Ponasterone A and F, Ecdysteroids from the Arctic Bryozoan Alcyonidium gelatinosum

 , and
, and
Abstract
1. Introduction
2. Results and Discussion
2.1. Biomass Collection, Extraction and Fractionation
2.2. Viability Screening of the Flash Fractions
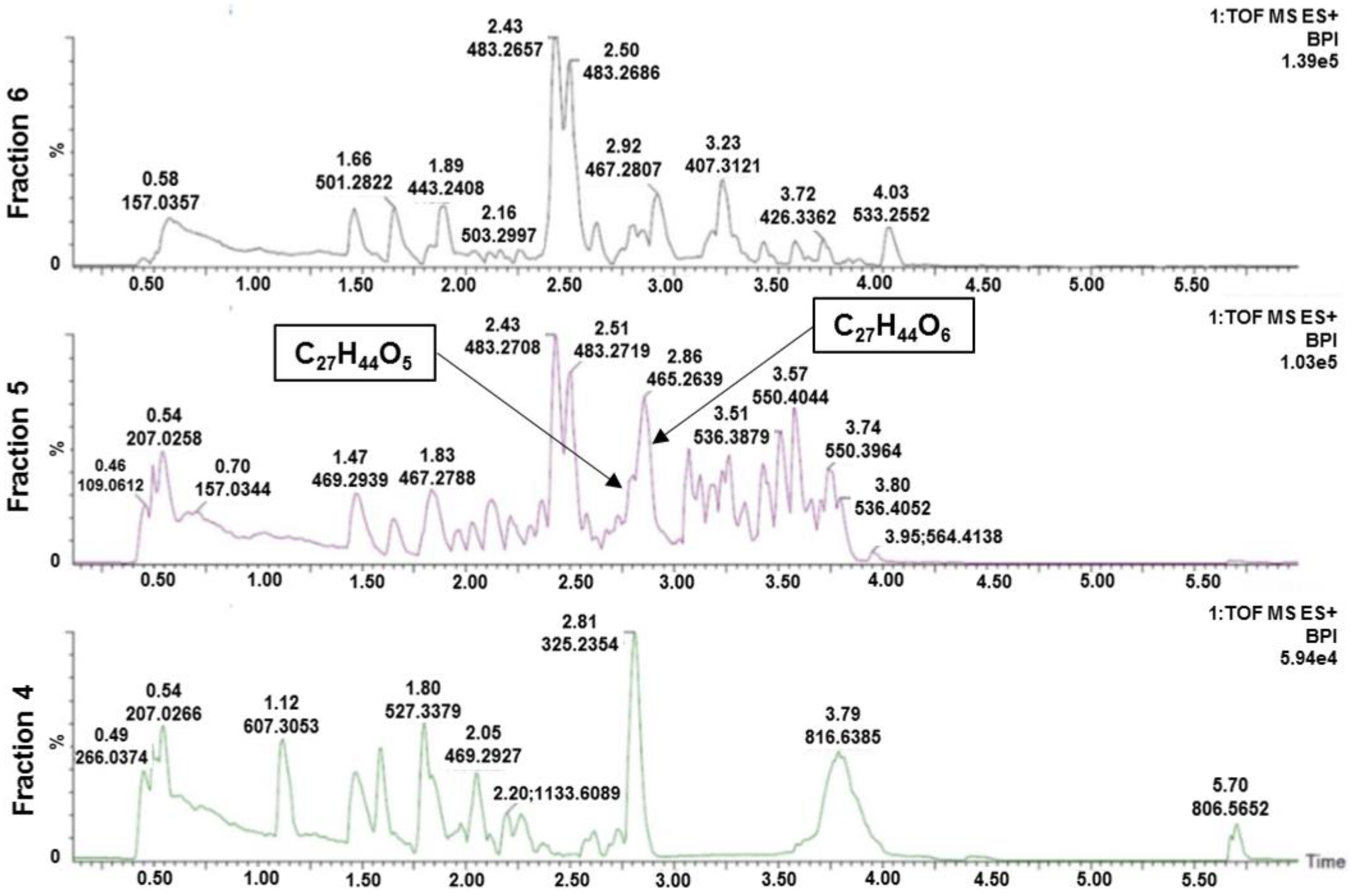
2.3. Dereplication of the Active Fraction
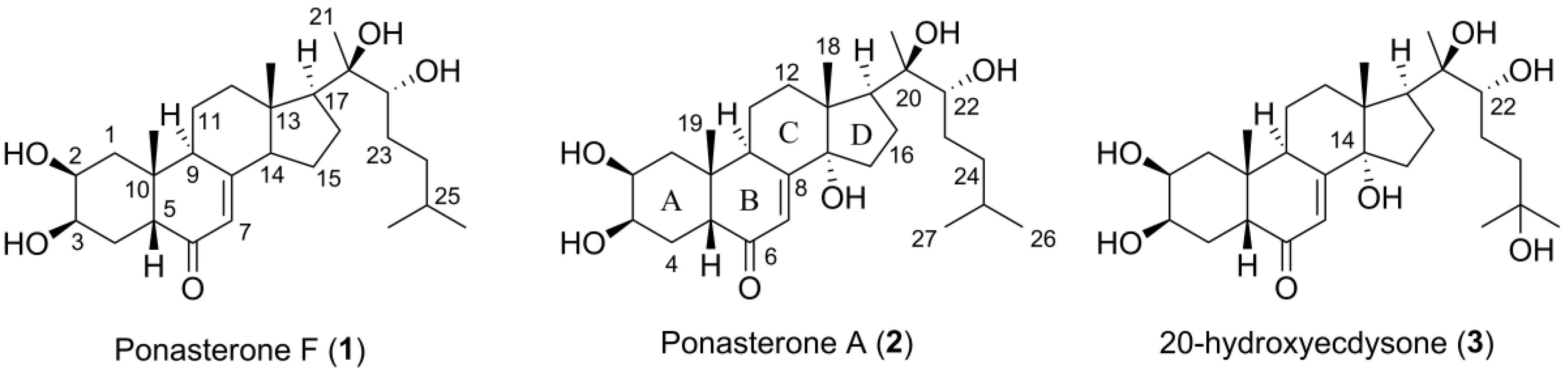
2.4. Isolation of 1 and 2
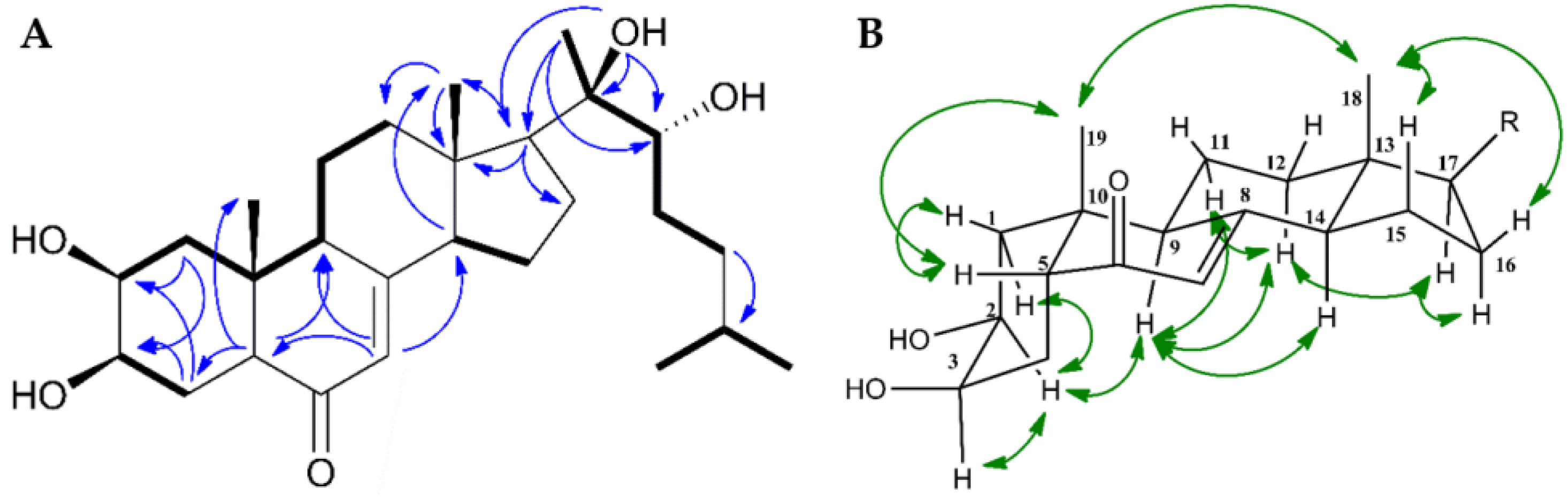
2.5. Structure Elucidation of Compound 1
2.6. Estrogen Receptor Agonist Activities of 1 and 2
2.7. Antiproliferative and Antibacterial Properties of 1 and 2
2.8. Probable Natural Functions of 1 and 2 in A. gelatinosum
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Extraction and Fractionation
3.4. Cytotoxicity Screening (MTS Assay) of the Flash Fractions
3.5. Dereplication of the Active Fraction
3.6. Isolation of 1 and 2
3.7. Agonistic Activity of 1 and 2 against Estrogen Receptors Alpha and Beta
3.8. Antiproliferative and Antibacterial Properties of 1 and 2
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Connor, J.D. Ecdysteroid action at the molecular level. In Comprehensive Insect Physiology and Biochemistry and Pharmacology, 2nd ed.; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1984; Volume 8, pp. 85–98. [Google Scholar]
- Lafont, R.; Harmatha, J.; Marion-Poll, F.; Dinan, L.; Wilson, I.D. The Ecdysone Handbook. Available online: http://ecdybase.org/ (accessed on 1 May 2018).
- Lafont, R.; Dauphin-Villemant, C.; Warren, J.T.; Rees, H.H. Ecdysteroid Chemistry and Biochemistry. In Reference Module in Life Sciences; Roitberg, B.D., Ed.; Elsevier: New York, NY, USA, 2017; pp. 125–195. [Google Scholar]
- Festucci-Buselli, R.A.; Contim, L.A.S.; Barbosa, L.C.A.; Stuart, J.; Otoni, W.C. Biosynthesis and potential functions of the ecdysteroid 20-hydroxyecdysone—A review. Botany 2008, 86, 978–987. [Google Scholar] [CrossRef]
- Bathori, M.; Pongracz, Z. Phytoecdysteroids-from isolation to their effects on humans. Curr. Med. Chem. 2005, 12, 153–172. [Google Scholar] [CrossRef] [PubMed]
- Thiem, B.; Kikowska, M.; Maliński, M.P.; Kruszka, D.; Napierała, M.; Florek, E. Ecdysteroids: Production in plant in vitro cultures. Phytochem. Rev. 2017, 16, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Baltaev, U.A. Phytoecdysteroids: Structure, sources, and biosynthesis in plants. Russ. J. Bioorg. Chem. 2000, 26, 799–831. [Google Scholar] [CrossRef]
- Speranza, A. Into the world of steroids: A biochemical “keep in touch” in plants and animals. Plant Signal. Behav. 2010, 5, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Soriano, I.R.; Riley, I.T.; Potter, M.J.; Bowers, W.S. Phytoecdysteroids: A Novel Defense Against Plant-Parasitic Nematodes. J. Chem. Ecol. 2004, 30, 1885–1899. [Google Scholar] [CrossRef] [PubMed]
- Jurenka, R.; Russell, K.; O’Neal, M. Phytoecdysteroids as antifeedants towards several beetles that include polyphagous and monophagous feeding guilds. Pest Manag. Sci. 2017, 73, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.A.; Grebenok, R.J.; Ohnmeiss, T.E.; Bowers, W.S. Interactions between Spinacia oleracea and Bradysia impatiens: A role for phytoecdysteroids. Arch. Insect Biochem. Physiol. 2002, 51, 204–221. [Google Scholar] [CrossRef] [PubMed]
- Dinan, L.; Lafont, R. Effects and applications of arthropod steroid hormones (ecdysteroids) in mammals. J. Endocrinol. 2006, 191, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kizelsztein, P.; Govorko, D.; Komarnytsky, S.; Evans, A.; Wang, Z.; Cefalu, W.T.; Raskin, I. 20-Hydroxyecdysone decreases weight and hyperglycemia in a diet-induced obesity mice model. Am. J. Physiol. Endoc. Metab. 2009, 296, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Gorelick-Feldman, J.; MacLean, D.; Ilic, N.; Poulev, A.; Lila, M.A.; Cheng, D.; Raskin, I. Phytoecdysteroids increase protein synthesis in skeletal muscle cells. J. Agric. Food Chem. 2008, 56, 3532–3537. [Google Scholar] [CrossRef] [PubMed]
- Lafont, R.; Dinan, L. Practical uses for ecdysteroids in mammals including humans: An update. J. Insect Sci. 2003, 3, 1–30. [Google Scholar] [CrossRef]
- Bathori, M.; Toth, N.; Hunyadi, A.; Marki, A.; Zador, E. Phytoecdysteroids and anabolic-androgenic steroids-structure and effects on humans. Curr. Med. Chem. 2008, 15, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Parr, M.K.; Zhao, P.; Haupt, O.; Ngueu, S.T.; Hengevoss, J.; Fritzemeier, K.H.; Piechotta, M.; Schlorer, N.; Muhn, P.; Zheng, W.Y.; et al. Estrogen receptor beta is involved in skeletal muscle hypertrophy induced by the phytoecdysteroid ecdysterone. Mol. Nutr. Food Res. 2014, 58, 1861–1872. [Google Scholar] [CrossRef] [PubMed]
- Kutyumov, V.A.; Maltseva, A.L.; Kotenko, O.N.; Ostrovsky, A.N. Functional differentiation in bryozoan colonies: A proteomic analysis. Cell Tissue Biol. 2016, 10, 152–159. [Google Scholar] [CrossRef]
- Fish, J.D.; Fish, S. Bryozoa. In A Student’s Guide to the Seashore; Springer: Dordrecht, The Netherlands, 1989; pp. 356–366. [Google Scholar]
- Figuerola, B.; Núñez-Pons, L.; Moles, J.; Avila, C. Feeding repellence in Antarctic bryozoans. Naturwissenschaften 2013, 100, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Figuerola, B.; Angulo-Preckler, C.; Núñez-Pons, L.; Moles, J.; Sala-Comorera, L.; García-Aljaro, C.; Blanch, A.R.; Avila, C. Experimental evidence of chemical defence mechanisms in Antarctic bryozoans. Mar. Environ. Res. 2017, 129, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed]
- Michael, P.; Hansen, K.O.; Isaksson, J.; Andersen, J.H.; Hansen, E. A novel brominated alkaloid securidine a, isolated from the marine bryozoan Securiflustra securifrons. Molecules 2017, 22, 1236. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.O.; Isaksson, J.; Bayer, A.; Johansen, J.A.; Andersen, J.H.; Hansen, E. Securamine derivatives from the arctic bryozoan Securiflustra securifrons. J. Nat. Prod. 2017, 80, 3276–3283. [Google Scholar] [CrossRef] [PubMed]
- Hoffmeister, H.; Nakanishi, K.; Koreeda, M.; Hsu, H.Y. The moulting hormone activity of ponasterones in the Calliphora test. J. Insect Physiol. 1968, 14, 53–54. [Google Scholar] [CrossRef]
- Wilson, J.E.; Håkansson, E. Molting by Cupuladria doma, A Free-Living Bryozoan. Bull. Mar. Sci. 1989, 44, 1152–1158. [Google Scholar]
- Vokáč, K.; Buděšnský, M.; Harmatha, J.; Kohoutová, J. Ecdysteroid constituents of the mushroom Tapinella panuoides. Phytochemistry 1998, 49, 2109–2114. [Google Scholar] [CrossRef]
- Buděšínský, M.; Vokáč, K.; Harmatha, J.; Cvačka, J. Additional minor ecdysteroid components of Leuzea carthamoides. Steroids 2008, 73, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Bandara, B.M.R.; Jayasinghe, L.; Karunaratne, V.; Wannigama, G.P.; Bokel, M.; Kraus, W.; Sotheeswaran, S. Ecdysterone from stem of Diploclisia glaucescens. Phytochemistry 1989, 28, 1073–1075. [Google Scholar] [CrossRef]
- Ferro, N.; Tacoronte, J.E.; Reinard, T.; Bultinck, P.; Montero, L.A. Structure–activity analysis on ecdysteroids: A structural and quantum chemical approach based on two biological systems. J. Mol. Struct. Theochem 2006, 758, 263–274. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Minakuchi, C.; Takahashi, K.; Ueno, T. Inhibition of [3H]ponasterone A binding by ecdysone agonists in the intact Kc cell line. Insect Biochem. Mol. Biol. 2002, 3, 175–180. [Google Scholar] [CrossRef]
- Krishnakumaran, A.; Schneiderman, H.A. Induction of molting in crustacea by an insect molting hormone. Gen. Comp. Endocrinol. 1969, 12, 515–518. [Google Scholar] [CrossRef]
- Hikino, H.; Takemoto, T. Ecdysones of Plant Origin. In Invertebrate Endocrinology and Hormonal Heterophylly, 1st ed.; Burdette, W.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1974; pp. 185–203. [Google Scholar]
- Tarkowska, D.; Strnad, M. Plant ecdysteroids: plant sterols with intriguing distributions, biological effects and relations to plant hormones. Planta 2016, 244, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Tomaschko, K.-H. Defensive Secretion of Ecdysteroids in Pycnogonum litorale (Arthropoda, Pantopoda). Z. Naturforsch. 1994, 49, 367–371. [Google Scholar]
- Adler, J.H.; Grebenok, R.J. Occurrence, biosynthesis, and putative role of ecdysteroids in plants. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 253–264. [Google Scholar] [CrossRef] [PubMed]
- ThermoFischer SCIENTIFIC. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/brochures/lanthascreen-tr-fret-coregulator-protocol.pdf (accessed on 1 May 2018).
- Hanssen, K.O.; Andersen, J.H.; Stiberg, T.; Engh, R.A.; Svenson, J.; Geneviere, A.M.; Hansen, E. Antitumoral and mechanistic studies of ianthelline isolated from the Arctic sponge Stryphnus fortis. Anticancer Res. 2012, 32, 4287–4297. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available. The extract of A. gelatinosum is stored at the Norwegian national biobank (Marbank) under ID M14055. |




{kind=link}
{kind=link}
{kind=link}
| Ponasterone F (1) | |||
|---|---|---|---|
| Position | δC, Type | δH (J in Hz) | δOH (J in Hz) |
| 1 | 36.6, CH2 | 1.25, dd, 13.3, 11.9/1.58, dd, 13.3, 4.3 | |
| 2 | 66.7, CH | 3.64, dt, 11.7, 3.3 | |
| 3 | 66.7, CH | 3.73, d, 3.8 | 4.35 |
| 4 | 31.8, CH2 | 1.49, m 1,2 | 4.37 |
| 5 | 50.1, CH | 2.19, dd, 11.9, 5.3 | |
| 6 | 201.9, C | ||
| 7 | 120.7, CH | 5.45, s | |
| 8 | 164.9, C | ||
| 9 | 37.5, CH | 2.59, t, 7.7 | |
| 10 | 37.3, C | ||
| 11 | 21.4, CH2 | 1.75, ddt, 13.4, 6.6, 3.1/1.59, m 1,2 | |
| 12 | 38.7, CH2 | 2.15, m 1,2/1.49, m 1,2 | |
| 13 | 45.0, C | ||
| 14 | 55.0, CH | 2.13, m 1,2 | |
| 15 | 22.01, CH2 | 1.55, m 1,2/1.44, m 1,2 | |
| 16 | 21.3, CH2 | 1.89, m 2/1.51, m 1,2 | |
| 17 | 54.4, CH | 1.66, t, 9.4 | |
| 18 | 14.0, CH3 | 0.71, s | |
| 19 | 24.1, CH3 | 0.83, s | |
| 20 | 75.4, C | 3.62 | |
| 21 | 20.8, CH3 | 1.08, s | |
| 22 | 75.5, CH | 3.12, dd, 10.0, 1.7 | 4.36 |
| 23 | 29.0, CH2 | 1.38, m 1,2/1.09, m 1,2 | |
| 24 | 36.1, CH2 | 1.37, m 1,2/1.13, m 1,2 | |
| 25 | 27.4, CH | 1.50, m 1,2 | |
| 26 | 23.0, CH3 | 0.86, d, 6.6 | |
| 27 | 22.3, CH3 | 0.85, d, 6.6 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hansen, K.Ø.; Isaksson, J.; Glomsaker, E.; Andersen, J.H.; Hansen, E. Ponasterone A and F, Ecdysteroids from the Arctic Bryozoan Alcyonidium gelatinosum. Molecules 2018, 23, 1481. https://doi.org/10.3390/molecules23061481
Hansen KØ, Isaksson J, Glomsaker E, Andersen JH, Hansen E. Ponasterone A and F, Ecdysteroids from the Arctic Bryozoan Alcyonidium gelatinosum. Molecules. 2018; 23(6):1481. https://doi.org/10.3390/molecules23061481
Chicago/Turabian StyleHansen, Kine Østnes, Johan Isaksson, Eirin Glomsaker, Jeanette Hammer Andersen, and Espen Hansen. 2018. "Ponasterone A and F, Ecdysteroids from the Arctic Bryozoan Alcyonidium gelatinosum" Molecules 23, no. 6: 1481. https://doi.org/10.3390/molecules23061481
APA StyleHansen, K. Ø., Isaksson, J., Glomsaker, E., Andersen, J. H., & Hansen, E. (2018). Ponasterone A and F, Ecdysteroids from the Arctic Bryozoan Alcyonidium gelatinosum. Molecules, 23(6), 1481. https://doi.org/10.3390/molecules23061481