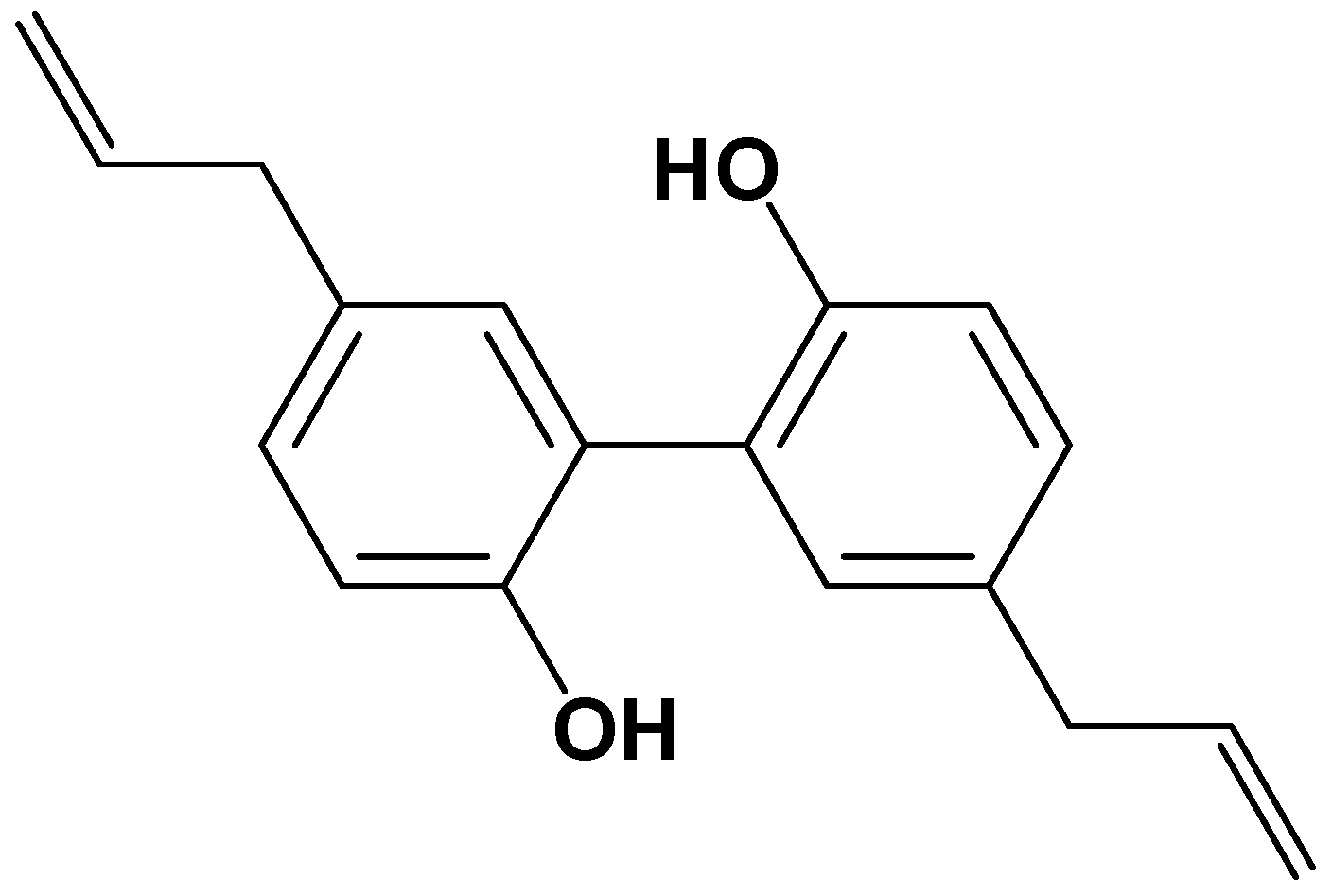
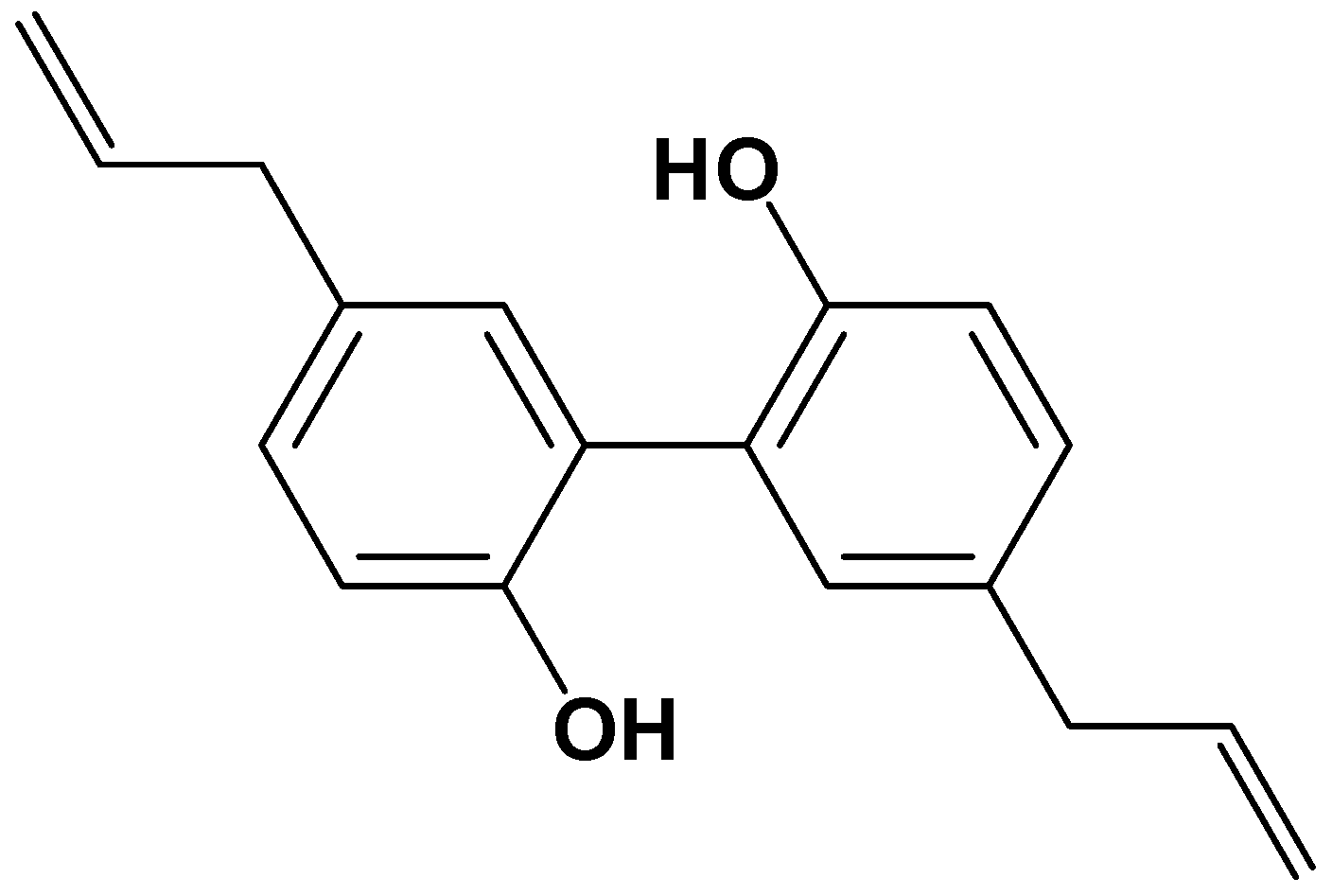
Magnolol Inhibits Osteoclast Differentiation via Suppression of RANKL Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
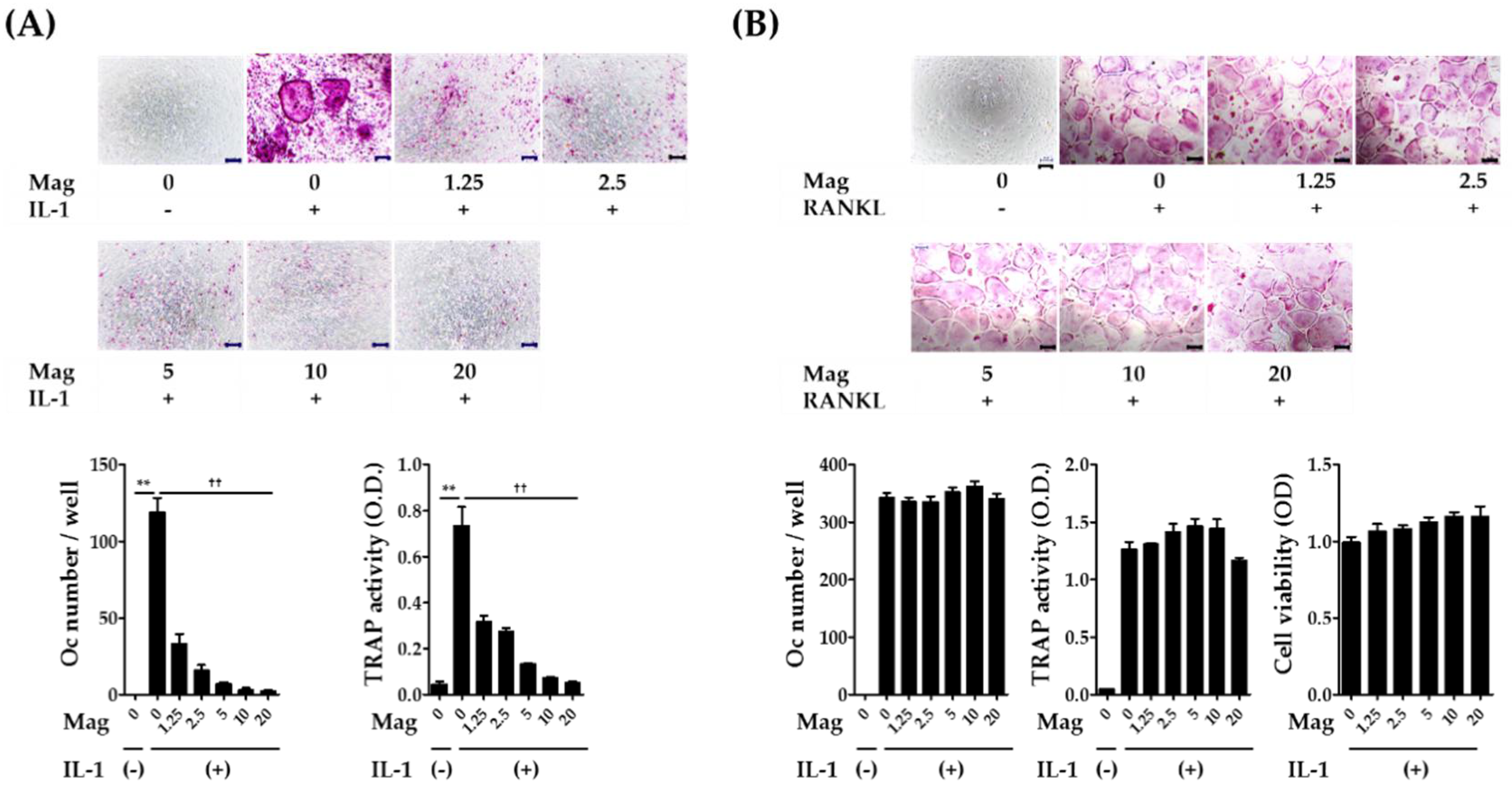
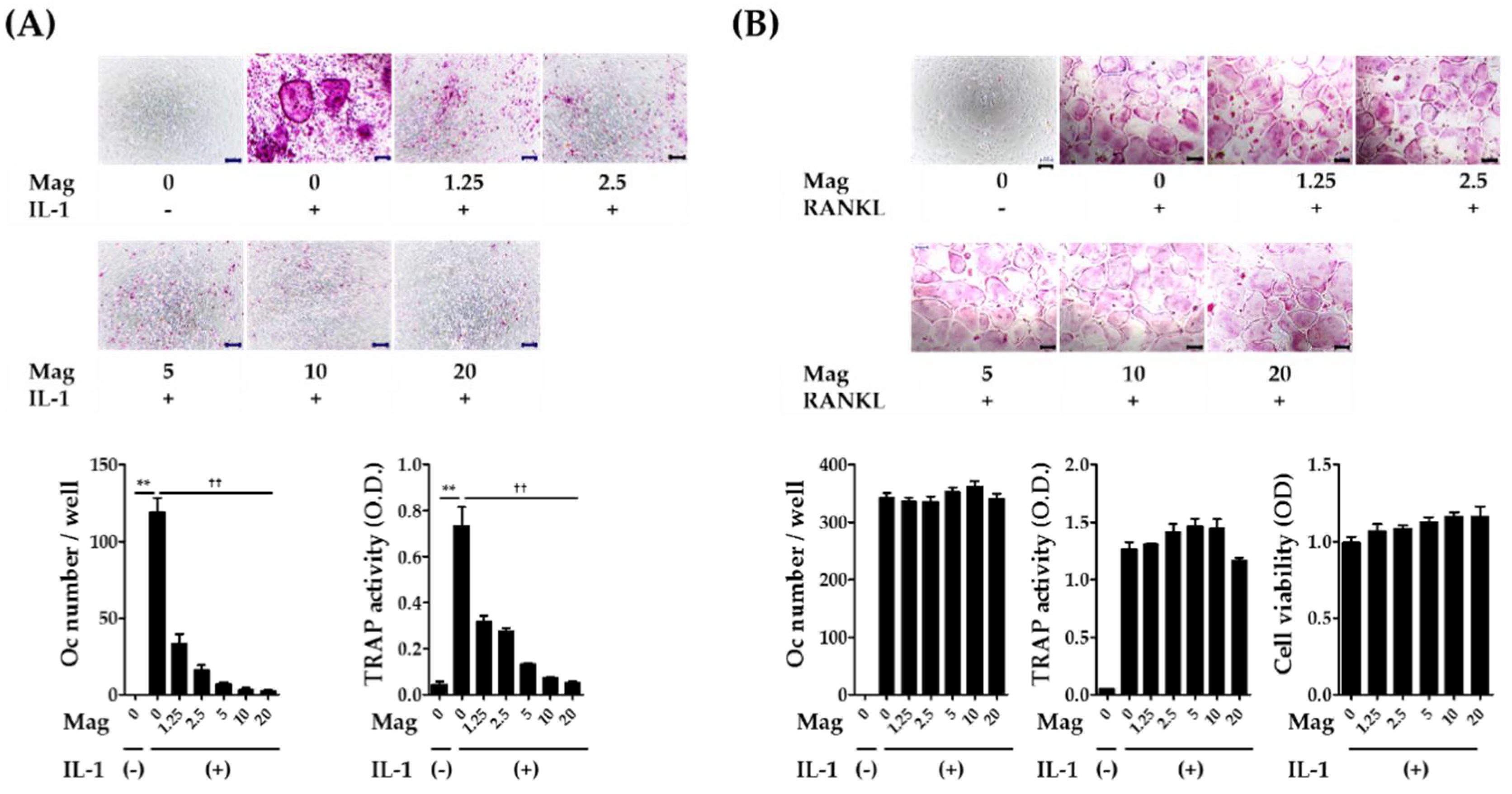
2.1. Magnolol Inhibits IL-1-Induced Osteoclast Differentiation in Co-Cultures
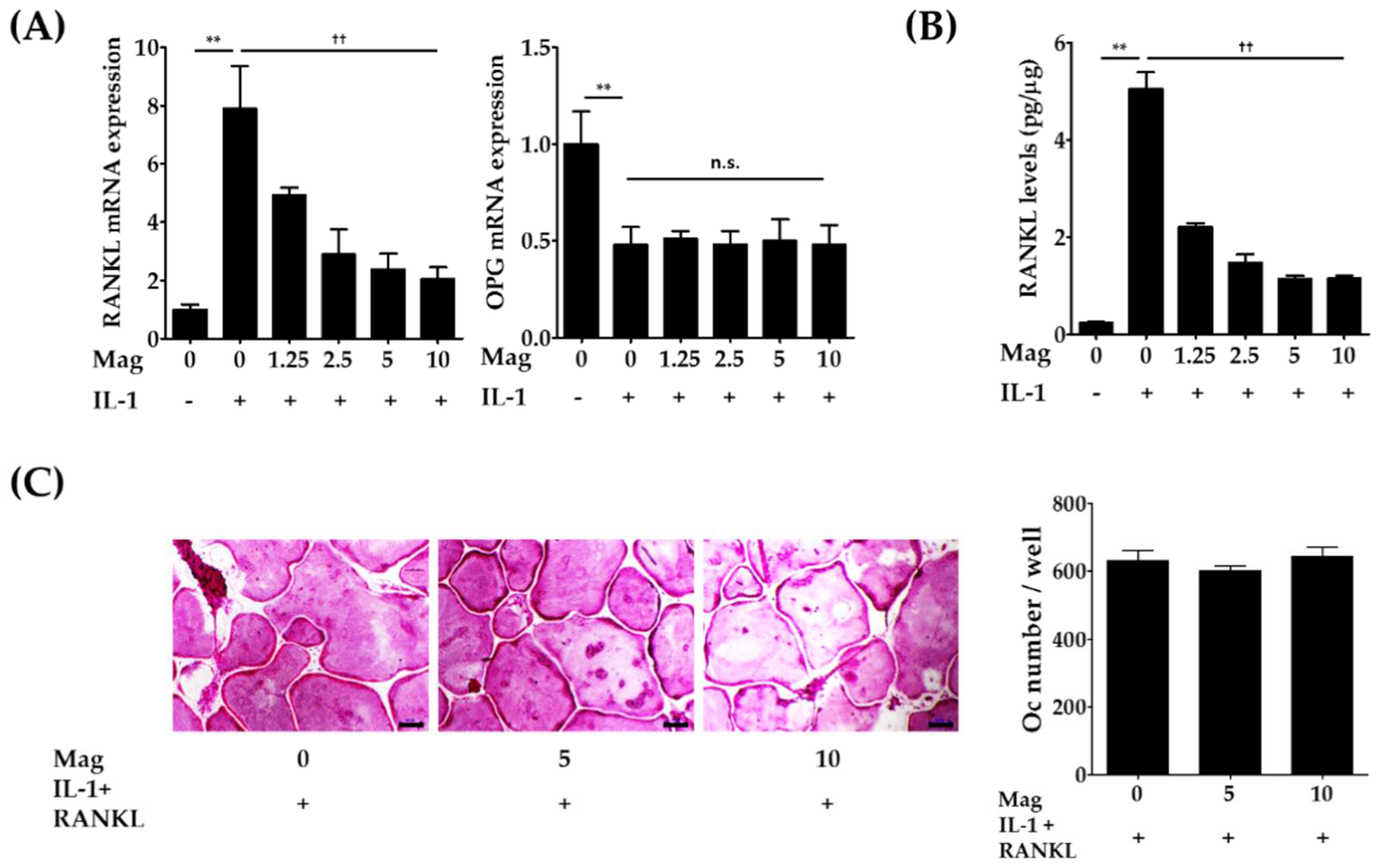
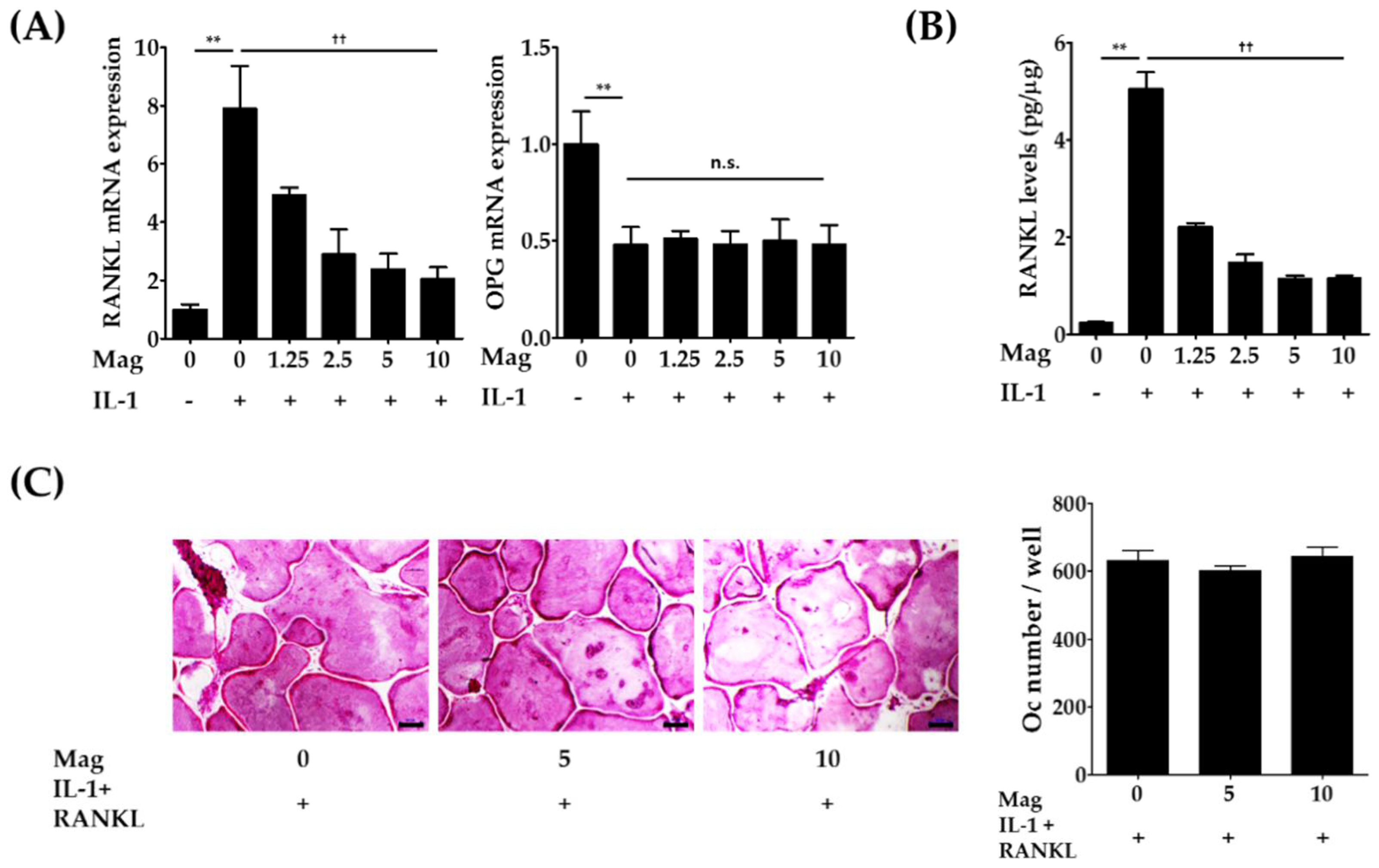
2.2. Magnolol Decreases the Expression of RANKL in IL-1-Stimulated Osteoblasts
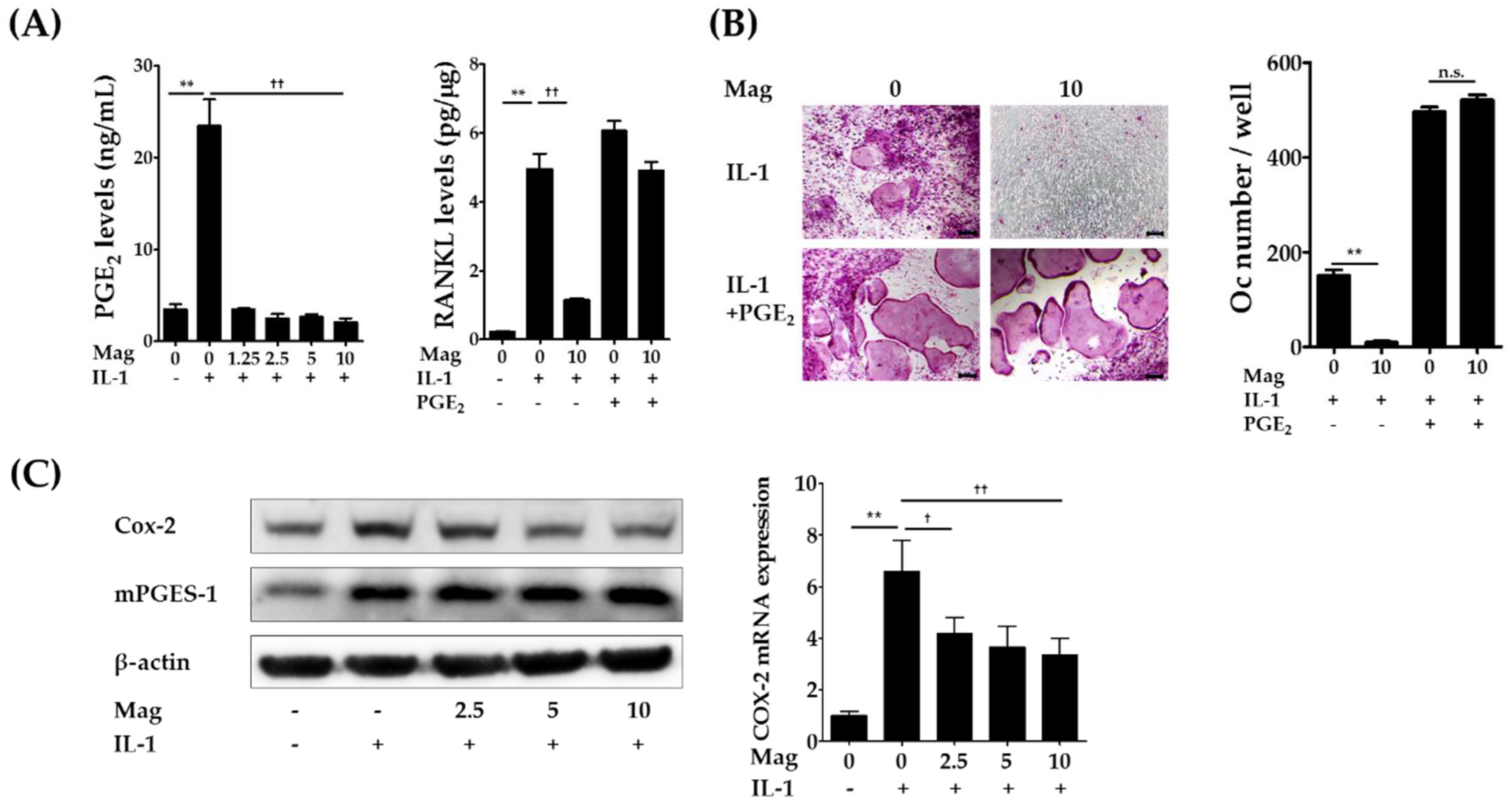
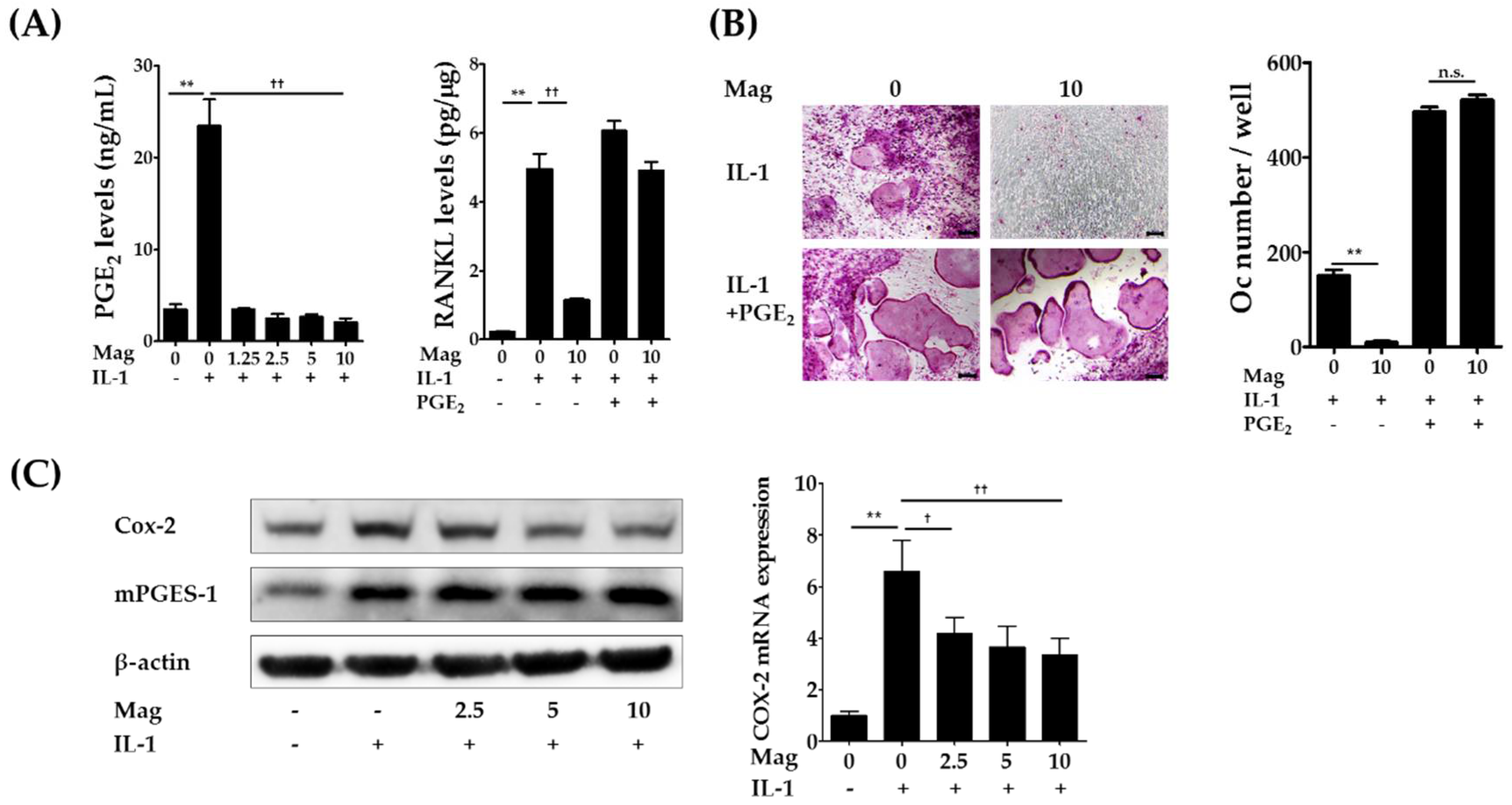
2.3. Magnolol Reduces IL-1-Induced PGE2 Production by Inhibiting Cyclooxygenase-2 (COX-2) Expression
3. Materials and Methods
3.1. Reagents
3.2. Cell Preparation
3.3. Osteoclast Formation Assays
3.4. RANKL, OPG, and PGE2 Productions in Primary Murine Osteoblasts
3.5. Real-Time Quantitative PCR
3.6. Western Blot Assay
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tanaka, S.; Nakamura, K.; Takahasi, N.; Suda, T. Role of RANKL in physiological and pathological bone resorption and therapeutics targeting the RANKL-RANK signaling system. Immunol. Res. 2005, 208, 30–49. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Zwerina, J. Positive regulators of osteoclastogenesis and bone resorption in rheumatoid arthritis. Arthritis Res. Ther. 2011, 13, e235. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Jimi, E. Regulation of osteoclast differentiation and function by interleukin-1. Vitam. Horm. 2006, 74, 357–370. [Google Scholar] [PubMed]
- Jimi, E.; Nakamura, I.; Duong, L.T.; Ikebe, T.; Takahashi, N.; Rodan, G.A.; Suda, T. Interleukin 1 induces multinucleation and bone-resorbing activity of osteoclasts in the absence of osteoblasts/stromal cells. Exp. Cell Res. 1999, 247, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.H.; Lee, S.E.; Kim, C.W.; Lee, S.H.; Kim, S.W.; Kwack, K.; Walsh, K.; Kim, H.H. IL-1α stimulation of osteoclast survival through the PI 3-kinase/Akt and ERK pathways. J. Biochem. 2002, 131, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.; Lee, J.H.; Kim, H.N.; Kim, H.M.; Kwak, H.B.; Lee, S.; Kim, H.H.; Lee, Z.H. α-Lipoic acid inhibits inflammatory bone resorption by suppressing prostaglandin E2 synthesis. J. Immunol. 2006, 176, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.C.; Zhang, S.W.; Sun, L.N.; Wang, H.; Ren, A.M. Magnolol attenuates sepsis-induced gastrointestinal dysmotility in rats by modulating inflammatory mediators. World J. Gastroenterol. 2008, 14, 7353–7360. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.H.; Huang, R.Y.; Chou, T.C. Magnolol ameliorates ligature-induced periodontitis in rats and osteoclastogenesis: In vivo and in vitro study. Evid. Based Complement. Altern. Med. 2013, 2013, e634095. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Park, H.J.; Chung, H.J.; Min, H.Y.; Park, E.J.; Lee, M.A.; Shin, Y.; Lee, S.K. Wnt/β-catenin signaling mediates the antitumor activity of magnolol in colorectal cancer cells. Mol. Pharmacol. 2012, 82, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Shih, K.S.; Liou, J.P.; Wu, Y.W.; Chang, A.S.; Wang, K.L.; Tsai, C.L.; Yang, C.R. Anti-arthritic effects of magnolol in human interleukin 1beta-stimulated fibroblast-like synoviocytes and in a rat arthritis model. PLoS ONE 2012, 7, e31368. [Google Scholar] [CrossRef]
- Lu, S.H.; Chen, T.H.; Chou, T.C. Magnolol Inhibits RANKL-induced osteoclast differentiation of raw 264.7 macrophages through heme oxygenase-1-dependent inhibition of NFATc1 expression. J. Nat. Prod. 2015, 78, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Shim, K.S.; Kim, T.; Ha, H.; Lee, C.J.; Lee, B.; Kim, H.S.; Park, J.H.; Ma, J.Y. Water extract of Magnolia officinalis cortex inhibits osteoclastogenesis and bone resorption by downregulation of nuclear factor of activated T cells cytoplasmic 1. Integr. Med. Res. 2015, 4, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.M.; Cheon, Y.H.; Kwak, S.C.; Jun, H.Y.; Yoon, K.H.; Lee, M.S.; Kim, J.Y. Claudin 11 regulates bone homeostasis via bidirectional EphB4-EphrinB2 signaling. Exp. Mol. Med. 2018, 50, e50. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Miyaura, C.; Inada, M.; Matsumoto, C.; Ohshiba, T.; Uozumi, N.; Shimizu, T.; Ito, A. An essential role of cytosolic phospholipase A2α in prostaglandin E2-mediated bone resorption associated with inflammation. Int. J. Clin. Exp. Med. 2003, 197, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Matsumoto, C.; Uematsu, S.; Akira, S.; Miyaura, C. Membrane-bound prostaglandin E synthase-1-mediated prostaglandin E2 production by osteoblast plays a critical role in lipopolysaccharide-induced bone loss associated with inflammation. J. Immunol. 2006, 177, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.H.; Chen, M.C.; Chen, T.H.; Chang, H.Y.; Chou, T.C. Magnolol ameliorates lipopolysaccharide-induced acute lung injury in rats through PPAR-γ-dependent inhibition of NF-kB activation. Int. Immunopharmacol. 2015, 28, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Kwak, E.J.; Lee, Y.S.; Choi, E.M. Effect of magnolol on the function of osteoblastic MC3T3-E1 cells. Mediat. Inflamm. 2012, 2012, e829650. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.; An, H.; Shim, K.S.; Kim, T.; Lee, K.J.; Hwang, Y.H.; Ma, J.Y. Ethanol extract of Atractylodes macrocephala protects bone loss by inhibiting osteoclast differentiation. Molecules 2013, 18, 7376–7388. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.; Shim, K.S.; Kim, T.; Lee, C.J.; Park, J.H.; Kim, H.S.; Ma, J.Y. Water extract of the fruits of Alpinia oxyphylla inhibits osteoclast differentiation and bone loss. BMC Complement. Altern. Med. 2014, 14, e352. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, Y.-H.; Kim, T.; Kim, R.; Ha, H. Magnolol Inhibits Osteoclast Differentiation via Suppression of RANKL Expression. Molecules 2018, 23, 1598. https://doi.org/10.3390/molecules23071598
Hwang Y-H, Kim T, Kim R, Ha H. Magnolol Inhibits Osteoclast Differentiation via Suppression of RANKL Expression. Molecules. 2018; 23(7):1598. https://doi.org/10.3390/molecules23071598
Chicago/Turabian StyleHwang, Youn-Hwan, Taesoo Kim, Rajeong Kim, and Hyunil Ha. 2018. "Magnolol Inhibits Osteoclast Differentiation via Suppression of RANKL Expression" Molecules 23, no. 7: 1598. https://doi.org/10.3390/molecules23071598
APA StyleHwang, Y.-H., Kim, T., Kim, R., & Ha, H. (2018). Magnolol Inhibits Osteoclast Differentiation via Suppression of RANKL Expression. Molecules, 23(7), 1598. https://doi.org/10.3390/molecules23071598