
Somatostatin Receptors as Molecular Targets in Human Uveal Melanoma

 , , ,
, , , 
Abstract
1. Introduction
2. Results
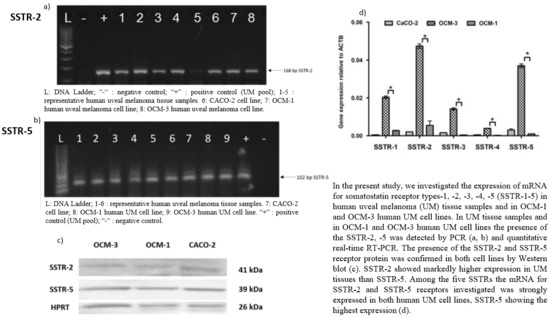
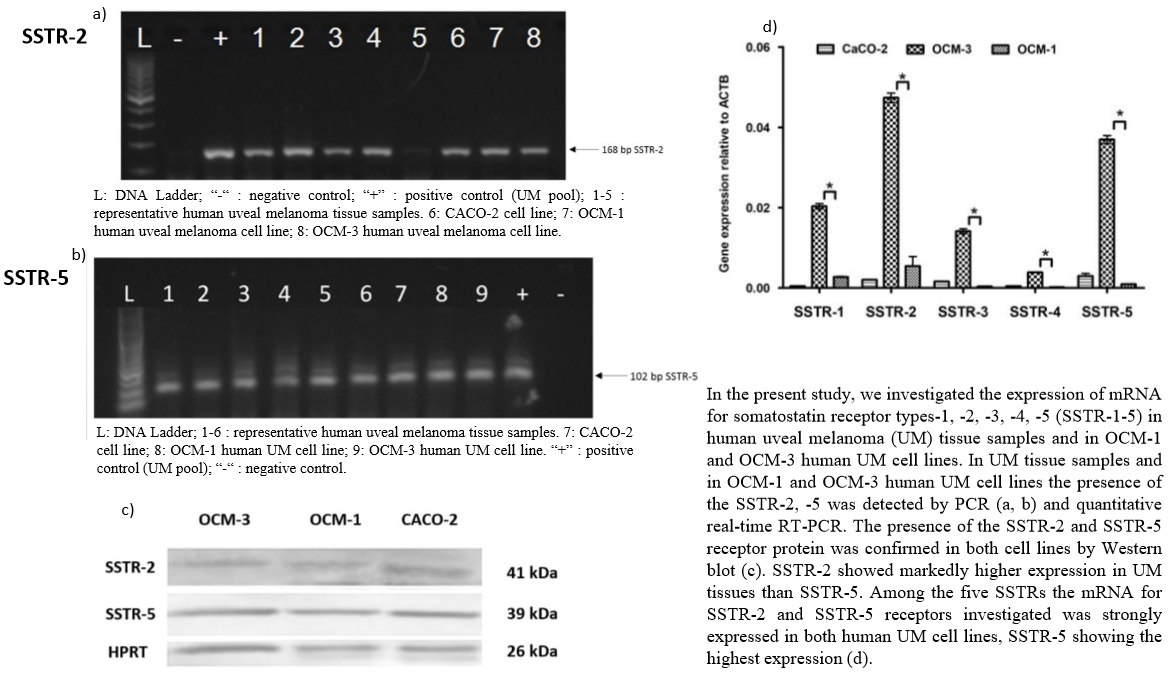
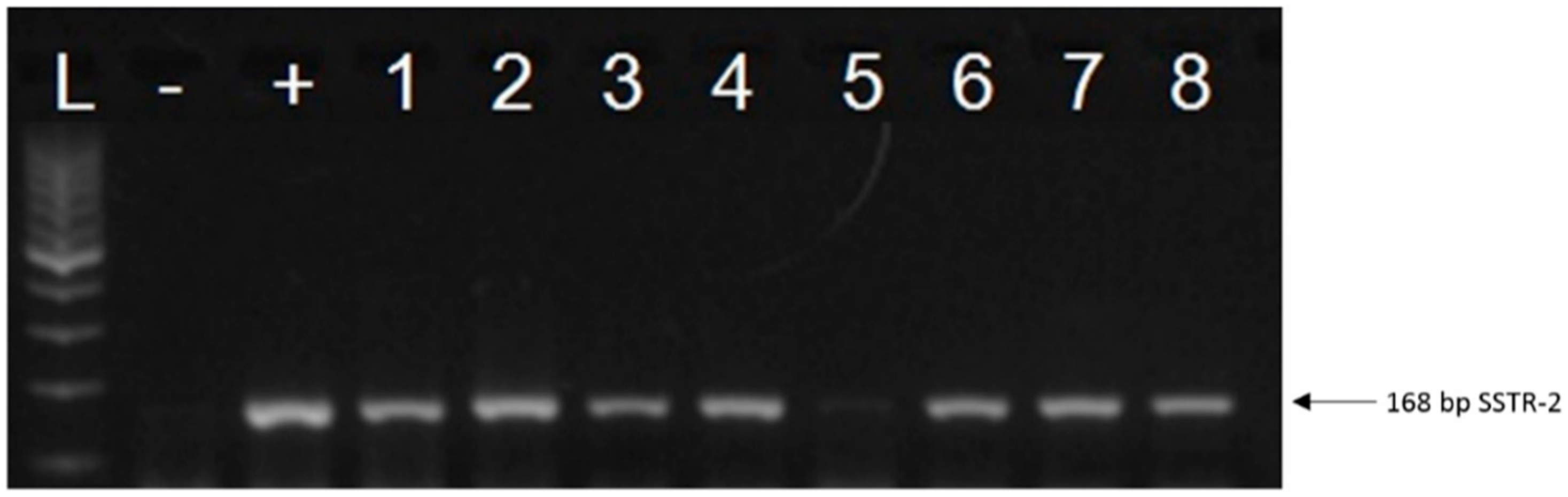
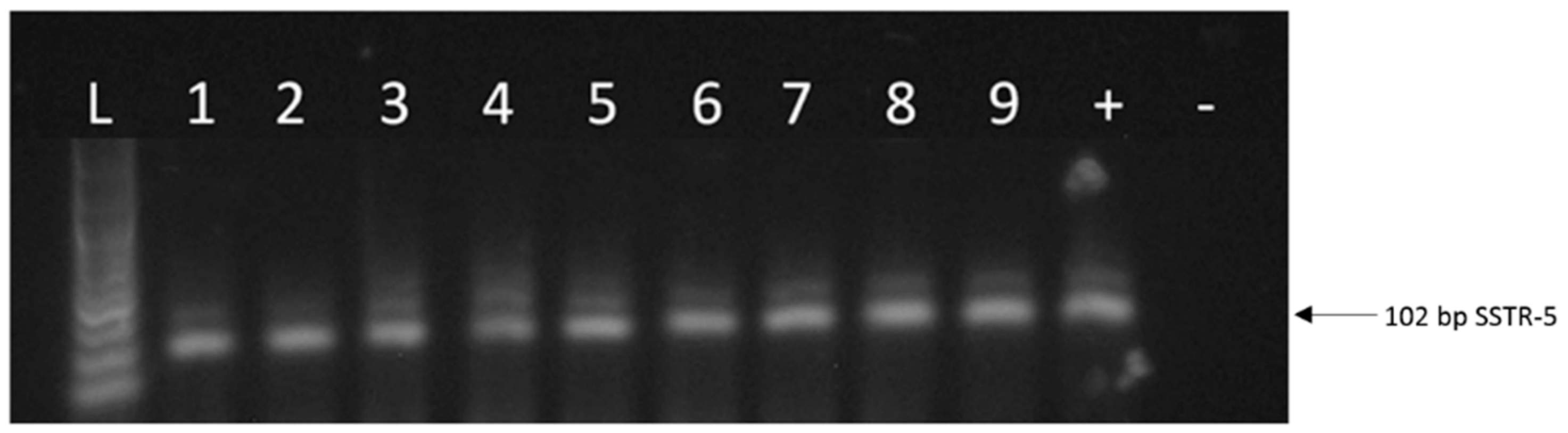
2.1. Expression of SSTRs in Human UM Tissue Samples
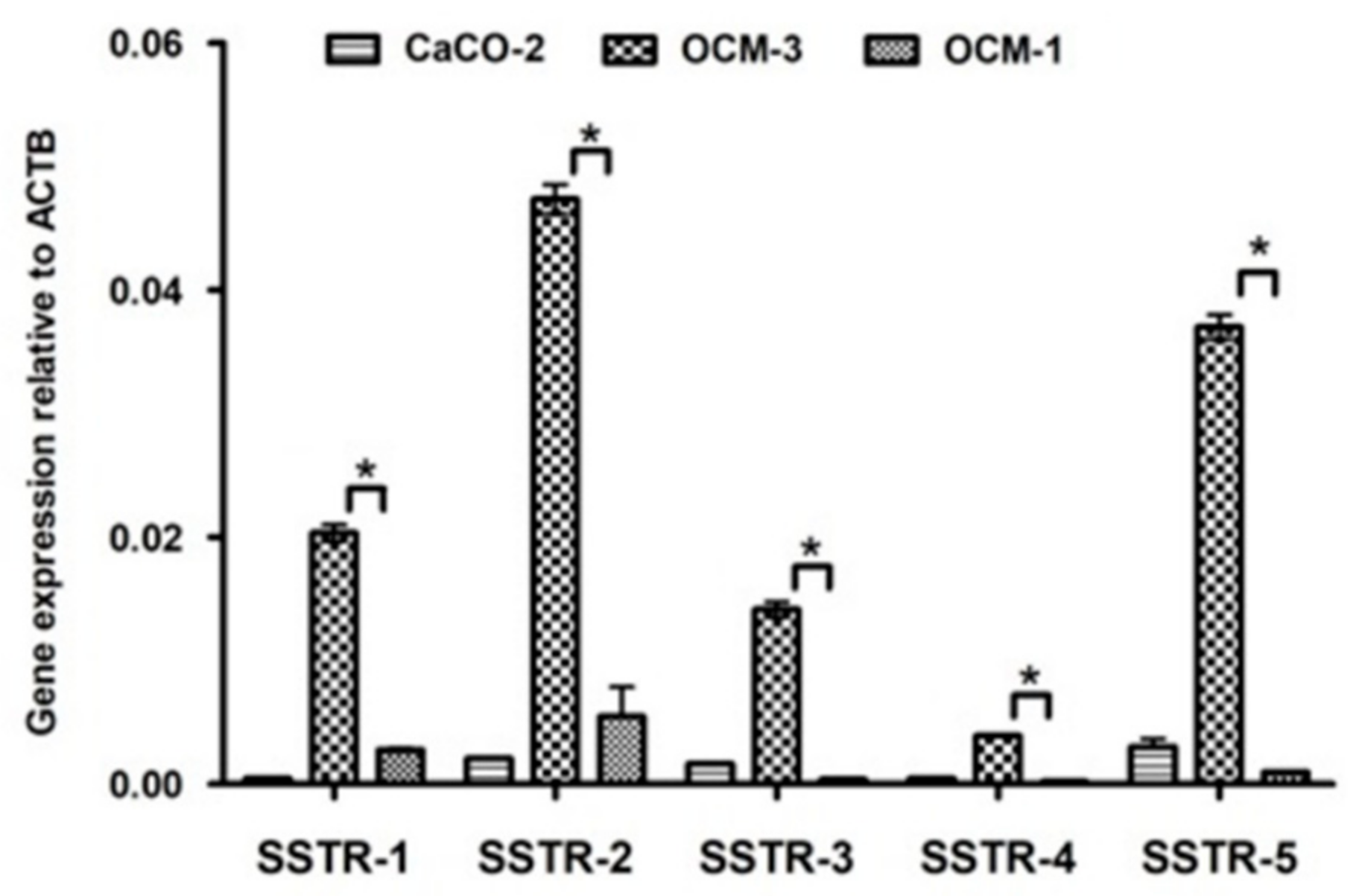
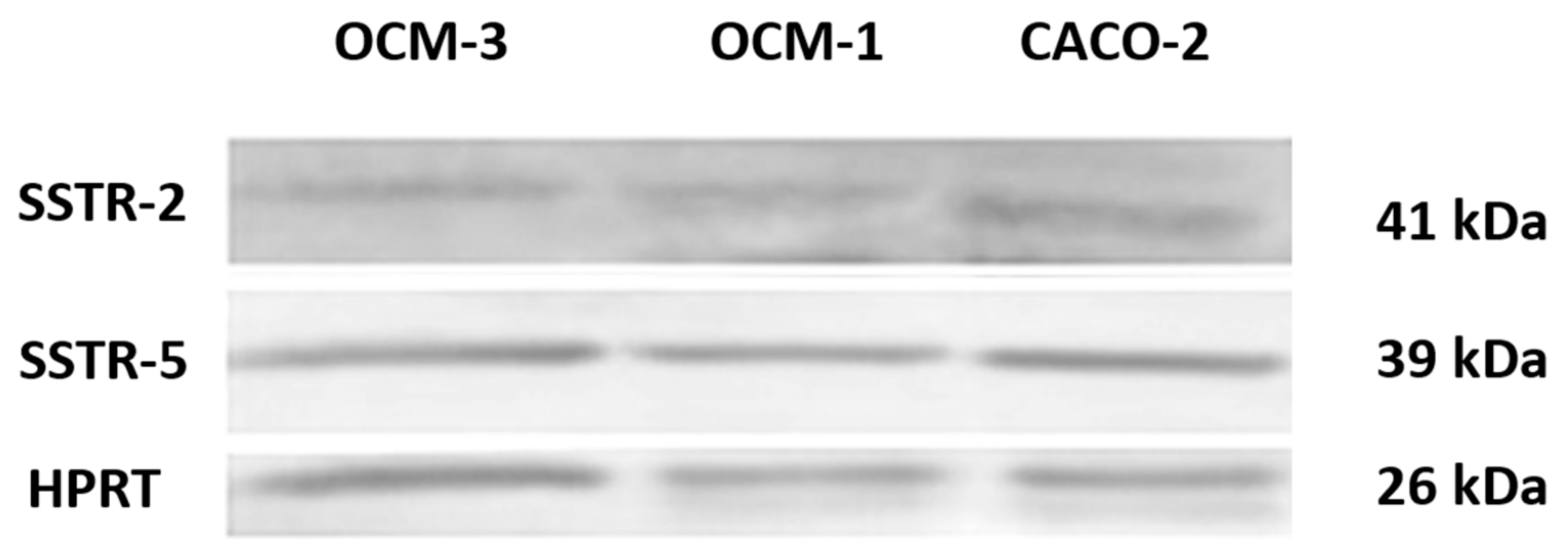
2.2. Expression of SSTRs in Human UM Cell Lines
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. Preparation of Uveal Melanoma Tissue Samples from Patients
4.3. RNA Isolation and Reverse Transcription PCR
4.4. Quantitative Real-Time PCR
4.5. Western Blot Analysis
4.6. Radioligand Binding Studies
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eskelin, S.; Pyrhonen, S.; Summanen, P.; Hahka-Kemppinen, M.; Kivela, T. Tumor doubling times in metastatic malignant melanoma of the uvea: Tumor progression before and after treatment. Ophthalmology 2000, 107, 1443–1449. [Google Scholar] [CrossRef]
- Singh, A.D.; Turell, M.E.; Topham, A.K. Uveal melanoma: Trends in incidence, treatment, and survival. Ophthalmology 2011, 118, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- Seddon, J.M.; Gragoudas, E.S.; Glynn, R.J.; Egan, K.M.; Albert, D.M.; Blitzer, P.H. Host factors, UV radiation, and risk of uveal melanoma. A case-control study. Arch. Ophthalmol. 1990, 108, 1274–1280. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.P.; Elwood, J.M.; Rootman, J.; Spinelli, J.J.; Hill, G.B.; Threlfall, W.J.; Birdsell, J.M. Risk factors for ocular melanoma: Western Canada melanoma study. J. Natl. Cancer Inst. 1985, 74, 775–778. [Google Scholar] [PubMed]
- Holly, E.A.; Aston, D.A.; Char, D.H.; Kristiansen, J.J.; Ahn, D.K. Uveal melanoma in relation to ultraviolet light exposure and host factors. Cancer Res. 1990, 50, 5773–5777. [Google Scholar] [PubMed]
- Ganley, J.P.; Comstock, G.W. Benign nevi and malignant melanomas of the choroid. Am. J. Ophthalmol. 1973, 76, 19–25. [Google Scholar] [CrossRef]
- Hammer, H.; Olah, J.; Toth-Molnar, E. Dysplastic nevi are a risk factor for uveal melanoma. Eur. J. Ophthalmol. 1996, 6, 472–474. [Google Scholar] [CrossRef] [PubMed]
- Albert, D.M.; Chang, M.A.; Lamping, K.; Weiter, J.; Sober, A. The dysplastic nevus syndrome. A pedigree with primary malignant melanomas of the choroid and skin. Ophthalmology 1985, 92, 1728–1734. [Google Scholar] [CrossRef]
- Singh, A.D.; De Potter, P.; Fijal, B.A.; Shields, C.L.; Shields, J.A.; Elston, R.C. Lifetime prevalence of uveal melanoma in white patients with oculo(dermal) melanocytosis. Ophthalmology 1998, 105, 195–198. [Google Scholar] [CrossRef]
- Duve, S.; Rakoski, J. Cutaneous melanoma in a patient with neurofibromatosis: A case report and review of the literature. Br. J. Dermatol. 1994, 131, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Antle, C.M.; Damji, K.F.; White, V.A.; Rootman, J. Uveal malignant melanoma and optic nerve glioma in von recklinghausen’s neurofibromatosis. Br. J. Ophthalmol. 1990, 74, 502–504. [Google Scholar] [CrossRef] [PubMed]
- Van den Bosch, T.; Kilic, E.; Paridaens, D.; de Klein, A. Genetics of uveal melanoma and cutaneous melanoma: Two of a kind? Dermatol. Res. Pract. 2010, 2010, 360136. [Google Scholar] [CrossRef] [PubMed]
- Amaro, A.; Gangemi, R.; Piaggio, F.; Angelini, G.; Barisione, G.; Ferrini, S.; Pfeffer, U. The biology of uveal melanoma. Cancer Metastasis Rev. 2017, 36, 109–140. [Google Scholar] [CrossRef] [PubMed]
- Donoso, L.A.; Folberg, R.; Naids, R.; Augsburger, J.J.; Shields, J.A.; Atkinson, B. Metastatic uveal melanoma. Hepatic metastasis identified by hybridoma-secreted monoclonal antibody MAb8-1h. Arch. Ophthalmol. 1985, 103, 799–801. [Google Scholar] [CrossRef] [PubMed]
- Kujala, E.; Makitie, T.; Kivela, T. Very long-term prognosis of patients with malignant uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4651–4659. [Google Scholar] [CrossRef]
- Prescher, G.; Bornfeld, N.; Hirche, H.; Horsthemke, B.; Jockel, K.H.; Becher, R. Prognostic implications of monosomy 3 in uveal melanoma. Lancet 1996, 347, 1222–1225. [Google Scholar] [PubMed]
- Sisley, K.; Rennie, I.G.; Parsons, M.A.; Jacques, R.; Hammond, D.W.; Bell, S.M.; Potter, A.M.; Rees, R.C. Abnormalities of chromosomes 3 and 8 in posterior uveal melanoma correlate with prognosis. Genes Chromosomes Cancer 1997, 19, 22–28. [Google Scholar] [CrossRef]
- Damato, B.; Duke, C.; Coupland, S.E.; Hiscott, P.; Smith, P.A.; Campbell, I.; Douglas, A.; Howard, P. Cytogenetics of uveal melanoma: A 7-year clinical experience. Ophthalmology 2007, 114, 1925–1931. [Google Scholar] [CrossRef] [PubMed]
- Dahlenfors, R.; Tornqvist, G.; Wettrell, K.; Mark, J. Cytogenetical observations in nine ocular malignant melanomas. Anticancer Res. 1993, 13, 1415–1420. [Google Scholar] [PubMed]
- Mooy, C.M.; De Jong, P.T. Prognostic parameters in uveal melanoma: A review. Surv. Ophthalmol. 1996, 41, 215–228. [Google Scholar] [CrossRef]
- Sato, T.; Han, F.; Yamamoto, A. The biology and management of uveal melanoma. Curr. Oncol. Rep. 2008, 10, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Ardjomand, N.; Ardjomand, N.; Schaffler, G.; Radner, H.; El-Shabrawi, Y. Expression of somatostatin receptors in uveal melanomas. Investig. Ophthalmol. Vis. Sci. 2003, 44, 980–987. [Google Scholar] [CrossRef]
- Ruscica, M.; Arvigo, M.; Steffani, L.; Ferone, D.; Magni, P. Somatostatin, somatostatin analogs and somatostatin receptor dynamics in the biology of cancer progression. Curr. Mol. Med. 2013, 13, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Schally, A.V.; Halmos, G. Targeting to peptide receptors. In Drug Delivery in Oncology; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2012; pp. 1219–1261. [Google Scholar]
- Treszl, A.; Steiber, Z.; Schally, A.V.; Block, N.L.; Dezso, B.; Olah, G.; Rozsa, B.; Fodor, K.; Buglyo, A.; Gardi, J.; et al. Substantial expression of luteinizing hormone-releasing hormone (LHRH) receptor type I in human uveal melanoma. Oncotarget 2013, 4, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Schally, A.V.; Comaru-Schally, A.M.; Nagy, A.; Kovacs, M.; Szepeshazi, K.; Plonowski, A.; Varga, J.L.; Halmos, G. Hypothalamic hormones and cancer. Front. Neuroendocrinol. 2001, 22, 248–291. [Google Scholar] [CrossRef] [PubMed]
- Kouch-el Filali, M.; Kilic, E.; Melis, M.; de Klein, A.; de Jong, M.; Luyten, G.P. Expression of the SST receptor 2 in uveal melanoma is not a prognostic marker. Graefe’s Arch. Clin. Exp. Ophthalmol. 2008, 246, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Benali, N.; Ferjoux, G.; Puente, E.; Buscail, L.; Susini, C. Somatostatin receptors. Digestion 2000, 62 (Suppl. 1), 27–32. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Dicitore, A.; Sciammarella, C.; Di Molfetta, S.; Rubino, M.; Faggiano, A.; Colao, A. Pasireotide in the treatment of neuroendocrine tumors: A review of the literature. Endocr.-Relat. Cancer 2018, 25, R351–R364. [Google Scholar] [CrossRef] [PubMed]
- Lamberts, S.W.; de Herder, W.W.; Hofland, L.J. Somatostatin analogs in the diagnosis and treatment of cancer. Trends Endocrinol. Metab. 2002, 13, 451–457. [Google Scholar] [CrossRef]
- Seitz, S.; Schally, A.V.; Treszl, A.; Papadia, A.; Rick, F.; Szalontay, L.; Szepeshazi, K.; Ortmann, O.; Halmos, G.; Hohla, F.; et al. Preclinical evaluation of properties of a new targeted cytotoxic somatostatin analog, AN-162 (AEZS-124), and its effects on tumor growth inhibition. Anti-Cancer Drugs 2009, 20, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.; Buchholz, S.; Schally, A.V.; Jayakumar, A.R.; Weber, F.; Papadia, A.; Rick, F.G.; Szalontay, L.; Treszl, A.; Koster, F.; et al. Targeting triple-negative breast cancer through the somatostatin receptor with the new cytotoxic somatostatin analogue AN-162 [AEZS-124]. Anti-Cancer Drugs 2013, 24, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Treszl, A.; Schally, A.V.; Seitz, S.; Szalontay, L.; Rick, F.G.; Szepeshazi, K.; Halmos, G. Inhibition of human non-small cell lung cancers with a targeted cytotoxic somatostatin analog, AN-162. Peptides 2009, 30, 1643–1650. [Google Scholar] [CrossRef] [PubMed]
- Pozsgai, E.; Schally, A.V.; Halmos, G.; Rick, F.; Bellyei, S. The inhibitory effect of a novel cytotoxic somatostatin analogue AN-162 on experimental glioblastoma. Hormone Metab. Res. 2010, 42, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Hohla, F.; Buchholz, S.; Schally, A.V.; Krishan, A.; Rick, F.G.; Szalontay, L.; Papadia, A.; Halmos, G.; Koster, F.; Aigner, E.; et al. Targeted cytotoxic somatostatin analog AN-162 inhibits growth of human colon carcinomas and increases sensitivity of doxorubicin resistant murine leukemia cells. Cancer Lett. 2010, 294, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Gilson, M.M.; Diener-West, M.; Hawkins, B.S. Comparison of survival among eligible patients not enrolled versus enrolled in the collaborative ocular melanoma study (COMS) randomized trial of pre-enucleation radiation of large choroidal melanoma. Ophthalmic Epidemiol. 2007, 14, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Elbadri, A.A.; Shaw, C.; Johnston, C.F.; Archer, D.B.; Buchanan, K.D. The distribution of neuropeptides in the ocular tissues of several mammals: A comparative study. Comp. Biochem. Physiol. C 1991, 100, 625–627. [Google Scholar] [CrossRef]
- Helboe, L.; Moller, M. Immunohistochemical localization of somatostatin receptor subtypes SST1 and SST2 in the rat retina. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2376–2382. [Google Scholar]
- Troger, J.; Sellemond, S.; Kieselbach, G.; Kralinger, M.; Schmid, E.; Teuchner, B.; Nguyen, Q.A.; Schretter-Irschick, E.; Gottinger, W. Inhibitory effect of certain neuropeptides on the proliferation of human retinal pigment epithelial cells. Br. J. Ophthalmol. 2003, 87, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.D.; Topham, A. Incidence of uveal melanoma in the United States: 1973–1997. Ophthalmology 2003, 110, 956–961. [Google Scholar] [CrossRef]
- Ossowski, L.; Aguirre-Ghiso, J.A. Dormancy of metastatic melanoma. Pigment Cell Melanoma Res. 2010, 23, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Bedikian, A.Y. Metastatic uveal melanoma therapy: Current options. Int. Ophthalmol. Clin. 2006, 46, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Buder, K.; Gesierich, A.; Gelbrich, G.; Goebeler, M. Systemic treatment of metastatic uveal melanoma: Review of literature and future perspectives. Cancer Med. 2013, 2, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Harbour, J.W. Molecular prognostic testing and individualized patient care in uveal melanoma. Am. J. Ophthalmol. 2009, 148, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.D.; Topham, A. Survival rates with uveal melanoma in the United States: 1973–1997. Ophthalmology 2003, 110, 962–965. [Google Scholar] [CrossRef]
- Abildgaard, S.K.; Vorum, H. Proteomics of uveal melanoma: A mini review. J. Oncol. 2013, 2013, 820953. [Google Scholar] [CrossRef] [PubMed]
- Nichols, E.E.; Richmond, A.; Daniels, A.B. Tumor characteristics, genetics, management, and the risk of metastasis in uveal melanoma. Semin. Ophthalmol. 2016, 31, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Sipos, E.; Dobos, N.; Rozsa, D.; Fodor, K.; Olah, G.; Szabo, Z.; Szekvolgyi, L.; Schally, A.V.; Halmos, G. Characterization of luteinizing hormone-releasing hormone receptor type I (LH-RH-I) as a potential molecular target in OCM-1 and OCM-3 human uveal melanoma cell lines. OncoTargets Ther. 2018, 11, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.C. Somatostatin and its receptor family. Front. Neuroendocrinol. 1999, 20, 157–198. [Google Scholar] [CrossRef] [PubMed]
- Bauer, W.; Briner, U.; Doepfner, W.; Haller, R.; Huguenin, R.; Marbach, P.; Petcher, T.J.; Pless, J. Sms 201-995: A very potent and selective octapeptide analogue of somatostatin with prolonged action. Life Sci. 1982, 31, 1133–1140. [Google Scholar] [CrossRef]
- Cai, R.Z.; Szoke, B.; Lu, R.; Fu, D.; Redding, T.W.; Schally, A.V. Synthesis and biological activity of highly potent octapeptide analogs of somatostatin. Proc. Natl. Acad. Sci. USA 1986, 83, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Schally, A.V.; Szepeshazi, K.; Nagy, A.; Comaru-Schally, A.M.; Halmos, G. New approaches to therapy of cancers of the stomach, colon and pancreas based on peptide analogs. Cell. Mol. Life Sci. 2004, 61, 1042–1068. [Google Scholar] [CrossRef] [PubMed]
- Bruns, C.; Lewis, I.; Briner, U.; Meno-Tetang, G.; Weckbecker, G. Som230: A novel somatostatin peptidomimetic with broad somatotropin release inhibiting factor (SRIF) receptor binding and a unique antisecretory profile. Eur. J. Endocrinol. 2002, 146, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C.; Laissue, J.A. Multiple actions of somatostatin in neoplastic disease. Trends Pharmacol. Sci. 1995, 16, 110–115. [Google Scholar] [CrossRef]
- Krenning, E.P.; Kwekkeboom, D.J.; Bakker, W.H.; Breeman, W.A.; Kooij, P.P.; Oei, H.Y.; van Hagen, M.; Postema, P.T.; de Jong, M.; Reubi, J.C.; et al. Somatostatin receptor scintigraphy with [111In-DTPA-D-Phe1]-and [123I-Tyr3]-octreotide: The rotterdam experience with more than 1000 patients. Eur. J. Nucl. Med. 1993, 20, 716–731. [Google Scholar] [CrossRef] [PubMed]
- Kwekkeboom, D.J.; Krenning, E.P.; Bakker, W.H.; Oei, H.Y.; Kooij, P.P.; Lamberts, S.W. Somatostatin analogue scintigraphy in carcinoid tumours. Eur. J. Nucl. Med. 1993, 20, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Schally, A.V.; Engel, J.B.; Emons, G.; Block, N.L.; Pinski, J. Use of analogs of peptide hormones conjugated to cytotoxic radicals for chemotherapy targeted to receptors on tumors. Curr. Drug Deliv. 2011, 8, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Schally, A.V.; Nagy, A. Chemotherapy targeted to cancers through tumoral hormone receptors. Trends Endocrinol. Metab. 2004, 15, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Halmos, G.; Schally, A.V.; Sun, B.; Davis, R.; Bostwick, D.G.; Plonowski, A. High expression of somatostatin receptors and messenger ribonucleic acid for its receptor subtypes in organ-confined and locally advanced human prostate cancers. J. Clin. Endocrinol. Metab. 2000, 85, 2564–2571. [Google Scholar] [PubMed]
Sample Availability: not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Case | Age (Years) | Sex | Histology | mRNA for SSTR-2 | mRNA for SSTR-5 | SSTR Binding | |
|---|---|---|---|---|---|---|---|
| Kd (nM) | Bmax (fmol/mg Protein) | ||||||
| 1 | 30 | female | Spindle | − | N/A | N/A | N/A |
| 2 | 33 | female | Spindle | + | N/A | N/A | N/A |
| 3 | 35 | male | Spindle | − | N/A | N/A | N/A |
| 4 | 38 | male | Spindle | + | N/A | 6.42 | 304 |
| 5 | 39 | male | N/A | + | N/A | N/A | N/A |
| 6 | 39 | male | Epithelioid | − | + | 9.19 | 527 |
| 7 | 43 | male | Spindle | + | N/A | N/A | N/A |
| 8 | 45 | male | Spindle and Epithelioid | + | N/A | N/A | N/A |
| 9 | 47 | male | Epithelioid | − | − | − | − |
| 10 | 47 | male | Epithelioid | + | + | 8.53 | 1052 |
| 11 | 50 | female | Spindle | + | N/A | N/A | N/A |
| 12 | 50 | female | Spindle | + | N/A | N/A | N/A |
| 13 | 52 | male | Spindle | + | N/A | N/A | N/A |
| 14 | 53 | male | Epithelioid | + | N/A | 6.24 | 260 |
| 15 | 53 | male | N/A | − | N/A | N/A | N/A |
| 17 | 60 | male | Epithelioid | + | N/A | N/A | N/A |
| 18 | 61 | male | Spindle | + | N/A | N/A | N/A |
| 19 | 62 | female | Epithelioid | + | N/A | 5.58 | 583 |
| 20 | 64 | male | N/A | + | N/A | N/A | N/A |
| 21 | 64 | male | Spindle | − | + | 4.75 | 482 |
| 22 | 65 | male | Spindle | − | + | 3.18 | 467 |
| 23 | 66 | male | Epithelioid | + | N/A | N/A | N/A |
| 24 | 67 | male | Epithelioid | + | N/A | 5.04 | 509 |
| 25 | 68 | female | Spindle | − | N/A | − | − |
| 26 | 68 | male | Spindle | + | N/A | N/A | N/A |
| 27 | 68 | female | Epithelioid | + | N/A | N/A | N/A |
| 28 | 69 | male | Spindle and Epithelioid | + | N/A | 9.48 | 774 |
| 29 | 70 | male | Spindle and Epithelioid | + | N/A | N/A | N/A |
| 30 | 70 | male | Spindle | + | N/A | 4.37 | 985 |
| 31 | 70 | male | Spindle | − | − | − | − |
| 32 | 72 | female | Epithelioid | + | N/A | N/A | N/A |
| 33 | 72 | male | Epithelioid | + | N/A | 11.8 | 473 |
| 34 | 72 | male | Epithelioid | − | N/A | N/A | N/A |
| 35 | 75 | female | Spindle | − | N/A | − | − |
| 36 | 75 | female | Epithelioid | + | N/A | N/A | N/A |
| 37 | 75 | female | Spindle | + | N/A | 11.1 | 668 |
| 38 | 75 | female | Spindle | − | + | 3.57 | 392 |
| 39 | 76 | male | Spindle | − | N/A | − | − |
| 40 | 76 | male | Spindle | + | N/A | N/A | N/A |
| 41 | 76 | male | Spindle | − | + | 10.7 | 980 |
| 42 | 79 | female | Epithelioid | + | N/A | N/A | N/A |
| 43 | 79 | female | N/A | + | N/A | N/A | N/A |
| 44 | 79 | female | N/A | + | N/A | N/A | N/A |
| 45 | 80 | female | Epithelioid | − | − | − | − |
| 46 | 84 | female | Spindle | + | N/A | N/A | N/A |
| Target | Forward Sequence | Reverse Sequence |
|---|---|---|
| SSTR-1 | tgagtcagctgtcggtcatc | ggaaagagcgcttgaagttg |
| SSTR-2 | ctttgtggtggtcctcacct | gcagaggacattctggaagc |
| SSTR-3 | ttcctctcctaccgcttcaa | ctcctcctcatcctcctcct |
| SSTR-4 | tctttgtgctctgctggatg | ggataagggacacgtggttg |
| SSTR-5 | tctttgtgctctgctggatg | gttggcgtaggagaggatga |
| β-actin | ggcatcctcaccctgaagta | ggggtgttgaaggtctcaaa |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harda, K.; Szabo, Z.; Szabo, E.; Olah, G.; Fodor, K.; Szasz, C.; Mehes, G.; Schally, A.V.; Halmos, G. Somatostatin Receptors as Molecular Targets in Human Uveal Melanoma. Molecules 2018, 23, 1535. https://doi.org/10.3390/molecules23071535
Harda K, Szabo Z, Szabo E, Olah G, Fodor K, Szasz C, Mehes G, Schally AV, Halmos G. Somatostatin Receptors as Molecular Targets in Human Uveal Melanoma. Molecules. 2018; 23(7):1535. https://doi.org/10.3390/molecules23071535
Chicago/Turabian StyleHarda, Kristof, Zsuzsanna Szabo, Erzsebet Szabo, Gabor Olah, Klara Fodor, Csaba Szasz, Gabor Mehes, Andrew V. Schally, and Gabor Halmos. 2018. "Somatostatin Receptors as Molecular Targets in Human Uveal Melanoma" Molecules 23, no. 7: 1535. https://doi.org/10.3390/molecules23071535
APA StyleHarda, K., Szabo, Z., Szabo, E., Olah, G., Fodor, K., Szasz, C., Mehes, G., Schally, A. V., & Halmos, G. (2018). Somatostatin Receptors as Molecular Targets in Human Uveal Melanoma. Molecules, 23(7), 1535. https://doi.org/10.3390/molecules23071535