Statistical Optimization of Medium for Pullulan Production by Aureobasidium pullulans NCPS2016 Using Fructose and Soybean Meal Hydrolysates

Abstract
:1. Introduction
2. Results and Discussion
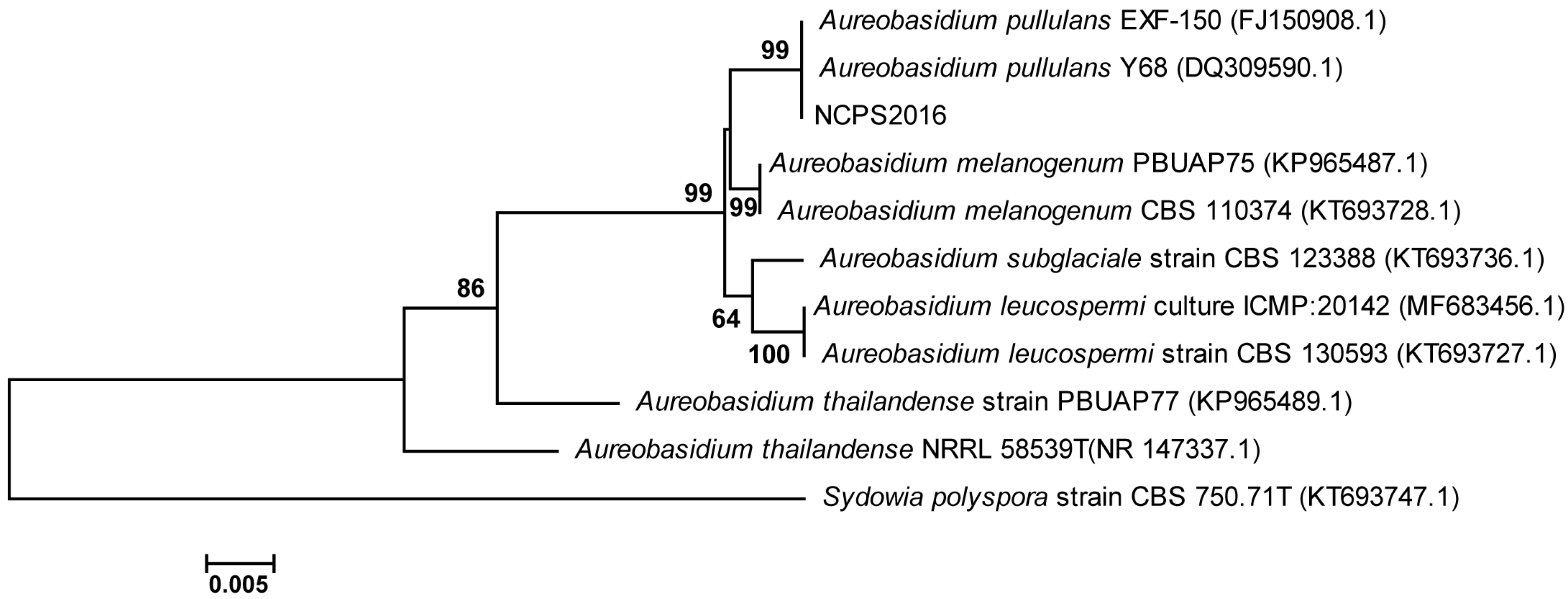
2.1. The Phylogenetic Analysis of Strain NCPS2016
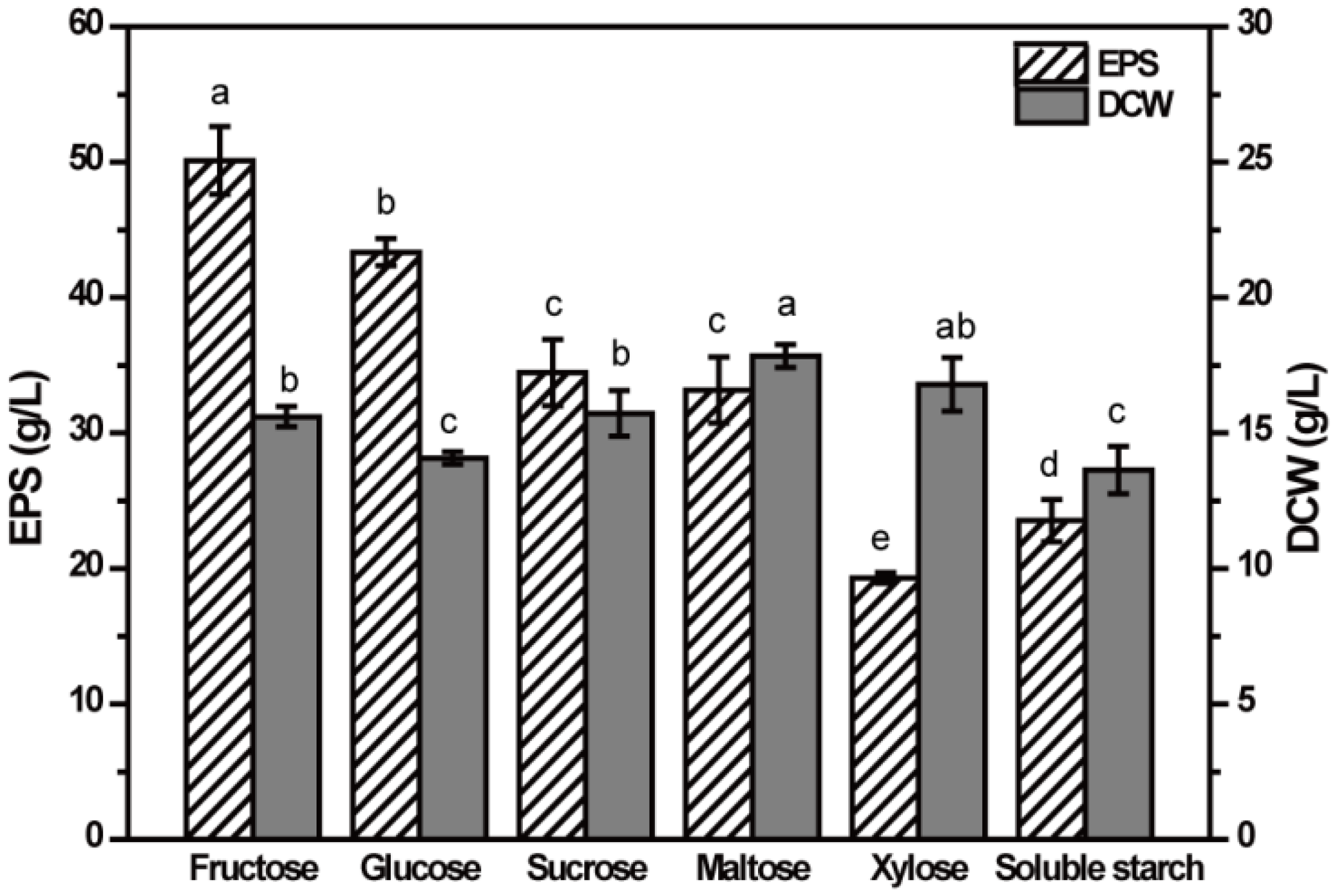
2.2. Effect of Carbon Source on EPS Production
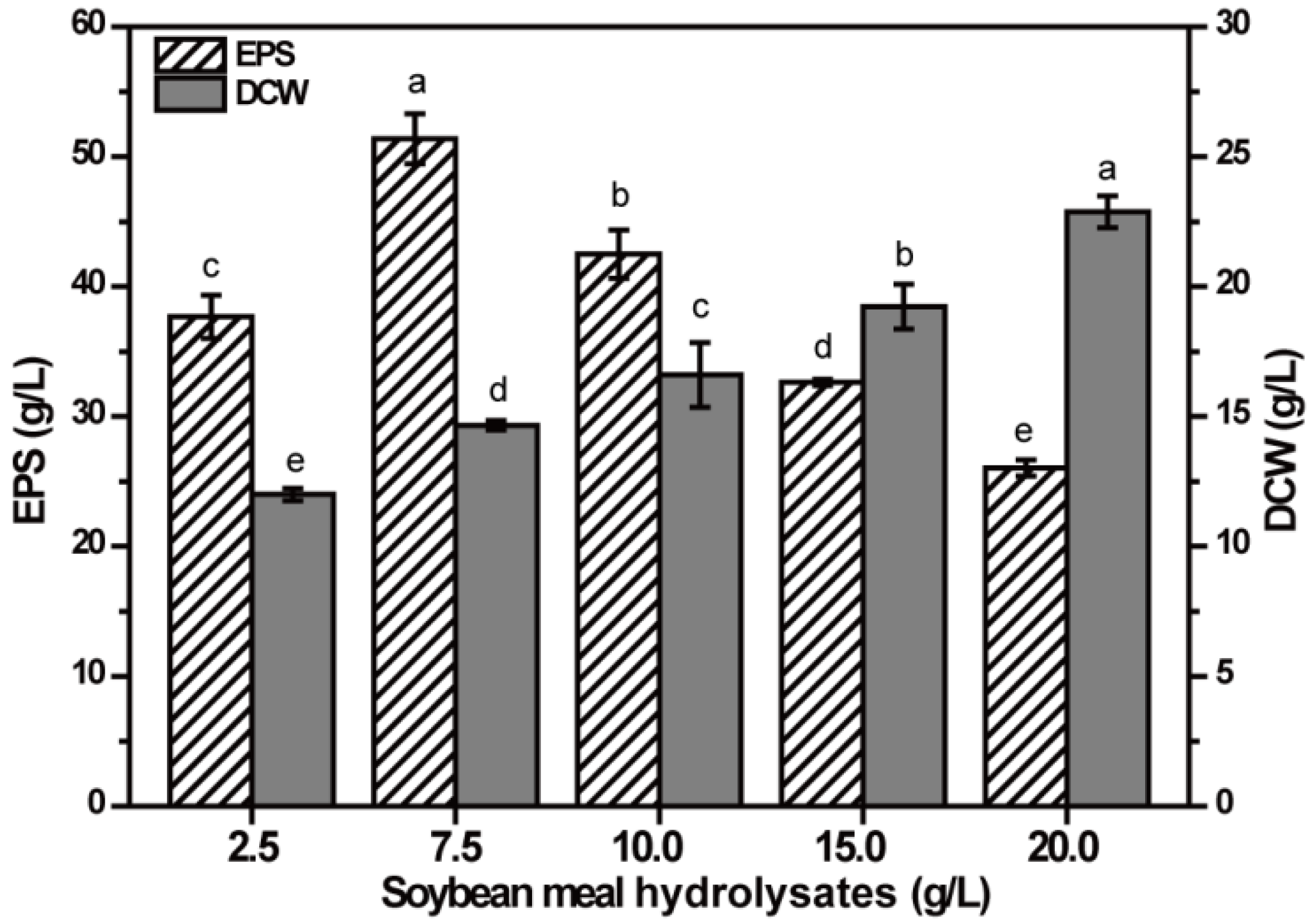
2.3. Effect of SMHs Concentration on EPS Production
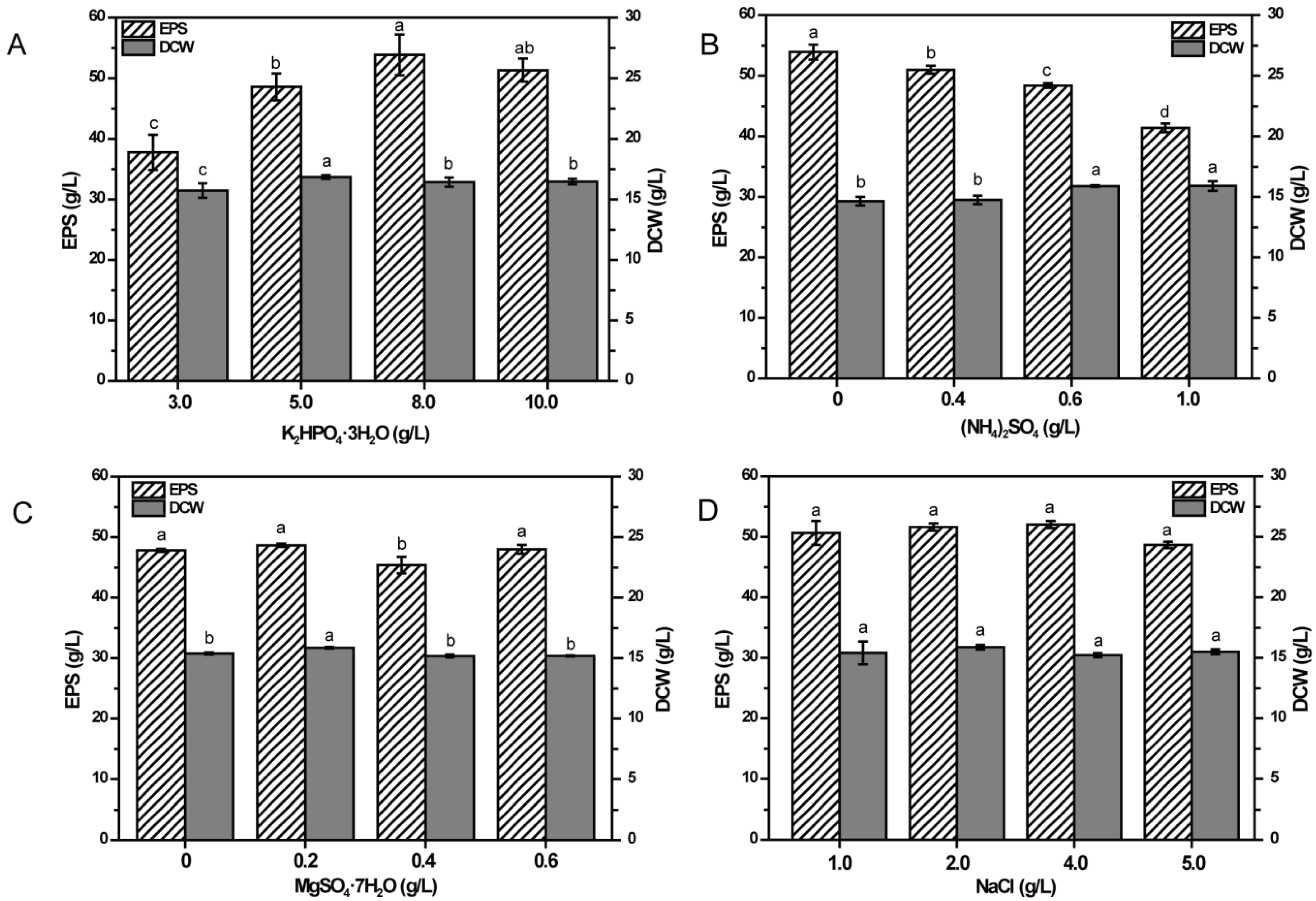
2.4. Effects of Inorganic Salts on EPS Production
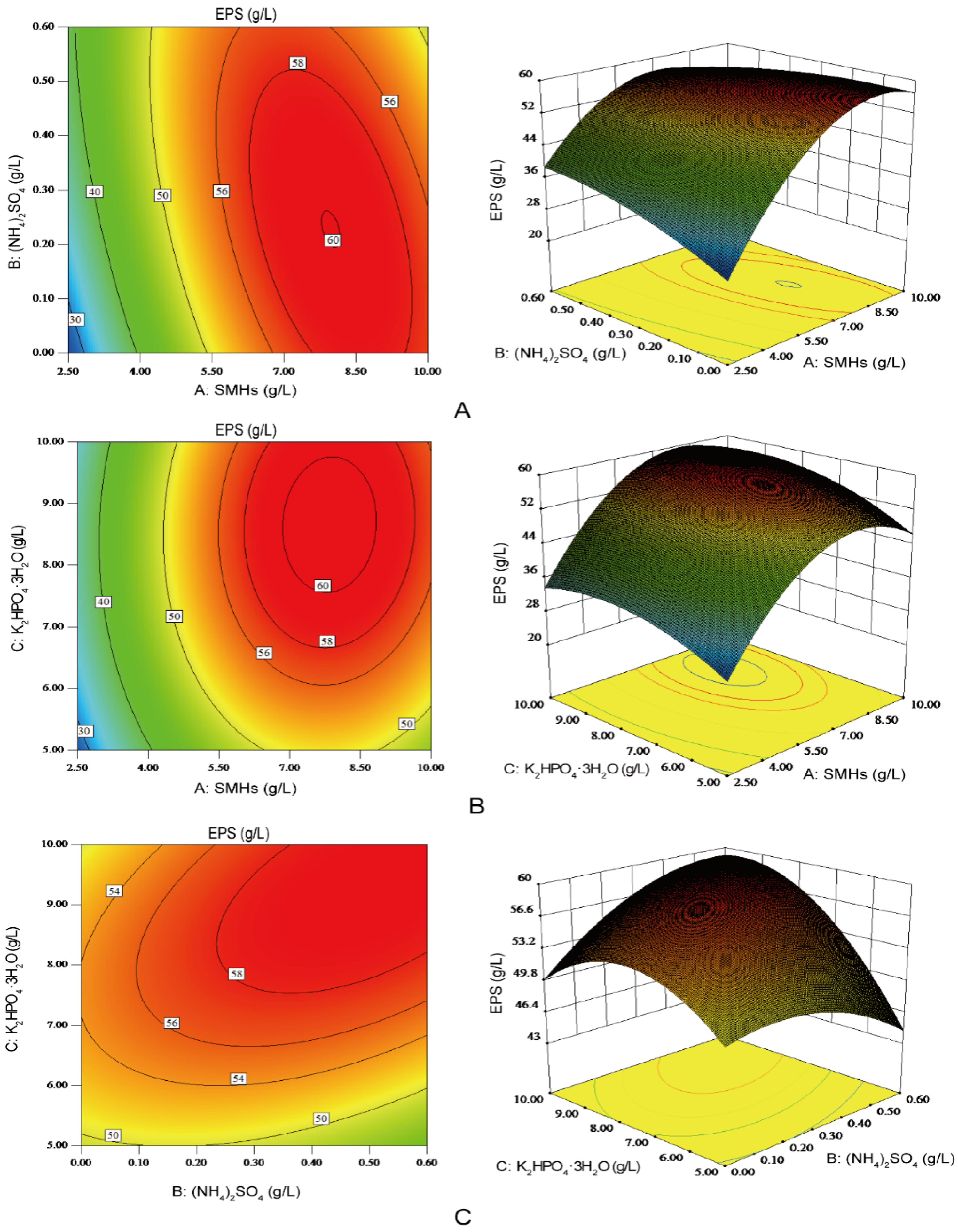
2.5. Optimization of the Medium Components Using Box–Behnken Design (BBD) for EPS Production
2.6. Verification of the Optimal Conditions on EPS Production
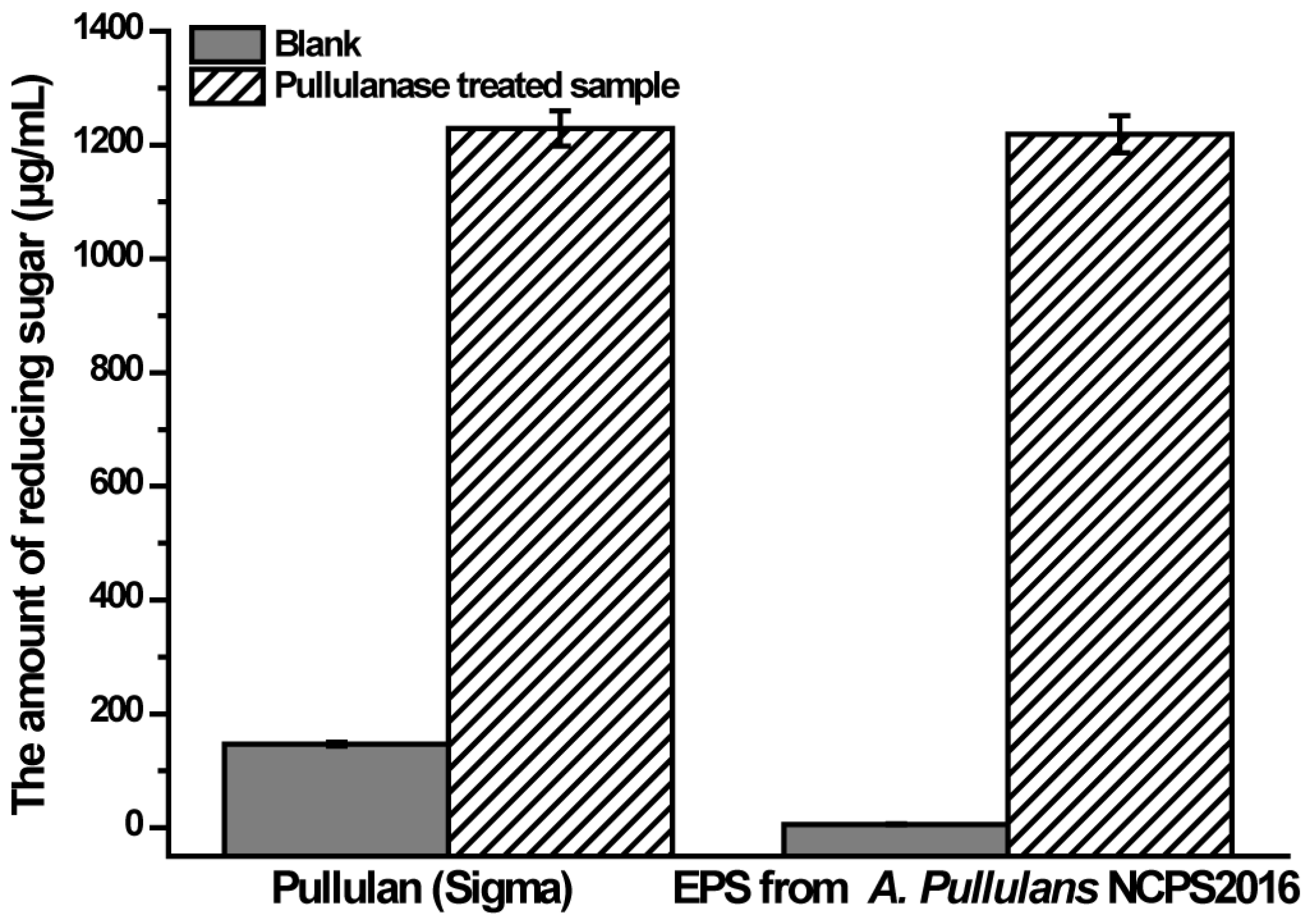
2.7. Pullulanase Hydrolysis Analysis of the Purified EPS
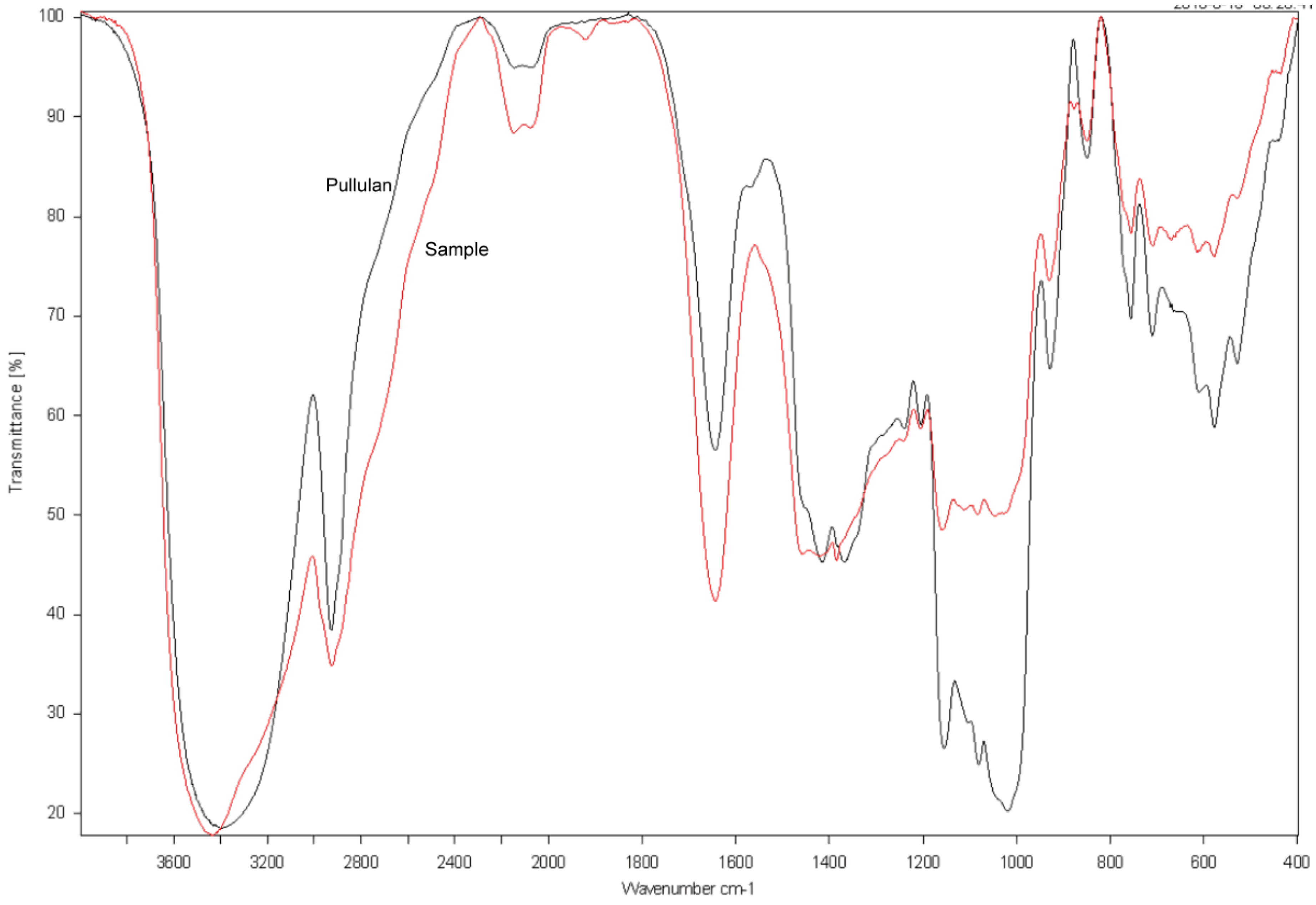
2.8. FTIR Spectroscopy of the Purified EPS Produced by NCPS2016
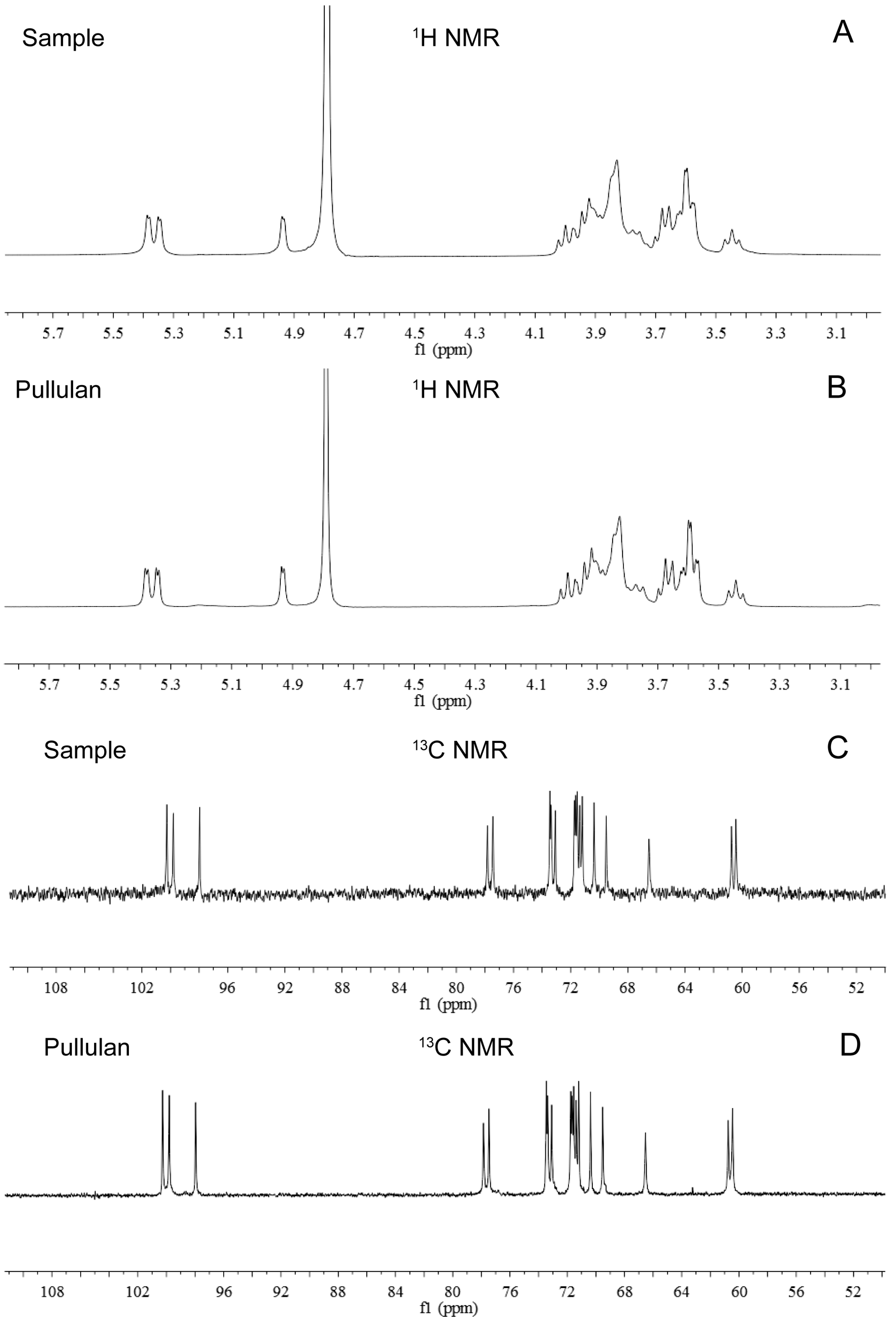
2.9. Nuclear Magnetic Resonance (NMR) Spectroscopy of the Purified EPS by NCPS2016
3. Materials and Methods
3.1. Strain and Media
3.2. Inoculum Preparation and Flask Fermentation
3.3. Preparation of Crude EPS and Determination of Dry Weight of Cells (DCW)
3.4. Effect of Varied Carbon Sources on the Production of EPS
3.5. Optimization by Single-Factor Test for EPS Production
3.6. Optimization by RSM for EPS Production
3.7. Statistical Analysis
3.8. Purification of Crude EPS
3.9. Hydrolysis Analysis of the Purified EPS
3.10. Fourier-Transform Infrared (FTIR) Spectroscopy of the Purified EPS
3.11. Nuclear Magnetic Resonance (NMR) Spectroscopy of the Purified EPS
3.12. DNA Extraction, PCR Amplification, ITS Sequencing, and Phylogenetic Analysis
3.13. Nucleotide Sequence Accession Number
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bender, H.; Lehmann, J.; Wallenfels, K. Pullulan, an extracellular glucan from Pullularia pullulans. BBA 1959, 36, 309–316. [Google Scholar] [PubMed]
- Li, Y.; Chi, Z.; Wang, G.Y.; Wang, Z.P.; Liu, G.L.; Lee, C.F.; Ma, Z.C.; Chi, Z.M. Taxonomy of Aureobasidium spp. and biosynthesis and regulation of their extracellular polymers. Crit. Rev. Microbiol. 2015, 41, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Ates, O. Systems biology of microbial exopolysaccharides production. Front. Bioeng. Biotechnol. 2015, 3, 200. [Google Scholar] [CrossRef] [PubMed]
- Farris, S.; Unalan, I.U.; Introzzi, L.; Fuentes-Alventosa, J.M.; Cozzolino, C.A. Pullulan-based films and coatings for food packaging: Present applications, emerging opportunities, and future challenges. J. Appl. Polym. Sci. 2014, 131, 40539. [Google Scholar] [CrossRef]
- Oguzhan, P.; Yangilar, F. Pullulan: Production and usage in food industry. Afr. J. Food Sci. Technol. 2013, 4, 57–63. [Google Scholar]
- Synowiec, A.; Gniewosz, M.; Krasniewska, K.; Chlebowska-Smigiel, A.; Przybyl, J.L.; Baczek, K.; Weglarz, Z. Effect of meadowsweet flower extract-pullulan coatings on rhizopus rot development and postharvest quality of cold-stored red peppers. Molecules 2014, 19, 12925–12939. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L. Pullulan-based coatings for preservation of razor clam Sinonovacula constricta. Int. J. Biol. Macromol. 2016, 92, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Kraśniewska, K.; Ścibisz, I.; Gniewosz, M.; Mitek, M.; Pobiega, K.; Cendrowski, A. Effect of pullulan coating on postharvest quality and shelf-life of highbush blueberry (Vaccinium corymbosum L.). Materials 2017, 10, 965. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dou, B.; Bao, Y. Efficient targeted pDNA/siRNA delivery with folate-low-molecular-weight polyethyleneimine-modified pullulan as non-viral carrier. Mater. Sci. Eng. C 2014, 34, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; An, T.; Wang, D.; Wan, G.; Zhang, M.; Wang, H.; Zhang, S.; Li, R.; Yang, X.; Wang, Y. Stepwise pH-responsive nanoparticles containing charge-reversible pullulan-based shells and poly(β-amino ester)/poly(lactic-co-glycolic acid) cores as carriers of anticancer drugs for combination therapy on hepatocellular carcinoma. J. Control. Release 2016, 226, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Nakashio, S.; Tsuji, K.; Toyota, N.; Fujita, F. Novel Cosmetics Containing Pullulan. U.S. Patent 3,972,997, 3 August 1976. [Google Scholar]
- Peterson, S.; Manitchotpisit, P.; Leathers, T. Aureobasidium thailandense sp. Nov. isolated from leaves and wooden surfaces. Int. J. Syst. Evol. Microbiol. 2013, 63, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Wang, F.; Chi, Z.; Yue, L.; Liu, G.; Zhang, T. Bioproducts from Aureobasidium pullulans, a biotechnologically important yeast. Appl. Microbiol. Biotechnol. 2009, 82, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, J.; Su, Y.; Zhu, Y.; Zhang, G.; Zhao, H.; Liu, H.; Yang, Y.; Nian, R.; Zhang, H.; et al. Pullulan production from synthetic medium by a new mutant of Aureobasidium pullulans. Prep. Biochem. Biotech. 2017, 47, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.R.; Saluja, P.; Prasad, G.S. Pullulan production by an osmotolerant Aureobasidium pullulans RBF-4A3 isolated from flowers of Caesulia axillaris. Carbohyd. Polym. 2011, 83, 1547–1552. [Google Scholar] [CrossRef]
- Padmanaban, S.; Balaji, N.; Muthukumaran, C.; Tamilarasan, K. Statistical optimization of process parameters for exopolysaccharide production by Aureobasidium pullulans using sweet potato based medium. 3 Biotech 2015, 5, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Demirci, A.; Catchmark, J.M. Evaluation of medium composition and cultivation parameters on pullulan production by Aureobasidium pullulans. Food Sci. Technol. Int. 2011, 17, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Tong, Q.; Ma, M. Why sucrose is the most suitable substrate for pullulan fermentation by Aureobasidium pullulans CGMCC1234? Enzyme Microb. Technol. 2016, 92, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. Efficient Fermentative Production of Pullulan by Aureobasidium Pullulans: Batch Process Optimization and Its Regulation Mechanism. Ph.D. Dissertation, Nanjing Agricultural University, Nanjing, China, 2015. [Google Scholar]
- Duan, X.; Chi, Z.; Wang, L.; Wang, X. Influence of different sugars on pullulan production and activities of alpha-phosphoglucose mutase, UDPG-pyrophosphorylase and glucosyltransferase involved in pullulan synthesis in Aureobasidium pullulans Y68. Carbohydr. Polym. 2008, 73, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Demirci, A.; Catchmark, J.M. Pullulan: Biosynthesis, production, and applications. Appl. Microbiol. Biotechnol. 2011, 92, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.R.; Sharma, N.; Prasad, G.S. De-oiled jatropha seed cake is auseful nutrient for pullulan production. Microb. Cell Fact. 2012, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Prasad, G.S.; Choudhury, A.R. Cost effective production of pullulan from agri-industrial residues using response surface methodology. Int. J. Biol. Macromol. 2014, 64, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, F.; Wei, G.; Jiang, M.; Dong, M. The mechanism of improved pullulan production by nitrogen limitation in batch culture of Aureobasidium pullulans. Carbohydr. Polym. 2015, 127, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yang, J.; Chen, X.; Su, H.; Zhang, X.; Huang, F.; Zhou, B.; Xie, B. Optimization of fermentation conditions for the production of the M23 protease pseudoalterin by deep-sea Pseudoalteromonas sp. CF6-2 with artery powder as an inducer. Molecules 2014, 19, 4779–4790. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, X.; Yu, D.; Sang, B.; Feng, J.; Han, L.; Zhang, X. Optimization of fermentation conditions and bench-scale for improvement of a novel glycoprotein GP-1 production by Streptomyces kanasenisi ZX01. Molecules 2018, 23, 137. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, U.; Lu, H.; Wang, J.; Han, J.; Zhu, X.; Lu, Z.; Tayyaba, S.; Hassan, Y.I. Optimizing the maximum recovery of dihydromyricetin from Chinese vine tea, Ampelopsis grossedentata, using response surface methodology. Molecules 2017, 22, 2250. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Qiao, L.; He, H.; Zhang, Q.; Chen, X.; Zhou, W.; Zhou, B.; Zhang, Y. Optimization of fermentation conditions and rheological properties of exopolysaccharide produced by deep-sea bacterium Zunongwangia profunda SM-A87. PLoS ONE 2011, 6, e26825. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Fu, W.; Liu, G.; Wang, Z.; Chi, Z. High-level pullulan production by Aureobasidium pullulans var. melanogenium P16 isolated from mangrove system. Appl. Microbiol. Biotechnol. 2014, 98, 4865–4873. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Sun, H.; Liang, F.; Guo, H.; Liu, L. Purification and determination of the structure of pulullan polysaccharide. Chem. J. Chin. Univ. 1999, 20, 1729–1732. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds pullulan are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | A: SMHs (g/L) | B: (NH4)2SO4 (g/L) | C: K2HPO4·3H2O (g/L) |
|---|---|---|---|
| −1 | 2.50 | 0 | 5.00 |
| 0 | 6.25 | 0.30 | 7.50 |
| 1 | 10.00 | 0.60 | 10.00 |
| Run | A | B | C | EPS (g/L) | |
|---|---|---|---|---|---|
| Predicted Value | Actual Value | ||||
| 1 | −1 | −1 | 0 | 26.54 | 24.93 |
| 2 | 0 | 0 | 0 | 57.83 | 57.55 |
| 3 | 1 | 1 | 0 | 48.94 | 50.55 |
| 4 | 0 | 0 | 0 | 57.83 | 55.23 |
| 5 | 0 | −1 | −1 | 49.29 | 49.48 |
| 6 | 1 | 0 | 1 | 55.72 | 54.29 |
| 7 | 0 | −1 | 1 | 49.94 | 51.66 |
| 8 | 1 | 0 | −1 | 46.45 | 46.56 |
| 9 | 0 | 1 | 1 | 59.1 | 58.91 |
| 10 | 1 | −1 | 0 | 56.98 | 56.68 |
| 11 | −1 | 0 | −1 | 27.78 | 29.21 |
| 12 | 0 | 1 | −1 | 44.4 | 42.68 |
| 13 | 0 | 0 | 0 | 57.83 | 59.03 |
| 14 | 0 | 0 | 0 | 57.83 | 58.78 |
| 15 | −1 | 0 | 1 | 33.86 | 33.75 |
| 16 | 0 | 0 | 0 | 57.83 | 58.56 |
| 17 | −1 | 1 | 0 | 38.84 | 39.14 |
| Source | df | EPS Production a | |||
|---|---|---|---|---|---|
| Sum of Squares | Mean Square | F-Value | p-Value Prob > F | ||
| Model | 9 | 1913.83 | 212.65 | 59.13 | <0.0001 * |
| A | 1 | 821.14 | 821.14 | 228.32 | <0.0001 * |
| B | 1 | 9.1 | 9.1 | 2.53 | 0.1558 |
| C | 1 | 117.66 | 117.66 | 32.72 | 0.0007 * |
| AB | 1 | 103.43 | 103.43 | 28.76 | 0.001 * |
| AC | 1 | 2.54 | 2.54 | 0.71 | 0.4281 |
| BC | 1 | 49.35 | 49.35 | 13.72 | 0.0076 * |
| A2 | 1 | 644.02 | 644.02 | 179.07 | <0.0001 * |
| B2 | 1 | 29.29 | 29.29 | 8.14 | 0.0246 * |
| C2 | 1 | 85.64 | 85.64 | 23.81 | 0.0018 * |
| Residual | 7 | 25.17 | 3.6 | ||
| Lack of Fit | 3 | 15.46 | 5.15 | 2.12 | 0.2401 |
| Pure Error | 4 | 9.71 | 2.43 | ||
| Cor Total | 16 | 1939 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Zhang, Y.; Zhao, S.; Zhou, Q.; Xin, X.; Chen, L. Statistical Optimization of Medium for Pullulan Production by Aureobasidium pullulans NCPS2016 Using Fructose and Soybean Meal Hydrolysates. Molecules 2018, 23, 1334. https://doi.org/10.3390/molecules23061334
Yang J, Zhang Y, Zhao S, Zhou Q, Xin X, Chen L. Statistical Optimization of Medium for Pullulan Production by Aureobasidium pullulans NCPS2016 Using Fructose and Soybean Meal Hydrolysates. Molecules. 2018; 23(6):1334. https://doi.org/10.3390/molecules23061334
Chicago/Turabian StyleYang, Jinyu, Yanhao Zhang, Shuangzhi Zhao, Qingxin Zhou, Xue Xin, and Leilei Chen. 2018. "Statistical Optimization of Medium for Pullulan Production by Aureobasidium pullulans NCPS2016 Using Fructose and Soybean Meal Hydrolysates" Molecules 23, no. 6: 1334. https://doi.org/10.3390/molecules23061334
APA StyleYang, J., Zhang, Y., Zhao, S., Zhou, Q., Xin, X., & Chen, L. (2018). Statistical Optimization of Medium for Pullulan Production by Aureobasidium pullulans NCPS2016 Using Fructose and Soybean Meal Hydrolysates. Molecules, 23(6), 1334. https://doi.org/10.3390/molecules23061334