Hydroxysafflor Yellow A Attenuates Lipopolysaccharide-Induced Neurotoxicity and Neuroinflammation in Primary Mesencephalic Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
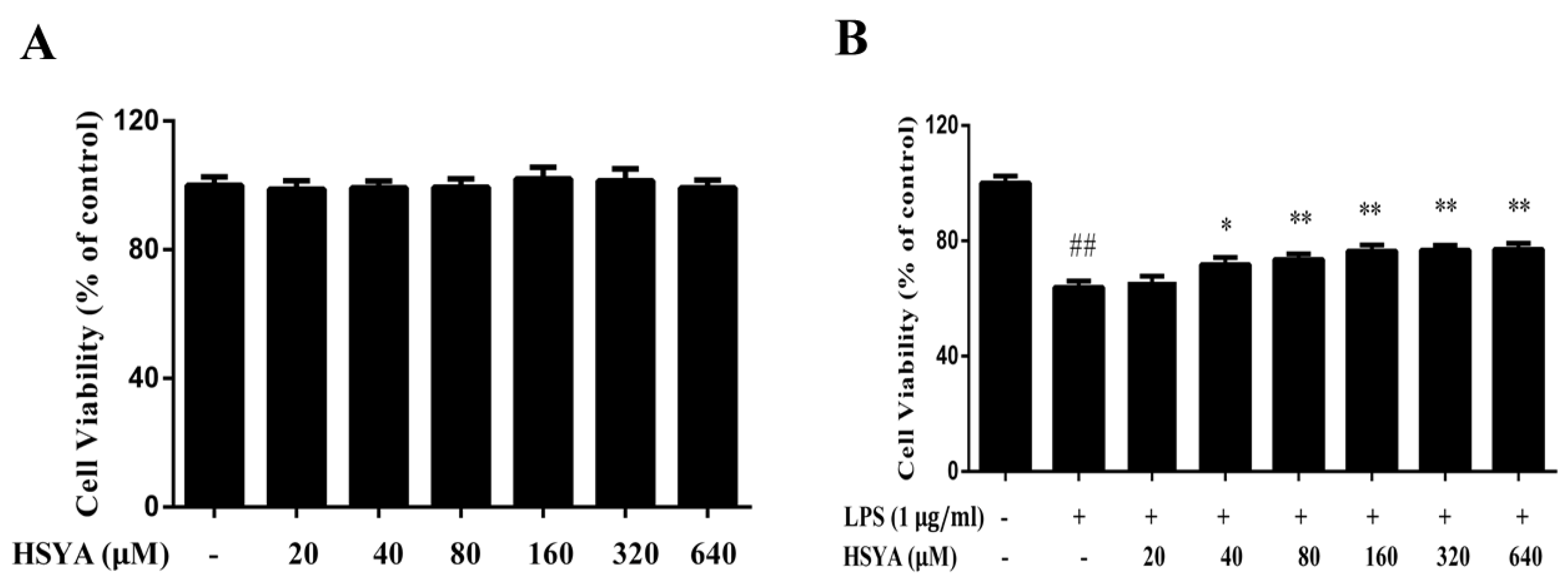
2.1. HSYA Protected Dopaminergic Neurons against LPS-Induced Neurotoxicity
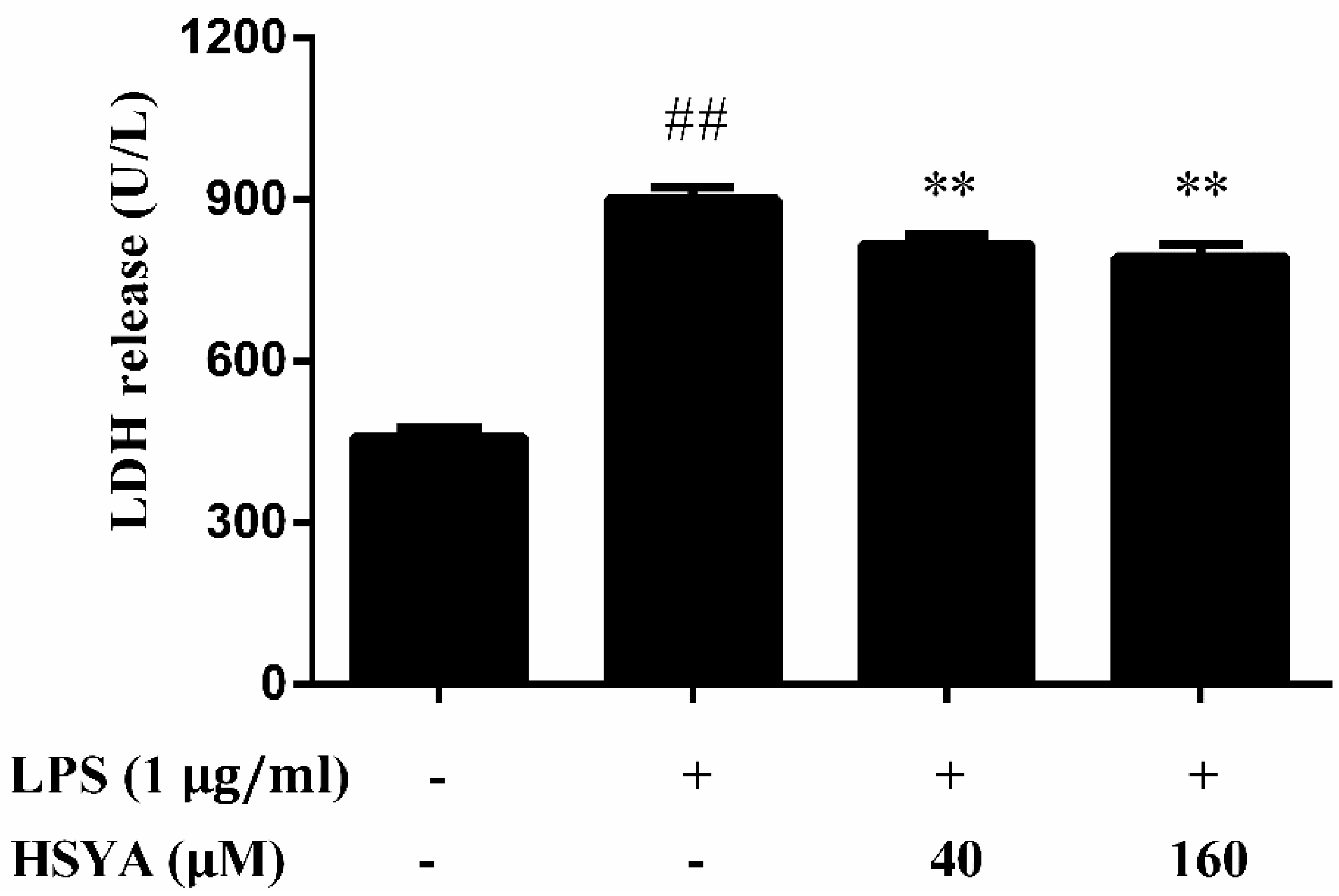
2.2. Effect of HSYA on LDH Release in LPS-Challenged Primary Mesencephalic Cultures
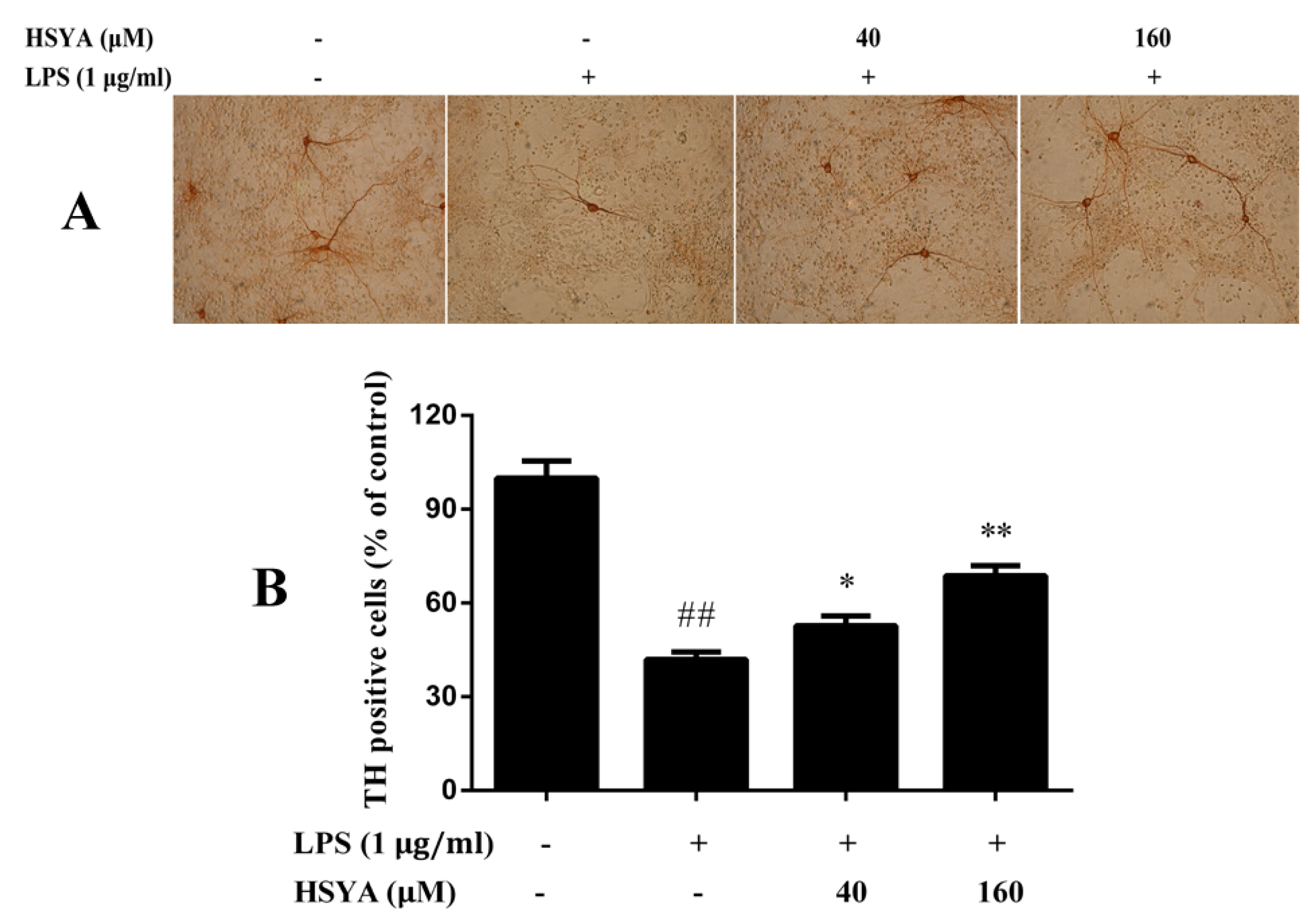
2.3. Effect of HSYA on TH Positive Cells in LPS-Challenged Primary Mesencephalic Cultures
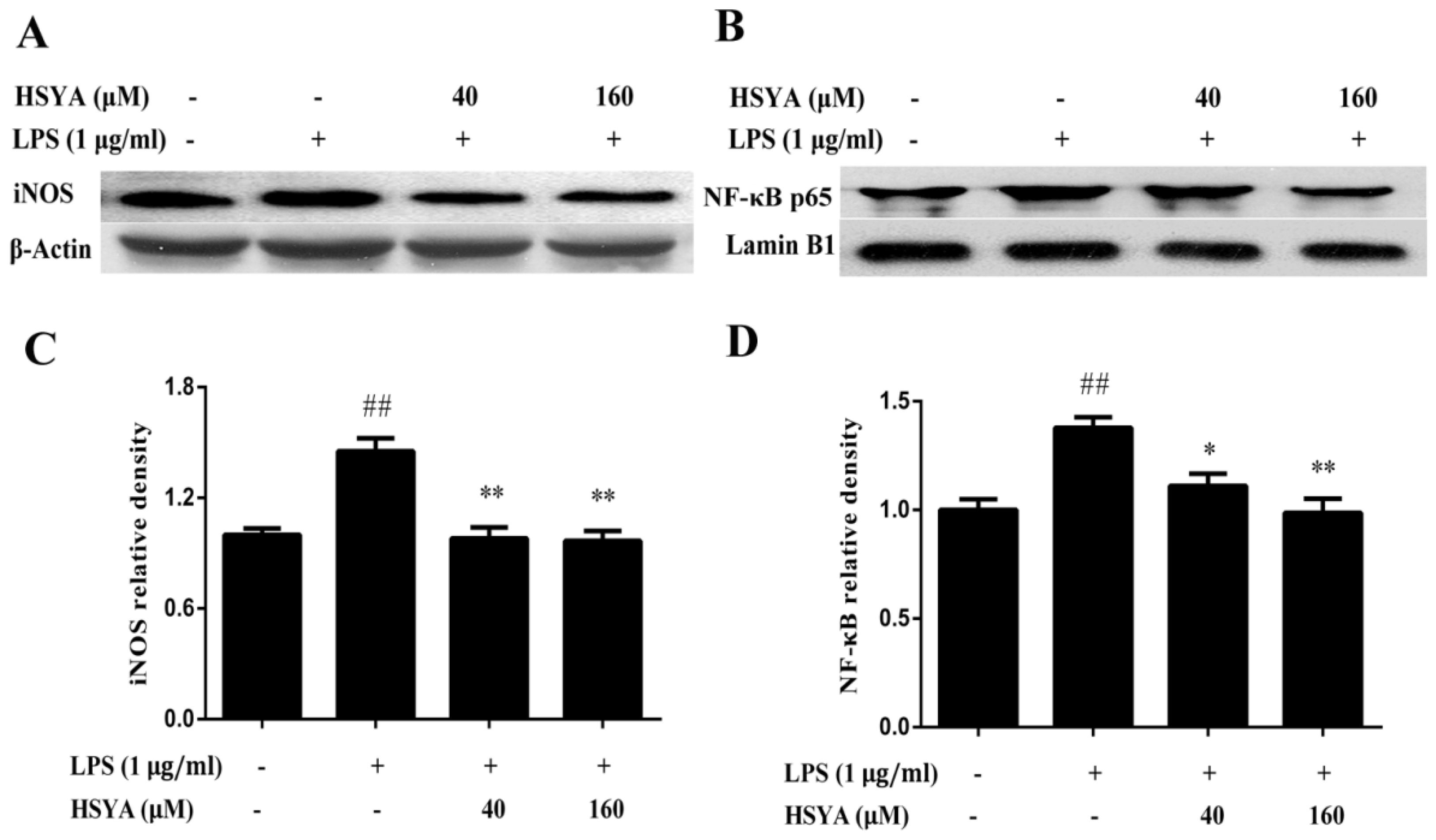
2.4. Effect of HSYA on the Expressions of NF-κB p65 and iNOS in LPS-Challenged Primary Mesencephalic Cultures
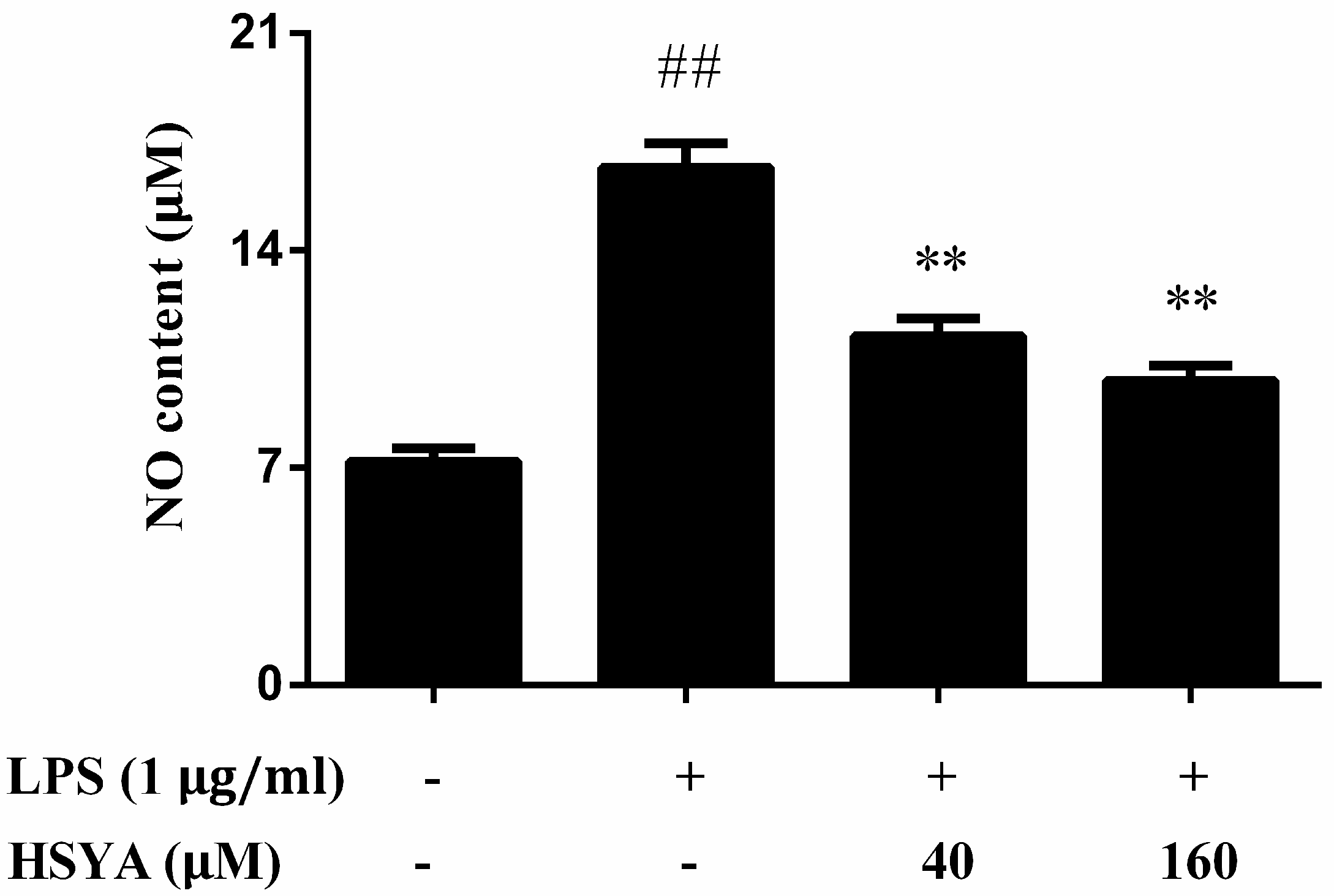
2.5. Effect of HSYA on NO Levels in LPS-Challenged in Primary Mesencephalic Cultures
2.6. Effect of HSYA on the Contents of IL-1β and TNF-α in LPS-Challenged Primary Mesencephalic Cultures
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures and Treatments
4.3. MTT Assay
4.4. Immunohistochemical Staining
4.5. Western Blotting Analysis
4.6. Nitrite Assay
4.7. Measurement of LDH Release, IL-1β and TNF-α
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaur, K.; Gill, J.S.; Bansal, P.K.; Deshmukh, R. Neuroinflammation—A major cause for striatal dopaminergic degeneration in Parkinson’s disease. J. Neurol. Sci. 2017, 381, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Kanaan, N.M.; Kordower, J.H.; Collier, T.J. Age-related changes in glial cells of dopamine midbrain subregions in rhesus monkeys. Neurobiol. Aging 2010, 31, 937–952. [Google Scholar] [CrossRef] [PubMed]
- Halliday, G.M.; Stevens, C.H. Glia: Initiators and progressors of pathology in Parkinson’s disease. Mov. Disord. 2011, 26, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Dzamko, N.; Geczy, C.L.; Halliday, G.M. Inflammation is genetically implicated in Parkinson’s disease. Neuroscience 2015, 302, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, S.; Cerami, C.; Alessio, M.; Garibotto, V.; Panzacchi, A.; Olivieri, S.; Gelsomino, G.; Moresco, R.M.; Perani, D. In vivo microglia activation in very early dementia with Lewy bodies, comparison with Parkinson’s disease. Parkinsonism Relat. Disord. 2013, 19, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, Y.; Yoshikawa, E.; Sekine, Y.; Futatsubashi, M.; Kanno, T.; Ogusu, T.; Torizuka, T. Microglial activation and dopamine terminal loss in early Parkinson’s disease. Ann. Neurol. 2005, 57, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Gillardon, F.; Schmid, R.; Draheim, H. Parkinson’s disease-linked leucine-rich repeat kinase 2(R1441G) mutation increases proinflammatory cytokine release from activated primary microglial cells and resultant neurotoxicity. Neuroscience 2012, 208, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Kum, W.F.; Durairajan, S.S.; Bian, Z.X.; Man, S.C.; Lam, Y.C.; Xie, L.X.; Lu, J.H.; Wang, Y.; Huang, X.Z.; Li, M. Treatment of idiopathic Parkinson’s disease with traditional Chinese herbal medicine: A randomized placebo-controlled pilot clinical study. Evid. Based Complement. Alternat. Med. 2011, 2011, 724353. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Zhao, H. Effects of hydroxysafflor yellow A in the attenuation of MPTP neurotoxicity in mice. Neurochem. Res. 2010, 35, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Guo, Y.; Zhao, J.; Wang, N.; Ding, J.; Liu, Q. Hydroxysafflor Yellow A Inhibits LPS-Induced NLRP3 Inflammasome Activation via Binding to Xanthine Oxidase in Mouse RAW264.7 Macrophages. Mediat. Inflamm. 2016, 2016, 8172706. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Zhu, Y.; Jin, M.; Zang, B. Hydroxysafflor Yellow A inhibits lipopolysaccharide-induced inflammatory signal transduction in human alveolar epithelial A549 cells. Fitoterapia 2013, 84, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Qian, Y.; Ou-Yang, A.; Fu, L. Hydroxysafflor Yellow A Attenuates Neuron Damage by Suppressing the Lipopolysaccharide-Induced TLR4 Pathway in Activated Microglial Cells. Cell. Mol. Neurobiol. 2016, 36, 1241–1256. [Google Scholar] [CrossRef] [PubMed]
- Castano, A.; Herrera, A.J.; Cano, J.; Machado, A. Lipopolysaccharide intranigral injection induces inflammatory reaction and damage in nigrostriatal dopaminergic system. J. Neurochem. 1998, 70, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.G.; Mohney, R.P.; Wilson, B.; Jeohn, G.H.; Liu, B.; Hong, J.S. Regional difference in susceptibility to lipopolysaccharide-induced neurotoxicity in the rat brain: Role of microglia. J. Neurosci. 2000, 20, 6309–6316. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, H.; Alexander, G.; Austenaa, L.M.; Ebihara, K.; Blomhoff, R. Molecular imaging of the transcription factor NF-kappaB, a primary regulator of stress response. Mutat. Res. 2004, 551, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Tsoulfas, G.; Geller, D.A. NF-kappaB in transplantation: Friend or foe? Transpl. Infect. Dis. 2001, 3, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Iravani, M.M.; Kashefi, K.; Mander, P.; Rose, S.; Jenner, P. Involvement of inducible nitric oxide synthase in inflammation-induced dopaminergic neurodegeneration. Neuroscience 2002, 110, 49–58. [Google Scholar] [CrossRef]
- Yuste, J.E.; Tarragon, E.; Campuzano, C.M.; Ros-Bernal, F. Implications of glial nitric oxide in neurodegenerative diseases. Front. Cell. Neurosci. 2015, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dawson, V.L.; Dawson, T.M. Role of nitric oxide in Parkinson’s disease. Pharmacol. Ther. 2006, 109, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kavya, R.; Saluja, R.; Singh, S.; Dikshit, M. Nitric oxide synthase regulation and diversity: Implications in Parkinson’s disease. Nitric Oxide 2006, 15, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, C.; Li, M.; Li, D.; Liu, K. The subchronic toxicity of hydroxysafflor yellow A of 90 days repeatedly intraperitoneal injections in rats. Toxicology 2004, 203, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Radad, K.; Gille, G.; Rausch, W.D. Dopaminergic neurons are preferentially sensitive to long-term rotenone toxicity in primary cell culture. Toxicol. In Vitro 2008, 22, 68–74. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Ding, Y.-X.; He, J.; Ma, C.-J.; Zhao, Y.; Wang, Z.-H.; Han, B. Hydroxysafflor Yellow A Attenuates Lipopolysaccharide-Induced Neurotoxicity and Neuroinflammation in Primary Mesencephalic Cultures. Molecules 2018, 23, 1210. https://doi.org/10.3390/molecules23051210
Wang T, Ding Y-X, He J, Ma C-J, Zhao Y, Wang Z-H, Han B. Hydroxysafflor Yellow A Attenuates Lipopolysaccharide-Induced Neurotoxicity and Neuroinflammation in Primary Mesencephalic Cultures. Molecules. 2018; 23(5):1210. https://doi.org/10.3390/molecules23051210
Chicago/Turabian StyleWang, Tian, Yu-Xin Ding, Jie He, Cheng-Jun Ma, Yue Zhao, Zhen-Hua Wang, and Bing Han. 2018. "Hydroxysafflor Yellow A Attenuates Lipopolysaccharide-Induced Neurotoxicity and Neuroinflammation in Primary Mesencephalic Cultures" Molecules 23, no. 5: 1210. https://doi.org/10.3390/molecules23051210
APA StyleWang, T., Ding, Y.-X., He, J., Ma, C.-J., Zhao, Y., Wang, Z.-H., & Han, B. (2018). Hydroxysafflor Yellow A Attenuates Lipopolysaccharide-Induced Neurotoxicity and Neuroinflammation in Primary Mesencephalic Cultures. Molecules, 23(5), 1210. https://doi.org/10.3390/molecules23051210