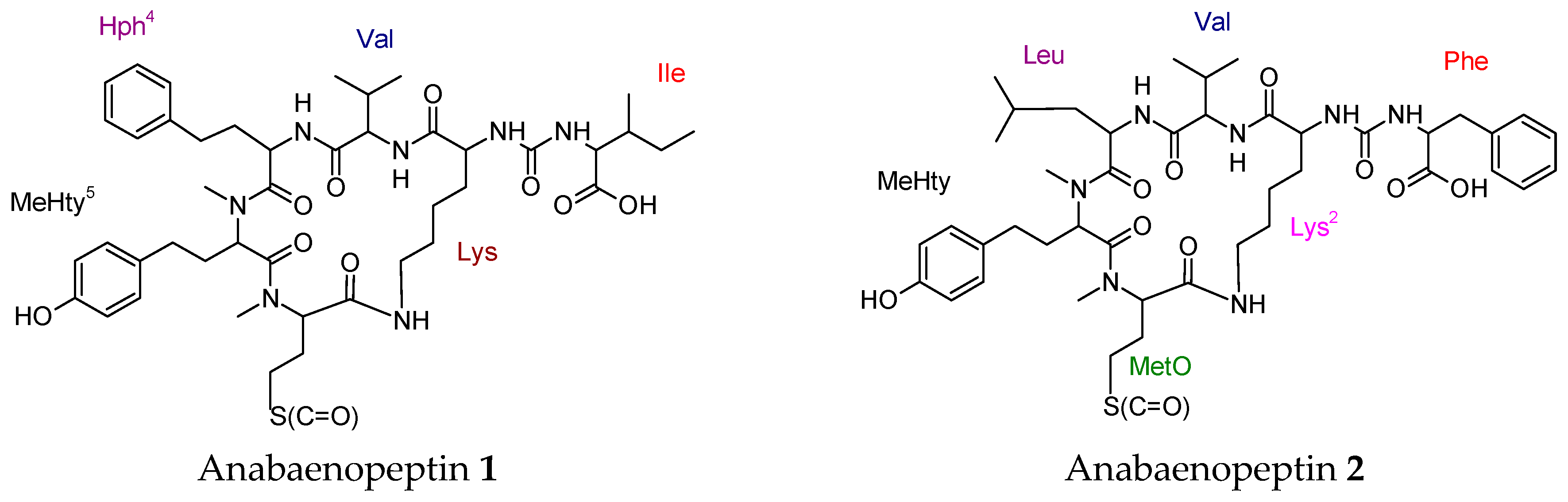
- Spoof, L.; Błaszczyk, A.; Meriluoto, J.; Cegłowska, M.; Mazur-Marzec, H. Structures and activity of new anabaenopeptins produced by baltic sea Cyanobacteria. Mar. Drugs 2015, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.P.; Hwang, G.J.; Kwon, M.C.; Ryoo, I.J.; Jang, M.; Takahashi, S.; Ko, S.K.; Osada, H.; Jang, J.H.; Ahn, J.S. Pentaminomycins A and B, hydroxyarginine-containing cyclic pentapeptides from Streptomyces sp. RK88-1441. J. Nat. Prod. 2018, 81, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.; El Maddah, F.; Kehraus, S.; Schnakenbur, G.; König, G.M. Endolides A and B, vasopressin and serotonin-receptor interacting N-methylated peptides from the sponge-derived fungus Stachylidium sp. Org. Lett. 2016, 18, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yue, Q.; Jayanetti, D.R.; Swenson, D.C.; Bartholomeusz, G.A.; An, Z.; Gloer, J.B.; Bills, G.F. Anti cryptococcus phenalenones and cyclic tetrapeptides from Auxarthron pseudauxarthron. J. Nat. Prod. 2017, 80, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Dewapriya, P.; Prasad, P.; Damodar, R.; Salim, A.A.; Capon, R.J. Talarolide A, a cyclic heptapeptide hydroxamate from an Australian marine tunicate-associated fungus, Talaromyces sp. (CMB-TU011). Org. Lett. 2017, 19, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wang, H.; Kamdem, R.S.T.; Orfali, R.S.; Dai, H.; Makhloufi, G.; Janiak, C.; Liu, Z.; Proksch, P. A new cyclohexapeptide, penitropeptide and a new polyketide, penitropone from the endophytic fungus Penicillium tropicum. Tetrahedron Lett. 2016, 57, 27–28. [Google Scholar] [CrossRef]
- May Zin, W.W.; Buttachon, S.; Dethoup, T.; Fernandes, C.; Cravo, S.; Pinto, M.M.; Gales, L.; Pereira, J.A.; Silva, A.M.; Sekeroglu, N.; et al. New cyclotetrapeptides and a new diketopiperzine derivative from the marine sponge-associated fungus Neosartorya glabra KUFA 0702. Mar. Drugs 2016, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Itoh, M.; Izumikawa, M.; Sakata, N.; Tsuchida, T.; Shin-Ya, K. Novel arginine-containing peptides MBJ-0173 and MBJ-0174 from Mortierella alpina f28740. J. Antibiot. (Tokyo) 2017, 70, 226–229. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyclic Peptide | Natural Source | Medicinal Significance | Mechanism of Action |
|---|---|---|---|
| Vancomycin | Amycolatopsis orientalis | An antibiotic mostly active against Gram-positive microorganisms, including MRSA but not vancomycin-resistant Enterococcus (VRE). It is used for bacterial prophylaxis in neurological, orthopedic, and vascular surgery. For patients who are allergic to penicillins and cephalosporins, it can be used as an alternative antibiotic [43]. | Inhibiting cell wall synthesis of bacteria by binding to the building blocks of peptidoglycan monomers of N-acetylmuramic acid and N-acetylglucosamine and blocking cross-linking of the peptidoglycan layer [44]. |
| Cyclosporin A | The fungus Tolypocladium inflatum | Has potent immunosuppressive properties. It prevents graft-versus-host disease following transplantation, rejection of kidney, heart, and liver transplants [45]. | It blocks the transcription of cytokine genes in activated T cells via the calcineurin-phosphatase pathway [46]. It binds to the cytosolic protein cyclophilin of T cells and consequently leads to reduced T-cell function. It also binds to the cyclophilin D protein that constitutes part of the mitochondrial permeability transition pore (MPTP), which causes movement of calcium ions (Ca2+) into the mitochondria and causes the contraction of the muscle cells (heart) [47]. |
| Romidepsin | Chromobacterium violaceum | A potent antitumor drug that reverses the malignancy of tumorigenic cell lines and induces apoptosis in malignant cell lines. | Promotes acetylated histones H3 and H4 in the peroxiredoxin 1 (Prdx1) promoter (as a tumor suppressor), thus activating Prdx1 expression in tumor tissues and inhibiting tumour growth [48]. |
| Griselimycin | Streptomyces bacteria | Griselimycin derivatives showed antibiotic activity in addition to oral bioavailability, absorption, and antitubercular activity [40]. Griselimycin exhibited formidable pharmacokinetic properties for its chemical class and size. | Prevents DNA replication (known as sliding clamp) by inhibiting the interaction of the replicative DNA polymerase with the DNA polymerase beta subunit [41]. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, M.A.; McGaw, L.J. Natural Cyclic Peptides as an Attractive Modality for Therapeutics: A Mini Review. Molecules 2018, 23, 2080. https://doi.org/10.3390/molecules23082080
Abdalla MA, McGaw LJ. Natural Cyclic Peptides as an Attractive Modality for Therapeutics: A Mini Review. Molecules. 2018; 23(8):2080. https://doi.org/10.3390/molecules23082080
Chicago/Turabian StyleAbdalla, Muna Ali, and Lyndy J. McGaw. 2018. "Natural Cyclic Peptides as an Attractive Modality for Therapeutics: A Mini Review" Molecules 23, no. 8: 2080. https://doi.org/10.3390/molecules23082080
APA StyleAbdalla, M. A., & McGaw, L. J. (2018). Natural Cyclic Peptides as an Attractive Modality for Therapeutics: A Mini Review. Molecules, 23(8), 2080. https://doi.org/10.3390/molecules23082080