Characterization of Small Molecules Inhibiting the Pro-Angiogenic Activity of the Zinc Finger Transcription Factor Vezf1



Abstract
1. Introduction
2. Results
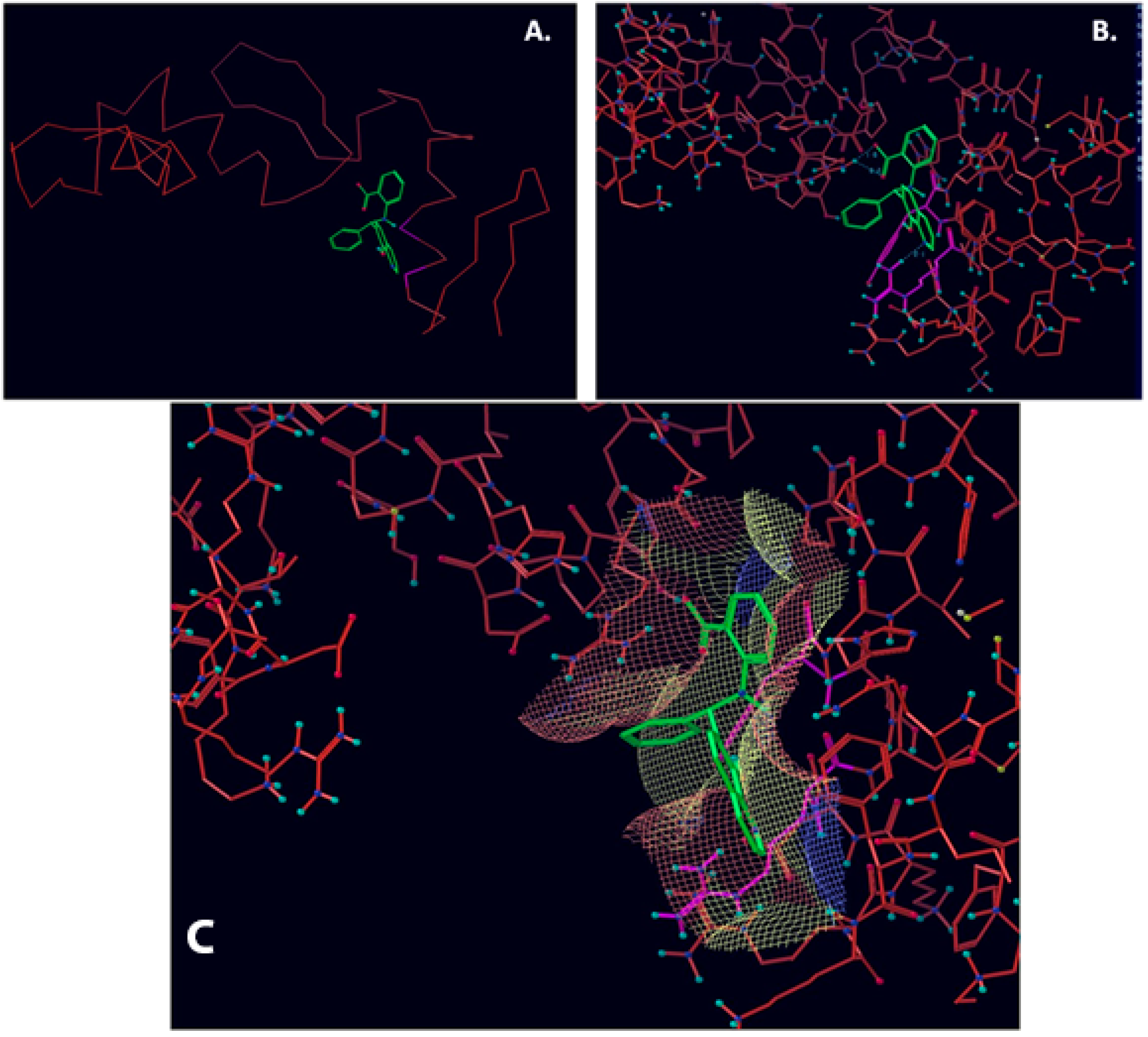
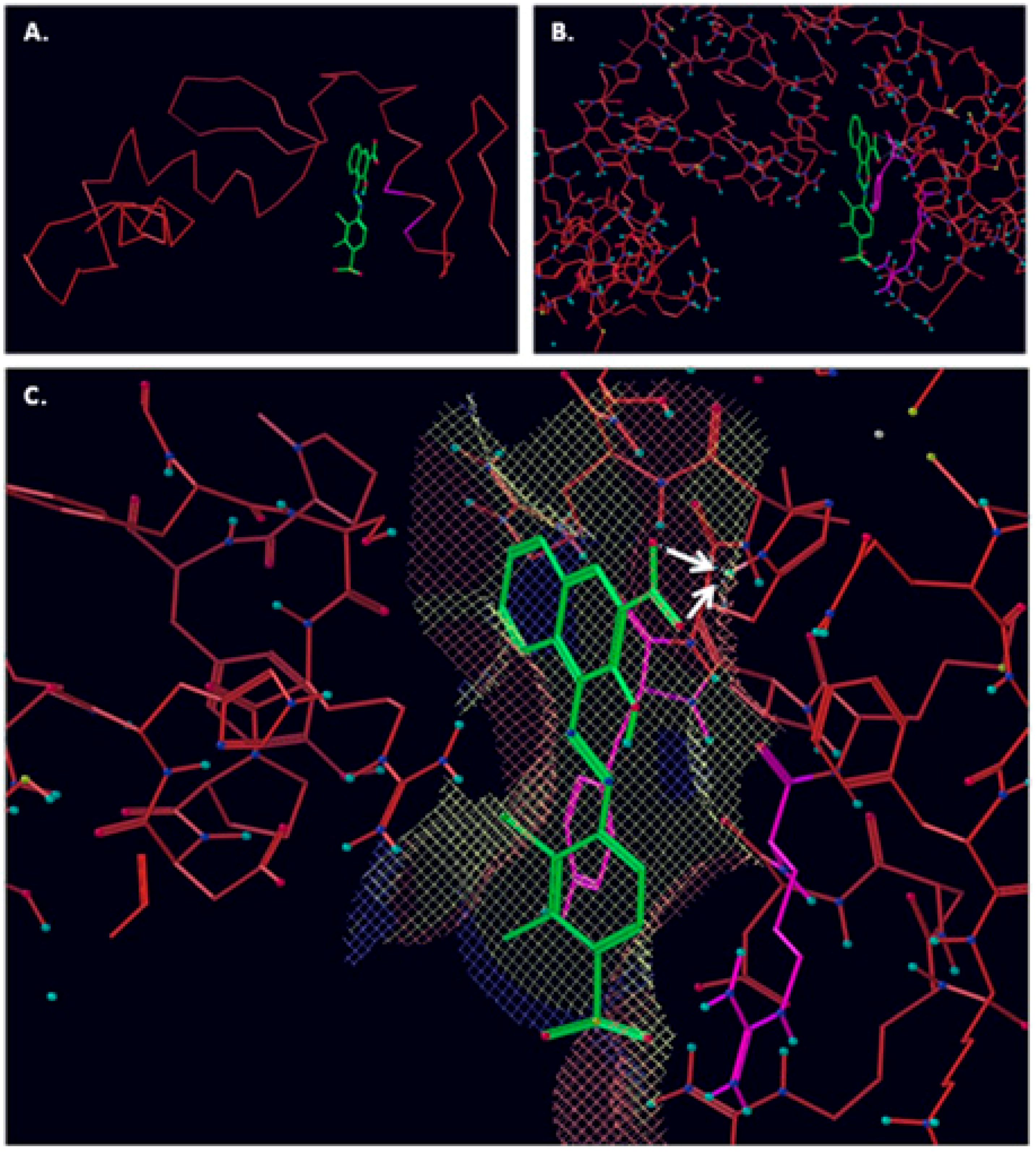
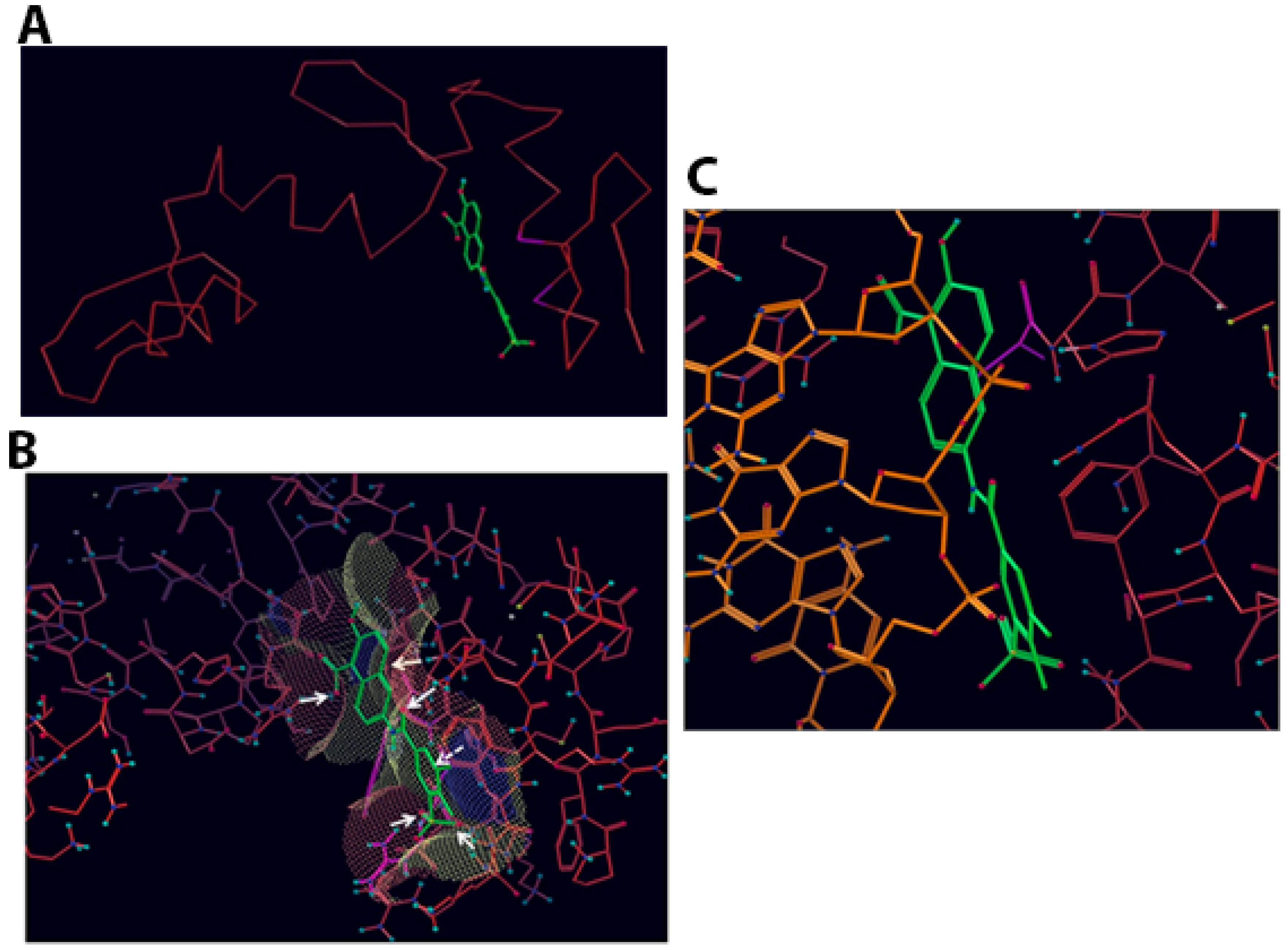
2.1. Computational Modelling of Vezf1 Structure, Evaluation of Potential Binding Sites and Design of Small Molecule Inhibitors
2.2. Virtual Screening
- (1)
- Sdock+ is a very fast docking algorithm that uses a novel method for generating vast numbers of conformers in the target-binding site. The best scoring conformers are energy minimized in the binding site.
- (2)
- McDock+ uses a Monte Carlo algorithm for generating new conformers. Conformers generated by McDock+ are optimized with energy minimization and the FLO scoring function. The calculations are much more time consuming than Sdock+, however, often the Mcdock+ results are more reliable.
- (3)
- SDock+ followed by McDock+ Conformers found by SDock+ served as a starting point for McDock+.
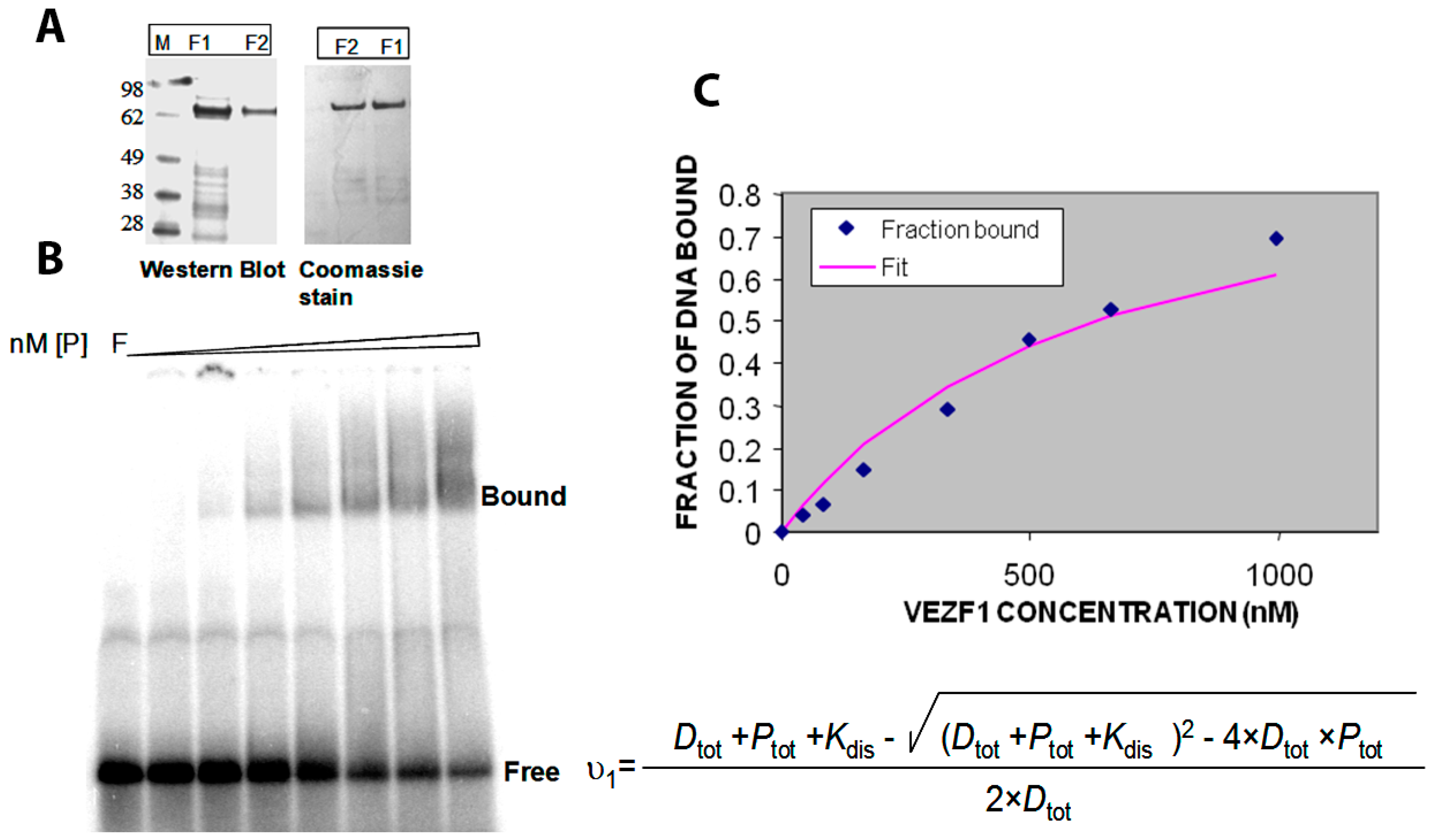
2.3. Cloning and Purification of Recombinant Vezf1
2.4. Determination of DNA Binding Constant Vezf1 to Its Specific DNA Sequence
2.5. Effect of Small Molecule Inhibitors on DNA Binding Property of Vezf1
2.6. Effect of Small Molecule Inhibitors on Cell Viability
2.7. T4 Treated MSS31 Cells Are Incapable of Tube Formation in Matrigel™
3. Discussion
4. Materials and Methods
4.1. Protein Purification and DNA Binding Assays
4.2. Small Molecule Inhibitors
4.3. Cell Viability and Tube Formation Assays
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Lewis, C.D.; Clark, S.P.; Felsenfeld, G.; Gould, H. An erythrocyte-specific protein that binds to the poly(dG) region of the chicken beta-globin gene promoter. Genes Dev. 1988, 2, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.P.; Lewis, C.D.; Felsenfeld, G. Properties of BGP1, a poly(dG)-binding protein from chicken erythrocytes. Nucleic Acids Res. 1990, 18, 5119–5126. [Google Scholar] [CrossRef] [PubMed]
- Dickson, J.; Gowher, H.; Strogantsev, R.; Gaszner, M.; Hair, A.; Felsenfeld, G.; West, A.G. VEZF1 elements mediate protection from DNA methylation. PLoS Genet. 2010, 6, e1000804. [Google Scholar] [CrossRef] [PubMed]
- Weth, O.; Renkawitz, R. CTCF function is modulated by neighboring DNA binding factors. Biochem. Cell Biol. 2011, 89, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Bruderer, M.; Alini, M.; Stoddart, M.J. Role of HOXA9 and VEZF1 in endothelial biology. J. Vasc. Res. 2013, 50, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, F.; Campagnolo, L.; Xiong, J.W.; Lemons, D.; Fitch, M.J.; Zou, Z.; Kiosses, W.B.; Gardner, H.; Stuhlmann, H. Dosage-dependent requirement for mouse Vezf1 in vascular system development. Dev. Biol. 2005, 283, 140–156. [Google Scholar] [CrossRef] [PubMed]
- Aitsebaomo, J.; Kingsley-Kallesen, M.L.; Wu, Y.; Quertermous, T.; Patterson, C. Vezf1/DB1 is an endothelial cell-specific transcription factor that regulates expression of the endothelin-1 promoter. J. Biol. Chem. 2001, 276, 39197–39205. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Ocaya, P.A.; Sun, H.; Kuhnert, F.; Stuhlmann, H. Targeted Vezf1-null mutation impairs vascular structure formation during embryonic stem cell differentiation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Gowher, H.; Brick, K.; Camerini-Otero, R.D.; Felsenfeld, G. Vezf1 protein binding sites genome-wide are associated with pausing of elongating RNA polymerase II. Proc. Natl. Acad. Sci. USA 2012, 109, 2370–2375. [Google Scholar] [CrossRef] [PubMed]
- Alabdi, L.; He, M.; Yang, Q.; Norvil, A.B.; Gowher, H. The transcription factor Vezf1 represses the expression of the antiangiogenic factor Cited2 in endothelial cells. J. Biol. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Sato, Y. Metallothionein 1 is a downstream target of vascular endothelial zinc finger 1 (VEZF1) in endothelial cells and participates in the regulation of angiogenesis. Endothel. J. Endothel. Cell Res. 2005, 12, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Kanemura, M.; Yamazaki, T.; Abe, M.; Sato, Y. Vascular endothelial zinc finger 1 is involved in the regulation of angiogenesis: Possible contribution of stathmin/OP18 as a downstream target gene. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Gerald, D.; Adini, I.; Shechter, S.; Perruzzi, C.; Varnau, J.; Hopkins, B.; Kazerounian, S.; Kurschat, P.; Blachon, S.; Khedkar, S.; et al. RhoB controls coordination of adult angiogenesis and lymphangiogenesis following injury by regulating VEZF1-mediated transcription. Nat. Commun. 2013, 4, 2824. [Google Scholar] [CrossRef] [PubMed]
- Aitsebaomo, J.; Wennerberg, K.; Der, C.J.; Zhang, C.; Kedar, V.; Moser, M.; Kingsley-Kallesen, M.L.; Zeng, G.Q.; Patterson, C. p68RacGAP is a novel GTPase-activating protein that interacts with vascular endothelial zinc finger-1 and modulates endothelial cell capillary formation. J. Biol. Chem. 2004, 279, 17963–17972. [Google Scholar] [CrossRef] [PubMed]
- Gowher, H.; Stuhlmann, H.; Felsenfeld, G. Vezf1 regulates genomic DNA methylation through its effects on expression of DNA methyltransferase Dnmt3b. Genes Dev. 2008, 22, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Mechanisms of angiogenesis and arteriogenesis. Nat. Med. 2000, 6, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Flamme, I.; Frolich, T.; Risau, W. Molecular mechanisms of vasculogenesis and embryonic angiogenesis. J. Cell. Physiol. 1997, 173, 206–210. [Google Scholar] [CrossRef]
- Eccles, S.A.; Box, C.; Court, W. Cell migration/invasion assays and their application in cancer drug discovery. Biotechnol. Annu. Rev. 2005, 11, 391–421. [Google Scholar] [PubMed]
- Gasparini, G.; Longo, R.; Fanelli, M.; Teicher, B.A. Combination of antiangiogenic therapy with other anticancer therapies: Results, challenges, and open questions. J. Clin. Oncol. 2005, 23, 1295–1311. [Google Scholar] [CrossRef] [PubMed]
- Hayden, E.C. Cutting off cancer’s supply lines. Nature 2009, 458, 686–687. [Google Scholar] [CrossRef] [PubMed]
- Pavletich, N.P.; Pabo, C.O. Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 A. Science 1991, 252, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E., Jr. Transcription factors as targets for cancer therapy. Nat. Rev. Cancer 2002, 2, 740. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.J.; Carroll, J.S. Transcription factors and chromatin proteins as therapeutic targets in cancer. Biochim. Biophys. Acta 2015, 1855, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Hegde, N.S.; Sanders, D.A.; Rodriguez, R.; Balasubramanian, S. The transcription factor FOXM1 is a cellular target of the natural product thiostrepton. Nat. Chem. 2011, 3, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, L.T.; Vu, B.T.; Graves, B.; Carvajal, D.; Podlaski, F.; Filipovic, Z.; Kong, N.; Kammlott, U.; Lukacs, C.; Klein, C.; et al. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science 2004, 303, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Ghirlando, R.; Giles, K.; Gowher, H.; Xiao, T.; Xu, Z.; Yao, H.; Felsenfeld, G. Chromatin domains, insulators, and the regulation of gene expression. Biochim. Biophys. Acta 2012, 1819, 644–651. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: The compounds can be ordered from MedPharma Partners who have standardized protocol for synthesis. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SDock+ | McDock+ | SDock+ Plus McDock+ | |
|---|---|---|---|
| M1012 | 2 | 7 | 3 |
| M16087 | 3 | 3 | 2 |
| M609974 | 12 | 15 | 17 |
| Compound Structure | Compound Number | % DNA Bound in EMSA (500 μM) | Activity Concentration |
|---|---|---|---|
 | NSC1012 (1) | 7.38% | 500 μM |
 | NSC16087 (2) | 22.56% | 500 μM |
 | NSC609974 (3) | 38.45% | 500 μM |
 | T4 | 5.94% | 20 μM |
 | T6 | 5.92% | 100 μM |
 | CID 272651 (NSC117274) | 27.13% | NA |
 | CID 23270 | 29.37% | NA |
 | T3 | 30.81% | NA |
 | T5 | 36.81% | NA |
 | T2 | 38.23% | |
 | CID 267103 | 38.95% | |
 | T1 | 43.58% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, M.; Yang, Q.; Norvil, A.B.; Sherris, D.; Gowher, H. Characterization of Small Molecules Inhibiting the Pro-Angiogenic Activity of the Zinc Finger Transcription Factor Vezf1. Molecules 2018, 23, 1615. https://doi.org/10.3390/molecules23071615
He M, Yang Q, Norvil AB, Sherris D, Gowher H. Characterization of Small Molecules Inhibiting the Pro-Angiogenic Activity of the Zinc Finger Transcription Factor Vezf1. Molecules. 2018; 23(7):1615. https://doi.org/10.3390/molecules23071615
Chicago/Turabian StyleHe, Ming, Qianyi Yang, Allison B. Norvil, David Sherris, and Humaira Gowher. 2018. "Characterization of Small Molecules Inhibiting the Pro-Angiogenic Activity of the Zinc Finger Transcription Factor Vezf1" Molecules 23, no. 7: 1615. https://doi.org/10.3390/molecules23071615
APA StyleHe, M., Yang, Q., Norvil, A. B., Sherris, D., & Gowher, H. (2018). Characterization of Small Molecules Inhibiting the Pro-Angiogenic Activity of the Zinc Finger Transcription Factor Vezf1. Molecules, 23(7), 1615. https://doi.org/10.3390/molecules23071615